
The PDE4 Inhibitor Tanimilast Restrains the Tissue-Damaging Properties of Human Neutrophils

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results and Discussion
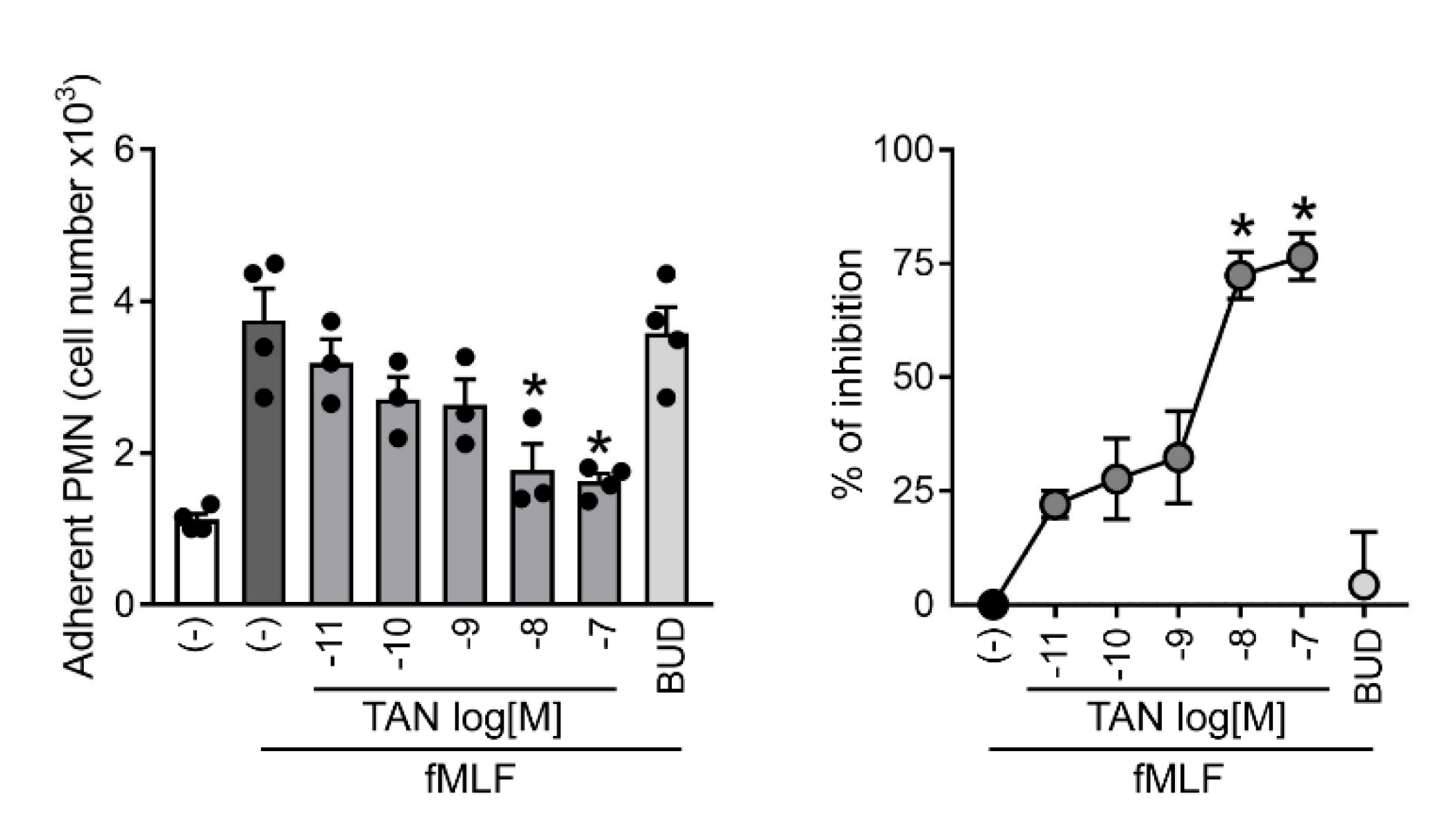
2.1. Tanimilast Inhibits Human Neutrophil Adhesion to Resting Endothelial Cells
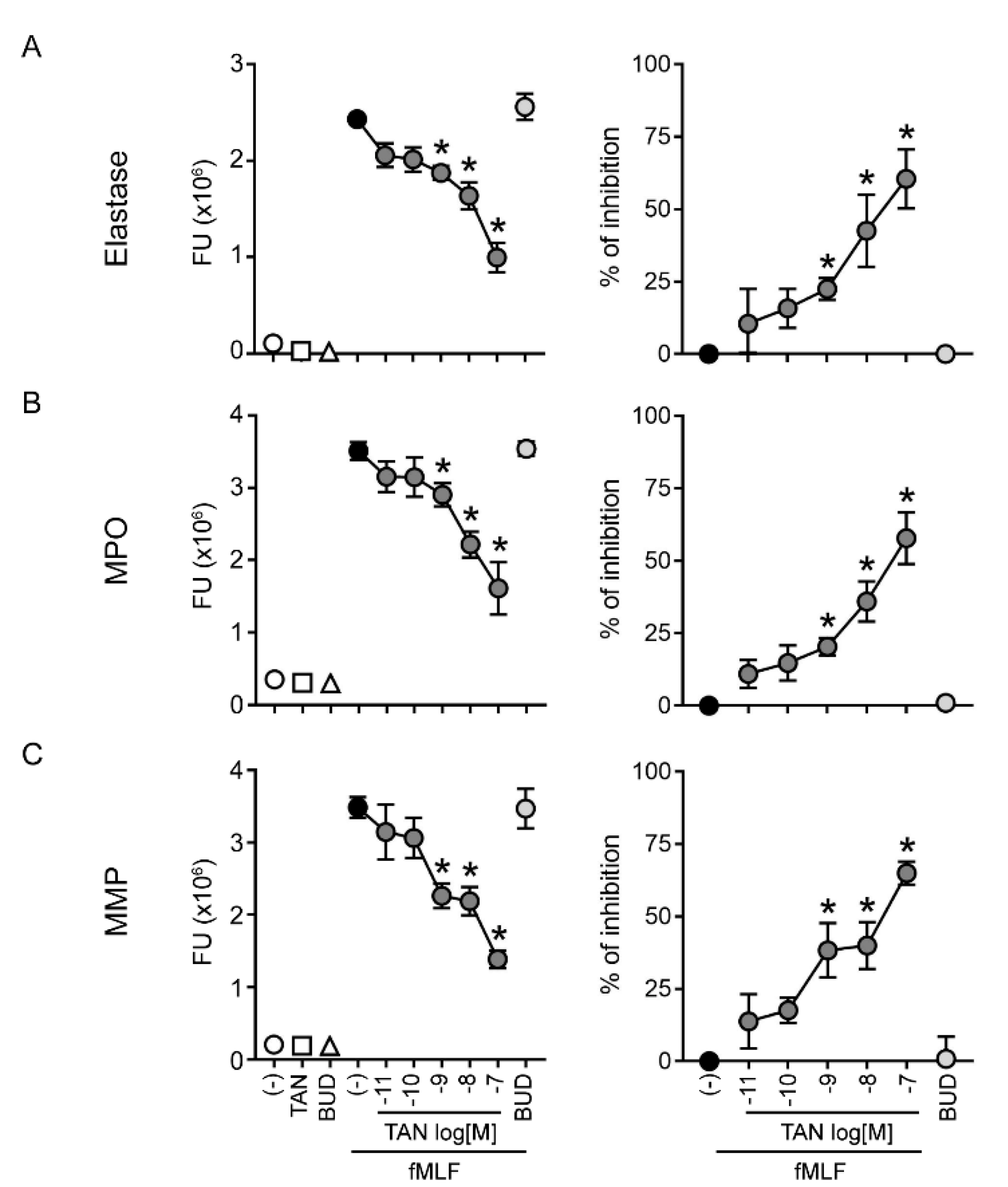
2.2. Tanimilast Restrains the Release of Granule Proteins by Human Neutrophils
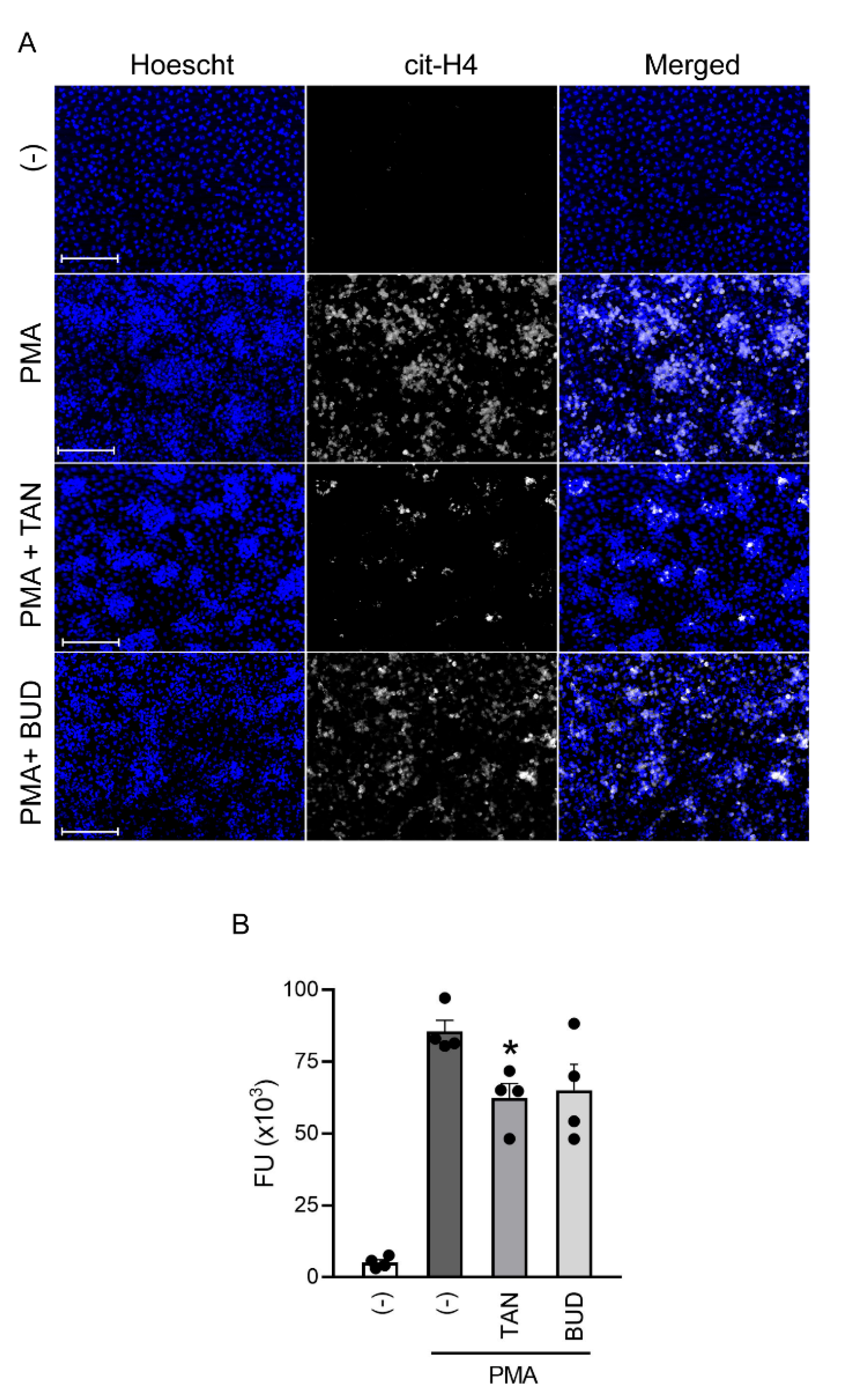
2.3. Tanimilast Reduces NET Production by Human Neutrophils
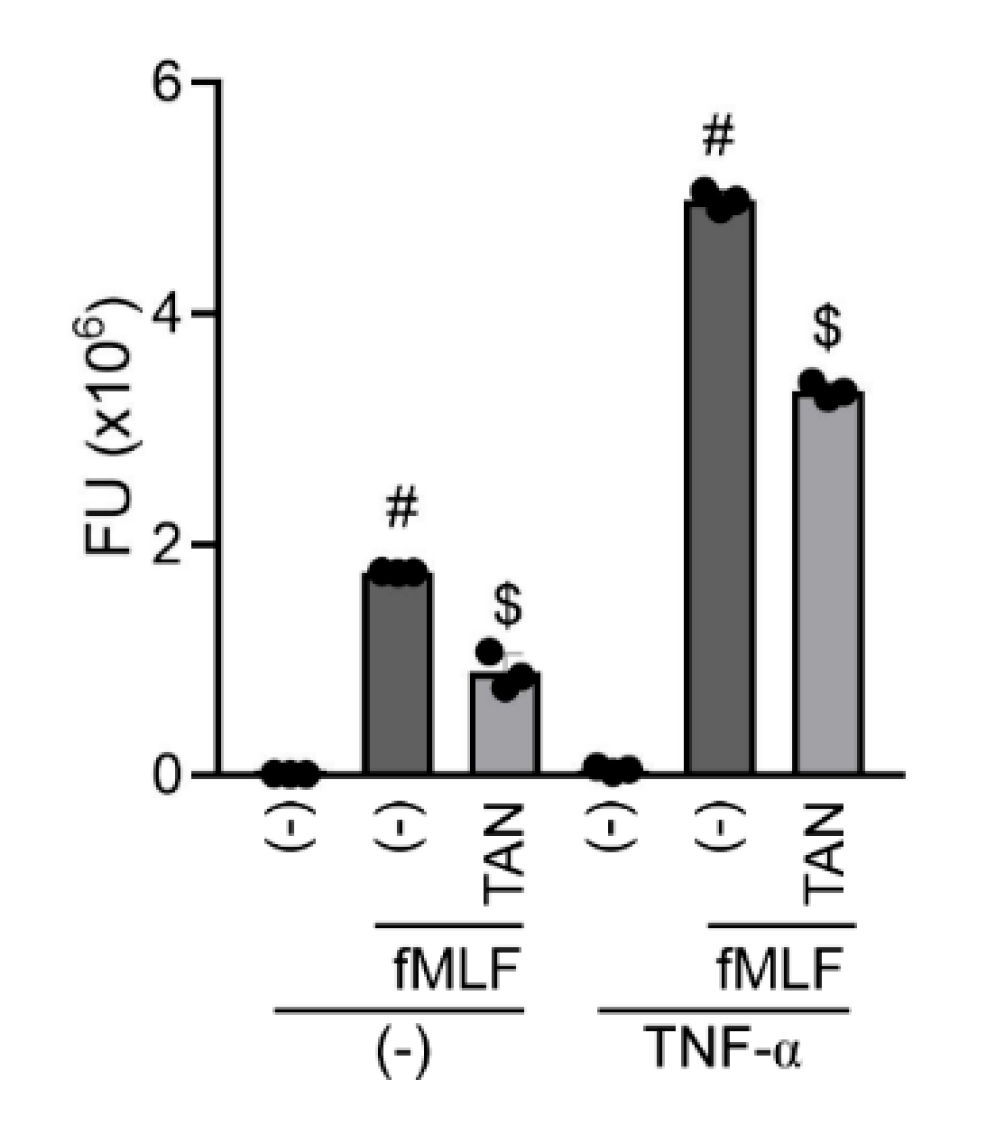
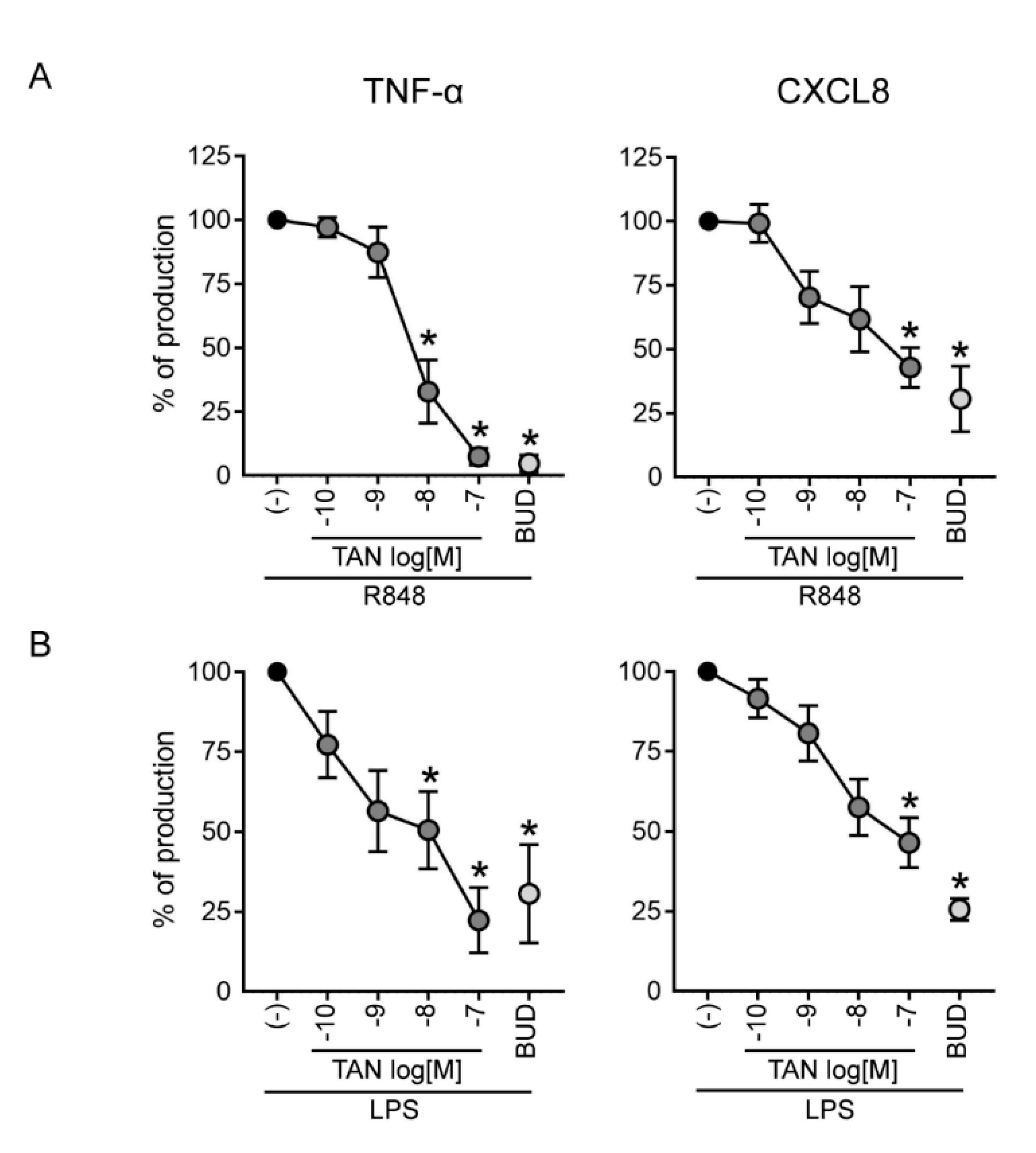
2.4. Tanimilast Inhibits the Secretion of Pro-Inflammatory Mediators by Human Neutrophils
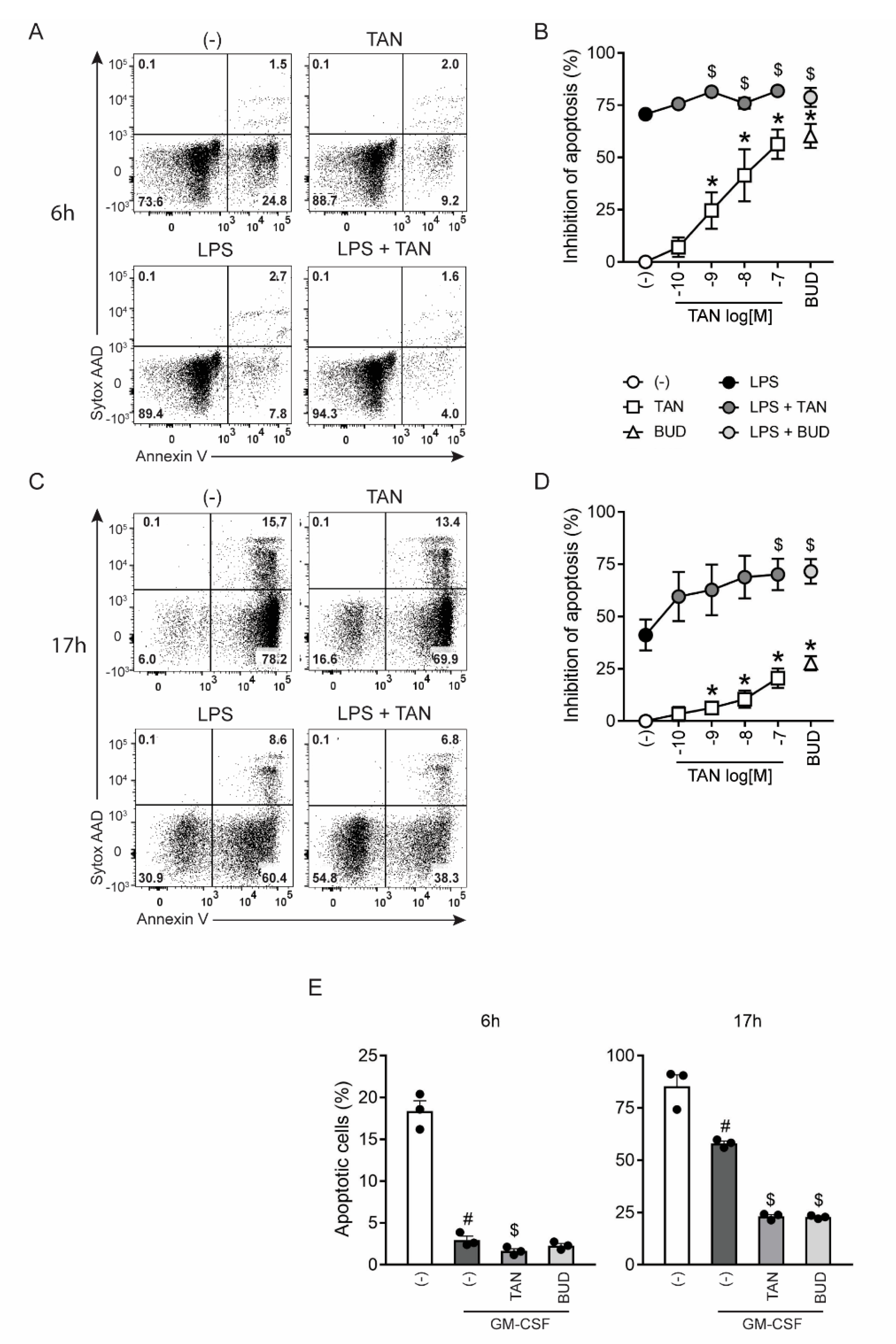
2.5. Tanimilast Promotes the Survival of Human Neutrophils
3. Materials and Methods
3.1. Drugs and Treatments
3.2. Neutrophil Isolation
3.3. Neutrophil Adhesion to HUVEC Monolayers
3.4. Degranulation Assay
3.5. NET Release
3.6. Cytokine Production
3.7. Assessment of PMN Apoptosis
3.8. Statistics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mårdh, C.K.; Root, J.; Uddin, M.; Stenvall, K.; Malmgren, A.; Karabelas, K.; Thomas, M. Targets of Neutrophil Influx and Weaponry: Therapeutic Opportunities for Chronic Obstructive Airway Disease. J. Immunol. Res. 2017, 2017, 5273201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, M.; Watz, H.; Malmgren, A.; Pedersen, F. NETopathic Inflammation in Chronic Obstructive Pulmonary Disease and Severe Asthma. Front. Immunol. 2019, 10, 47. [Google Scholar] [CrossRef]
- Maurice, D.H.; Ke, H.; Ahmad, F.; Wang, Y.; Chung, J.; Manganiello, V.C. Advances in targeting cyclic nucleotide phosphodiesterases. Nat. Rev. Drug Discov. 2014, 13, 290–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, H.; Cattani-Cavalieri, I.; Musheshe, N.; Nikolaev, V.O.; Schmidt, M. Phosphodiesterases as therapeutic targets for respiratory diseases. Pharmacol. Ther. 2019, 197, 225–242. [Google Scholar] [CrossRef] [PubMed]
- Vestbo, J.; Hurd, S.S.; Agustí, A.G.; Jones, P.W.; Vogelmeier, C.; Anzueto, A.; Barnes, P.J.; Fabbri, L.M.; Martinez, F.J.; Nishimura, M.; et al. Global strategy for the diagnosis, management, and prevention of chronic obstructive pulmonary disease: GOLD executive summary. Am. J. Respir. Crit. Care Med. 2013, 187, 347–365. [Google Scholar] [CrossRef] [PubMed]
- Rhee, C.K.; Kim, D.K. Role of phosphodiesterase-4 inhibitors in chronic obstructive pulmonary disease. Korean J. Intern. Med. 2020, 35, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Facchinetti, F.; Civelli, M.; Singh, D.; Papi, A.; Emirova, A.; Govoni, M. Tanimilast, A Novel Inhaled Pde4 Inhibitor for the Treatment of Asthma and Chronic Obstructive Pulmonary Disease. Front. Pharmacol. 2021, 12, 740803. [Google Scholar] [CrossRef]
- Stellari, F.F.; Sala, A.; Ruscitti, F.; Buccellati, C.; Allen, A.; Risé, P.; Civelli, M.; Villetti, G. CHF6001 Inhibits NF-κB Activation and Neutrophilic Recruitment in LPS-Induced Lung Inflammation in Mice. Front. Pharmacol. 2019, 10, 1337. [Google Scholar] [CrossRef] [Green Version]
- Moretto, N.; Caruso, P.; Bosco, R.; Marchini, G.; Pastore, F.; Armani, E.; Amari, G.; Rizzi, A.; Ghidini, E.; De Fanti, R.; et al. CHF6001 I: A novel highly potent and selective phosphodiesterase 4 inhibitor with robust anti-inflammatory activity and suitable for topical pulmonary administration. J. Pharmacol. Exp. Ther. 2015, 352, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, E.P.; Lee, W.L.; Zemans, R.L.; Yamashita, C.; Downey, G.P. On, around, and through: Neutrophil-endothelial interactions in innate immunity. Physiology 2011, 26, 334–347. [Google Scholar] [CrossRef] [Green Version]
- Cassatella, M.A.; Östberg, N.K.; Tamassia, N.; Soehnlein, O. Biological Roles of Neutrophil-Derived Granule Proteins and Cytokines. Trends Immunol. 2019, 40, 648–664. [Google Scholar] [CrossRef]
- Jones, N.A.; Boswell-Smith, V.; Lever, R.; Page, C.P. The effect of selective phosphodiesterase isoenzyme inhibition on neutrophil function in vitro. Pulm. Pharmacol. Ther. 2005, 18, 93–101. [Google Scholar] [CrossRef]
- Braga, P.C.; Dal Sasso, M.; Culici, M.; Bianchi, T.; Guffanti, E.E. Budesonide reduces superoxide and peroxynitrite anion chemiluminescence during human neutrophil bursts. Pharmacology 2005, 75, 179–186. [Google Scholar] [CrossRef]
- Zhang, X.; Moilanen, E.; Kankaanranta, H. Beclomethasone, budesonide and fluticasone propionate inhibit human neutrophil apoptosis. Eur. J. Pharmacol. 2001, 431, 365–371. [Google Scholar] [CrossRef]
- Schleimer, R.P.; Freeland, H.S.; Peters, S.P.; Brown, K.E.; Derse, C.P. An assessment of the effects of glucocorticoids on degranulation, chemotaxis, binding to vascular endothelium and formation of leukotriene B4 by purified human neutrophils. J. Pharmacol. Exp. Ther. 1989, 250, 598–605. [Google Scholar]
- Singh, D.; Beeh, K.M.; Colgan, B.; Kornmann, O.; Leaker, B.; Watz, H.; Lucci, G.; Geraci, S.; Emirova, A.; Govoni, M.; et al. Effect of the inhaled PDE4 inhibitor CHF6001 on biomarkers of inflammation in COPD. Respir. Res. 2019, 20, 180. [Google Scholar] [CrossRef] [Green Version]
- Tonnesen, M.G.; Anderson, D.C.; Springer, T.A.; Knedler, A.; Avdi, N.; Henson, P.M. Adherence of neutrophils to cultured human microvascular endothelial cells. Stimulation by chemotactic peptides and lipid mediators and dependence upon the Mac-1, LFA-1, p150,95 glycoprotein family. J. Clin. Investig. 1989, 83, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Yin, C.; Heit, B. Armed for destruction: Formation, function and trafficking of neutrophil granules. Cell Tissue Res. 2018, 371, 455–471. [Google Scholar] [CrossRef]
- Calzetti, F.; Tamassia, N.; Arruda-Silva, F.; Gasperini, S.; Cassatella, M.A. The importance of being “pure” neutrophils. J. Allergy Clin. Immunol. 2017, 139, 352–355. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Latif, D.; Steward, M.; Macdonald, D.L.; Francis, G.A.; Dinauer, M.C.; Lacy, P. Rac2 is critical for neutrophil primary granule exocytosis. Blood 2004, 104, 832–839. [Google Scholar] [CrossRef] [Green Version]
- Kruger, P.; Saffarzadeh, M.; Weber, A.N.; Rieber, N.; Radsak, M.; von Bernuth, H.; Benarafa, C.; Roos, D.; Skokowa, J.; Hartl, D. Neutrophils: Between host defence, immune modulation, and tissue injury. PLoS Pathog. 2015, 11, e1004651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, R.C.; Wehner, N.G.; Skubitz, K.M.; Gray, B.H.; Hoidal, J.R. Proteinase 3. A distinct human polymorphonuclear leukocyte proteinase that produces emphysema in hamsters. J. Clin. Investig. 1988, 82, 1963–1973. [Google Scholar] [CrossRef] [PubMed]
- Park, J.A.; He, F.; Martin, L.D.; Li, Y.; Chorley, B.N.; Adler, K.B. Human neutrophil elastase induces hypersecretion of mucin from well-differentiated human bronchial epithelial cells in vitro via a protein kinase Cδ-mediated mechanism. Am. J. Pathol. 2005, 167, 651–661. [Google Scholar] [CrossRef]
- Okada, Y.; Watanabe, S.; Nakanishi, I.; Kishi, J.; Hayakawa, T.; Watorek, W.; Travis, J.; Nagase, H. Inactivation of tissue inhibitor of metalloproteinases by neutrophil elastase and other serine proteinases. FEBS Lett. 1988, 229, 157–160. [Google Scholar] [CrossRef] [Green Version]
- Gaggar, A.; Li, Y.; Weathington, N.; Winkler, M.; Kong, M.; Jackson, P.; Blalock, J.E.; Clancy, J.P. Matrix metalloprotease-9 dysregulation in lower airway secretions of cystic fibrosis patients. Am. J. Physiol.-Lung Cell Mol. Physiol. 2007, 293, L96–L104. [Google Scholar] [CrossRef] [Green Version]
- Hussell, T.; Lui, S.; Jagger, C.; Morgan, D.; Brand, O. The consequence of matrix dysfunction on lung immunity and the microbiome in COPD. Eur. Respir. Rev. 2018, 27, 180032. [Google Scholar] [CrossRef] [Green Version]
- Ammit, A.J. Glucocorticoid insensitivity as a source of drug targets for respiratory disease. Curr. Opin. Pharmacol. 2013, 13, 370–376. [Google Scholar] [CrossRef]
- Rennard, S.I.; Fogarty, C.; Kelsen, S.; Long, W.; Ramsdell, J.; Allison, J.; Mahler, D.; Saadeh, C.; Siler, T.; Snell, P.; et al. The safety and efficacy of infliximab in moderate to severe chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2007, 175, 926–934. [Google Scholar] [CrossRef]
- Jorch, S.K.; Kubes, P. An emerging role for neutrophil extracellular traps in noninfectious disease. Nat. Med. 2017, 23, 279–287. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Lefrançais, E.; Mallavia, B.; Zhuo, H.; Calfee, C.S.; Looney, M.R. Maladaptive role of neutrophil extracellular traps in pathogen-induced lung injury. JCI Insight 2018, 3, e98178. [Google Scholar] [CrossRef] [Green Version]
- Dicker, A.J.; Crichton, M.L.; Pumphrey, E.G.; Cassidy, A.J.; Suarez-Cuartin, G.; Sibila, O.; Furrie, E.; Fong, C.J.; Ibrahim, W.; Brady, G.; et al. Neutrophil extracellular traps are associated with disease severity and microbiota diversity in patients with chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2018, 141, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Wright, T.K.; Gibson, P.G.; Simpson, J.L.; McDonald, V.M.; Wood, L.G.; Baines, K.J. Neutrophil extracellular traps are associated with inflammation in chronic airway disease. Respirology 2016, 21, 467–475. [Google Scholar] [CrossRef] [Green Version]
- Tamassia, N.; Bianchetto-Aguilera, F.; Arruda-Silva, F.; Gardiman, E.; Gasperini, S.; Calzetti, F.; Cassatella, M.A. Cytokine production by human neutrophils: Revisiting the “dark side of the moon”. Eur. J. Clin. Investig. 2018, 48 (Suppl. S2), e12952. [Google Scholar] [CrossRef]
- Strandberg, K.; Blidberg, K.; Sahlander, K.; Palmberg, L.; Larsson, K. Effect of formoterol and budesonide on chemokine release, chemokine receptor expression and chemotaxis in human neutrophils. Pulm. Pharmacol. Ther. 2010, 23, 316–323. [Google Scholar] [CrossRef] [Green Version]
- Gianello, V.; Salvi, V.; Parola, C.; Moretto, N.; Facchinetti, F.; Civelli, M.; Villetti, G.; Bosisio, D.; Sozzani, S. The PDE4 inhibitor CHF6001 modulates pro-inflammatory cytokines, chemokines and Th1- and Th17-polarizing cytokines in human dendritic cells. Biochem. Pharmacol. 2019, 163, 371–380. [Google Scholar] [CrossRef]
- Barnes, P.J. Mediators of chronic obstructive pulmonary disease. Pharmacol. Rev. 2004, 56, 515–548. [Google Scholar] [CrossRef] [Green Version]
- Edwards, M.R.; Facchinetti, F.; Civelli, M.; Villetti, G.; Johnston, S.L. Anti-inflammatory effects of the novel inhaled phosphodiesterase type 4 inhibitor CHF6001 on virus-inducible cytokines. Pharmacol. Res. Perspect. 2016, 4, e00202. [Google Scholar] [CrossRef]
- Pérez-Figueroa, E.; Álvarez-Carrasco, P.; Ortega, E.; Maldonado-Bernal, C. Neutrophils: Many Ways to Die. Front. Immunol. 2021, 12, 631821. [Google Scholar] [CrossRef]
- Colotta, F.; Re, F.; Polentarutti, N.; Sozzani, S.; Mantovani, A. Modulation of granulocyte survival and programmed cell death by cytokines and bacterial products. Blood 1992, 80, 2012–2020. [Google Scholar] [CrossRef] [Green Version]
- Parkkonen, J.; Hasala, H.; Moilanen, E.; Giembycz, M.A.; Kankaanranta, H. Phosphodiesterase 4 inhibitors delay human eosinophil and neutrophil apoptosis in the absence and presence of salbutamol. Pulm. Pharmacol. Ther. 2008, 21, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Totani, L.; Amore, C.; Piccoli, A.; Dell’Elba, G.; Di Santo, A.; Plebani, R.; Pecce, R.; Martelli, N.; Rossi, A.; Ranucci, S.; et al. Type-4 Phosphodiesterase (PDE4) Blockade Reduces NETosis in Cystic Fibrosis. Front. Pharmacol. 2021, 12, 702677. [Google Scholar] [CrossRef] [PubMed]
- Lima, K.M.; Vago, J.P.; Caux, T.R.; Negreiros-Lima, G.L.; Sugimoto, M.A.; Tavares, L.P.; Arribada, R.G.; Carmo, A.A.F.; Galvão, I.; Costa, B.R.C.; et al. The resolution of acute inflammation induced by cyclic AMP is dependent on annexin A1. J. Biol. Chem. 2017, 292, 13758–13773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, L.P.; Lopes, F.; Silva, D.M.; Tavares, L.P.; Vieira, A.T.; Rezende, B.M.; Carmo, A.F.; Russo, R.C.; Garcia, C.C.; Bonjardim, C.A.; et al. PDE4 inhibition drives resolution of neutrophilic inflammation by inducing apoptosis in a PKA-PI3K/Akt-dependent and NF-kappaB-independent manner. J. Leukoc. Biol. 2010, 87, 895–904. [Google Scholar] [CrossRef]
- Greenlee-Wacker, M.C. Clearance of apoptotic neutrophils and resolution of inflammation. Immunol. Rev. 2016, 273, 357–370. [Google Scholar] [CrossRef] [Green Version]
- Sanz, M.J.; Cortijo, J.; Taha, M.A.; Cerdá-Nicolás, M.; Schatton, E.; Burgbacher, B.; Klar, J.; Tenor, H.; Schudt, C.; Issekutz, A.C.; et al. Roflumilast inhibits leukocyte-endothelial cell interactions, expression of adhesion molecules and microvascular permeability. Br. J. Pharmacol. 2007, 152, 481–492. [Google Scholar] [CrossRef]
- Crampton, S.P.; Davis, J.; Hughes, C.C. Isolation of human umbilical vein endothelial cells (HUVEC). J. Vis. Exp. 2007, 3, 183. [Google Scholar] [CrossRef] [Green Version]
- Del Prete, A.; Martínez-Muñoz, L.; Mazzon, C.; Toffali, L.; Sozio, F.; Za, L.; Bosisio, D.; Gazzurelli, L.; Salvi, V.; Tiberio, L.; et al. The atypical receptor CCRL2 is required for CXCR2-dependent neutrophil recruitment and tissue damage. Blood 2017, 130, 1223–1234. [Google Scholar] [CrossRef]
- Pulli, B.; Ali, M.; Forghani, R.; Schob, S.; Hsieh, K.L.; Wojtkiewicz, G.; Linnoila, J.J.; Chen, J.W. Measuring myeloperoxidase activity in biological samples. PLoS ONE 2013, 8, e67976. [Google Scholar] [CrossRef] [Green Version]
- Day, J.A.; Cohen, S.M. Investigating the selectivity of metalloenzyme inhibitors. J. Med. Chem. 2013, 56, 7997–8007. [Google Scholar] [CrossRef] [Green Version]
- Maugeri, N.; Campana, L.; Gavina, M.; Covino, C.; De Metrio, M.; Panciroli, C.; Maiuri, L.; Maseri, A.; D’Angelo, A.; Bianchi, M.E.; et al. Activated platelets present high mobility group box 1 to neutrophils, inducing autophagy and promoting the extrusion of neutrophil extracellular traps. J. Thromb. Haemost. 2014, 12, 2074–2088. [Google Scholar] [CrossRef]
- Scapini, P.; Carletto, A.; Nardelli, B.; Calzetti, F.; Roschke, V.; Merigo, F.; Tamassia, N.; Pieropan, S.; Biasi, D.; Sbarbati, A.; et al. Proinflammatory mediators elicit secretion of the intracellular B-lymphocyte stimulator pool (BLyS) that is stored in activated neutrophils: Implications for inflammatory diseases. Blood 2005, 105, 830–837. [Google Scholar] [CrossRef]
- Chong, J.; Leung, B.; Poole, P. Phosphodiesterase 4 inhibitors for chronic obstructive pulmonary disease. Cochrane Database Syst. Rev. 2017, 9, CD002309. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Stimulus | IC50 (nM) |
|---|---|---|
| Adhesion to endothelium | fMLF | 1.97 |
| Degranulation: | ||
| NE | fMLF | 49.2 # |
| MPO | fMLF | 48.8 § |
| MMP | fMLF | 9.80 #,§ |
| Cytokine secretion: | ||
| TNF-α | R848 | 53.3 ° |
| LPS | 145 * | |
| CXCL8 | R848 | 8.36 ° |
| LPS | 4.20 * | |
| Spontaneous apoptosis inhibition (6 h) | NONE | 39.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schioppa, T.; Nguyen, H.O.; Salvi, V.; Maugeri, N.; Facchinetti, F.; Villetti, G.; Civelli, M.; Gaudenzi, C.; Passari, M.; Sozio, F.; et al. The PDE4 Inhibitor Tanimilast Restrains the Tissue-Damaging Properties of Human Neutrophils. Int. J. Mol. Sci. 2022, 23, 4982. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094982
Schioppa T, Nguyen HO, Salvi V, Maugeri N, Facchinetti F, Villetti G, Civelli M, Gaudenzi C, Passari M, Sozio F, et al. The PDE4 Inhibitor Tanimilast Restrains the Tissue-Damaging Properties of Human Neutrophils. International Journal of Molecular Sciences. 2022; 23(9):4982. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094982
Chicago/Turabian StyleSchioppa, Tiziana, Hoang Oanh Nguyen, Valentina Salvi, Norma Maugeri, Fabrizio Facchinetti, Gino Villetti, Maurizio Civelli, Carolina Gaudenzi, Mauro Passari, Francesca Sozio, and et al. 2022. "The PDE4 Inhibitor Tanimilast Restrains the Tissue-Damaging Properties of Human Neutrophils" International Journal of Molecular Sciences 23, no. 9: 4982. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23094982