
Responses of Growth, Oxidative Injury and Chloroplast Ultrastructure in Leaves of Lolium perenne and Festuca arundinacea to Elevated O3 Concentrations

Abstract
:1. Introduction
2. Results
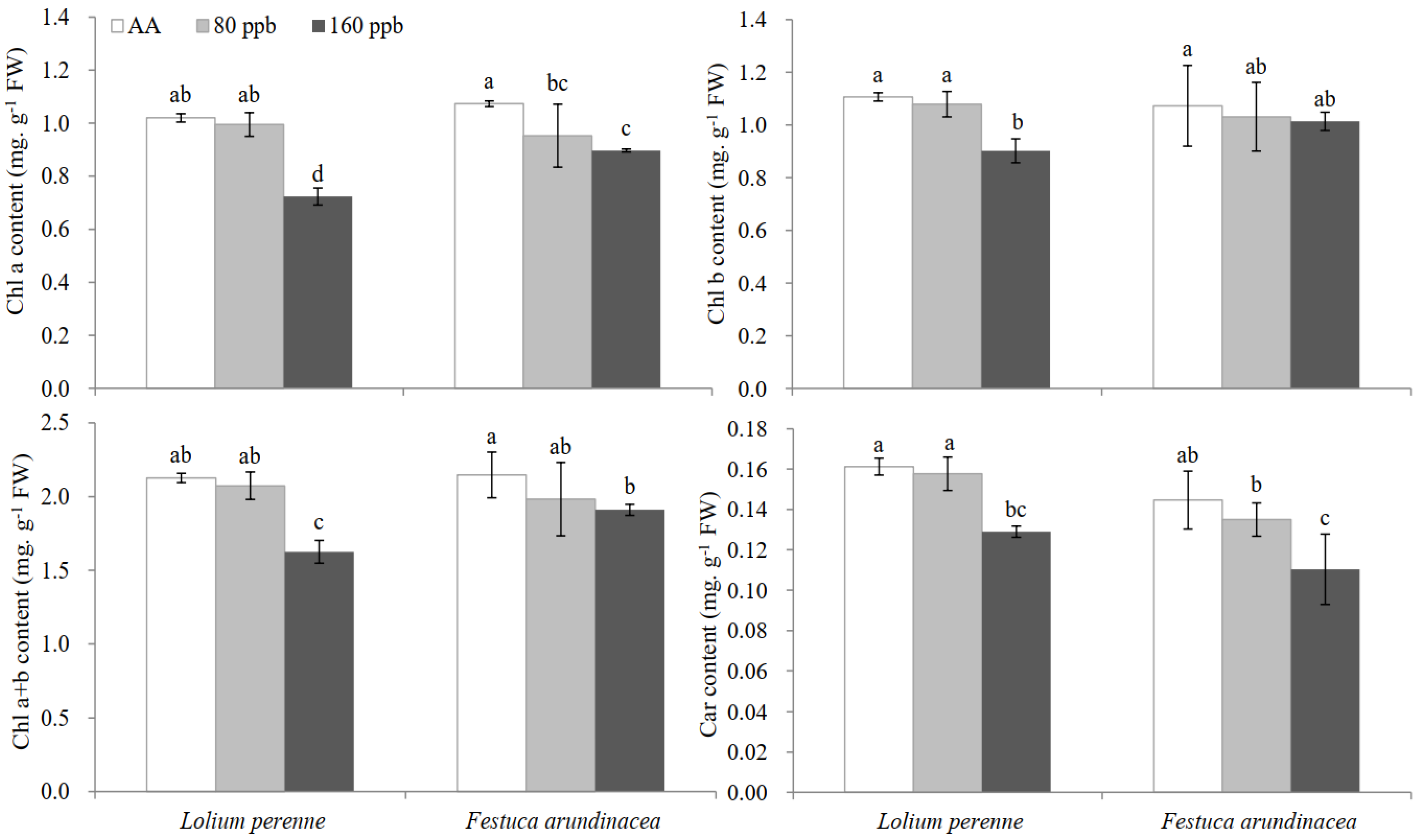
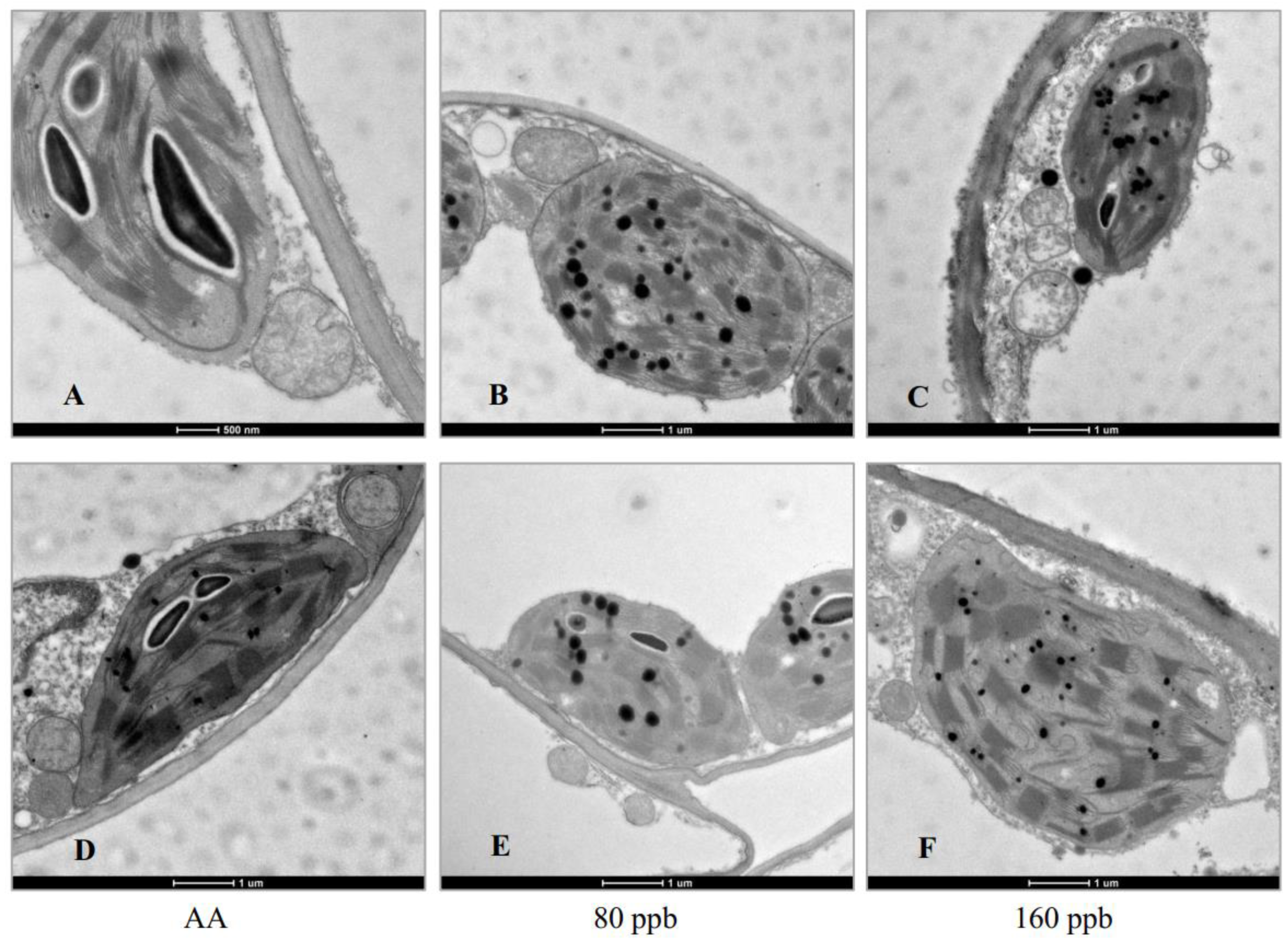
2.1. Visible Injury, Pigment Contents and Chloroplast Ultrastructure
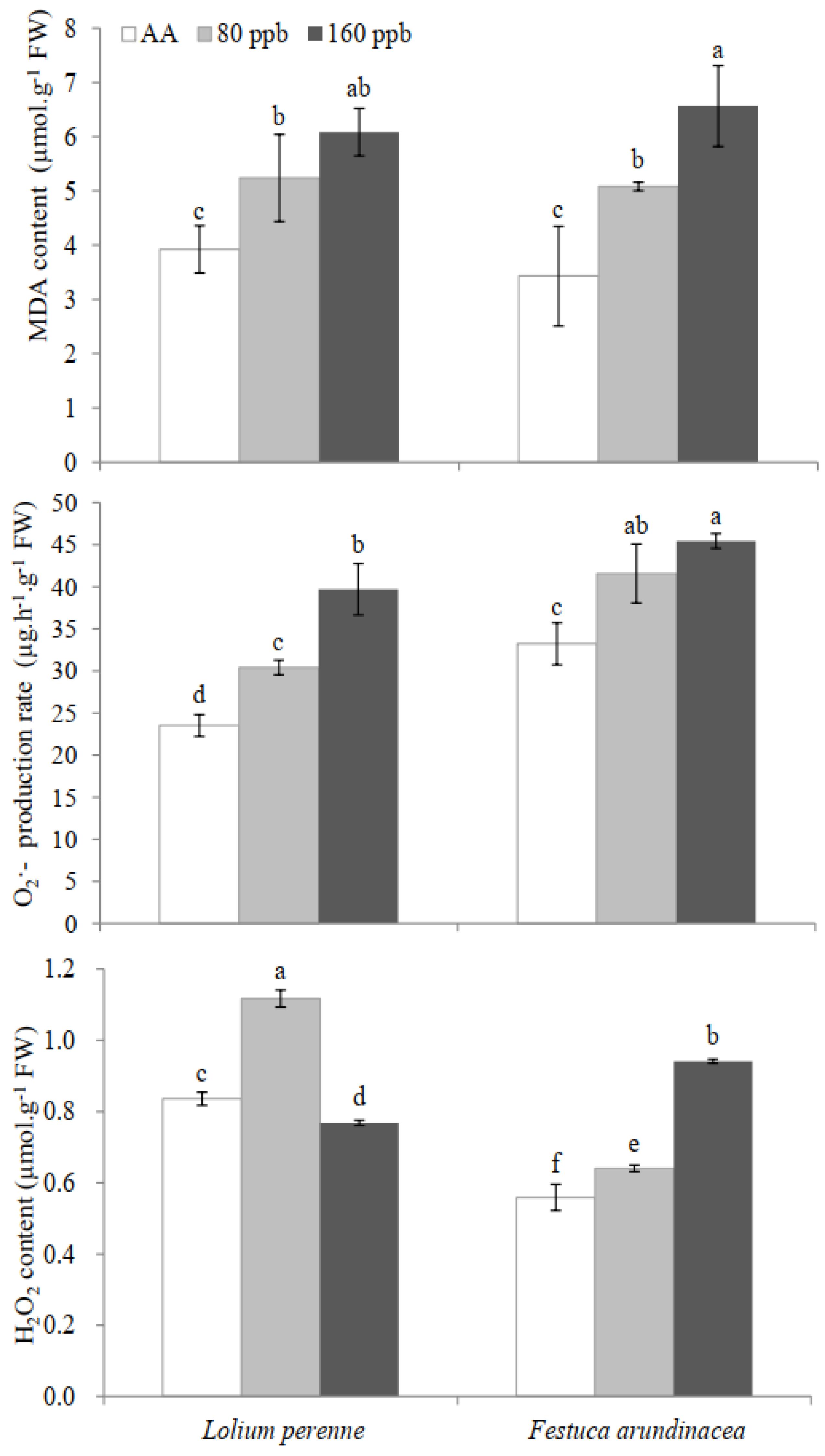
2.2. Oxidative Injury
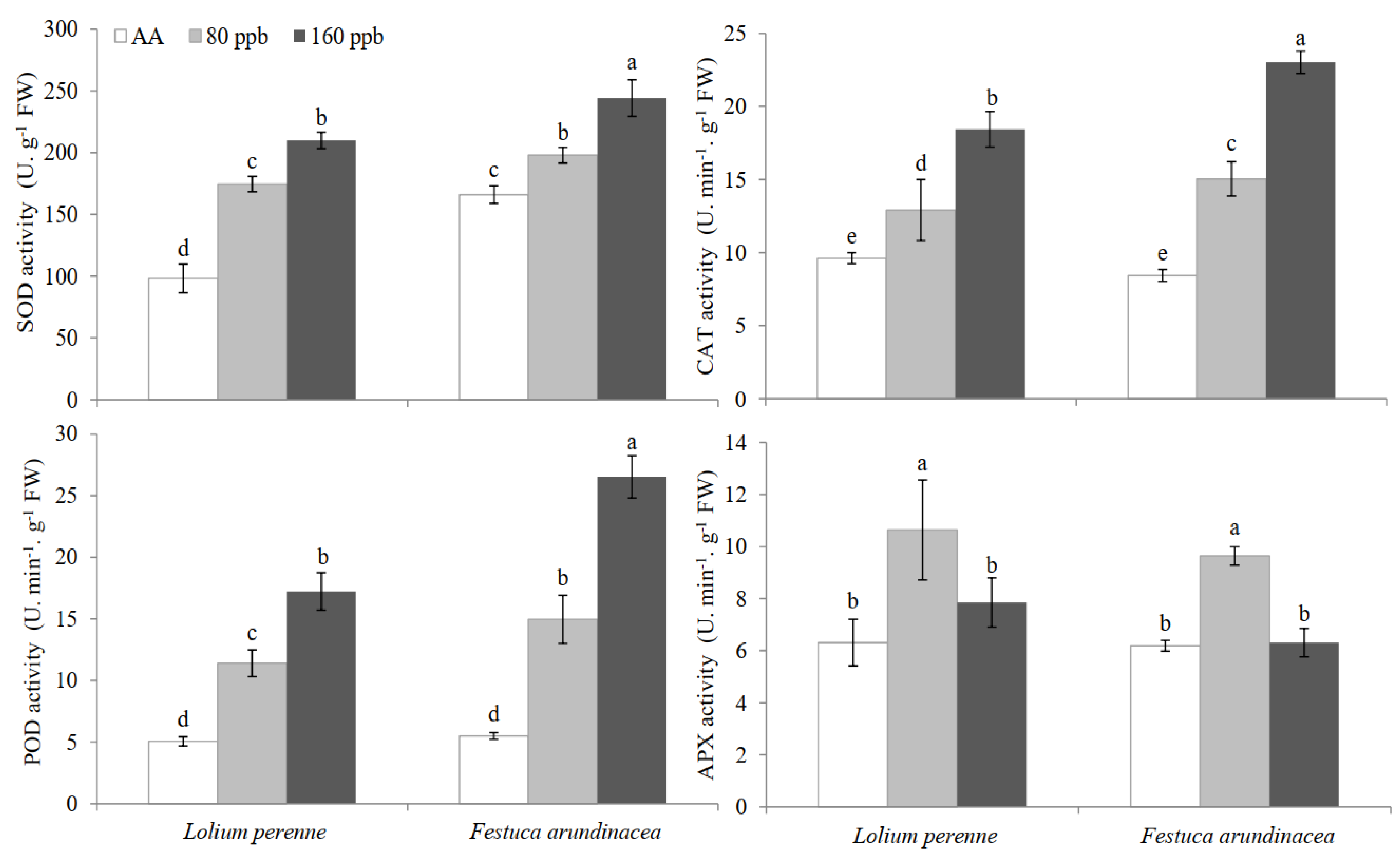
2.3. Changes in Antioxidative Enzyme Activities
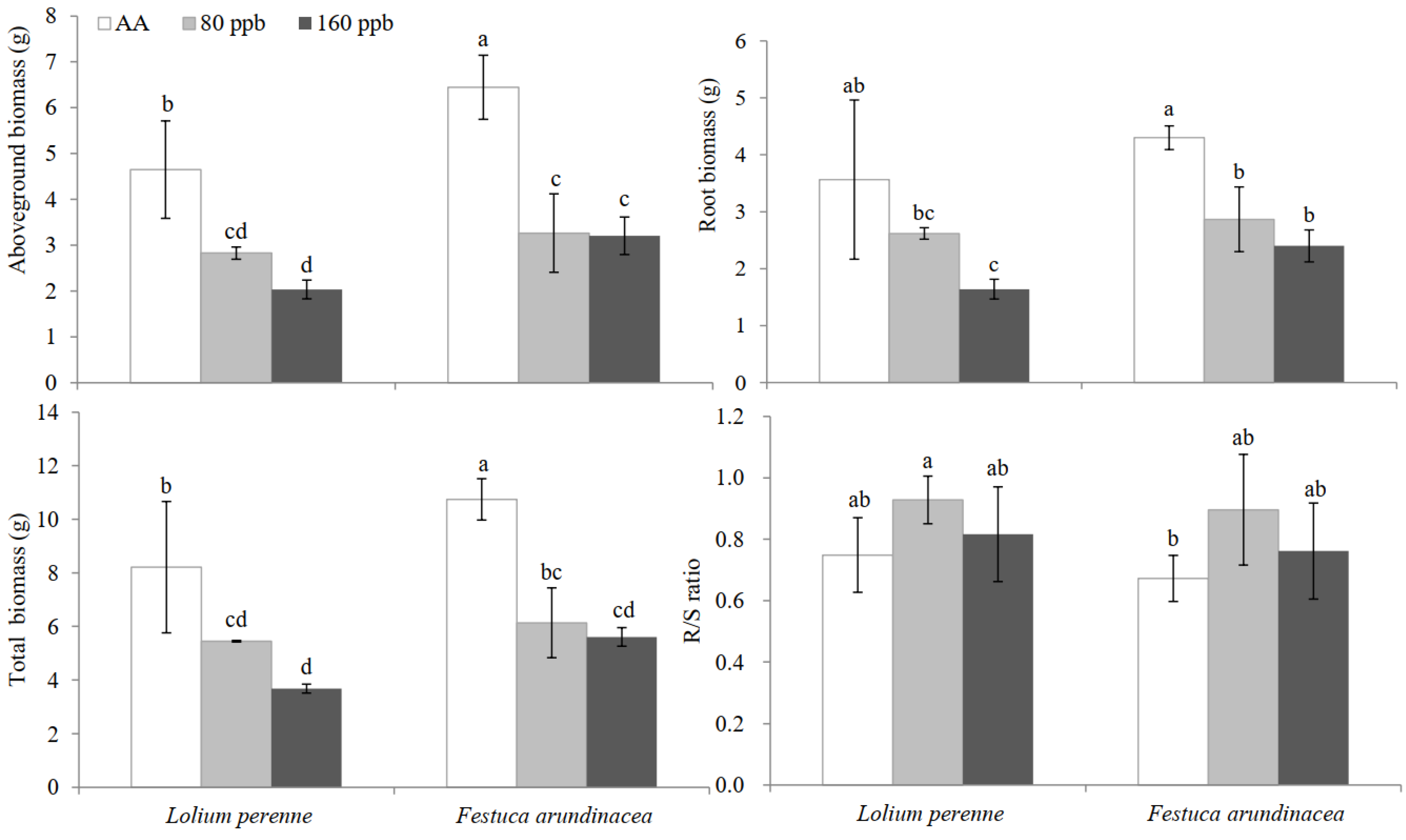
2.4. Growth Response
3. Discussion
3.1. Visible Injury and Growth Characteristics
3.2. Pigment Content and Chloroplast Ultrastructure Changes
3.3. Oxidative Injury and Antioxidative Metabolism
4. Materials and Methods
4.1. Study Site
4.2. Experimental Design
4.3. Plant Materials and Treatments
4.4. Measurements of Growth and Photosynthetic Pigment Contents
4.5. Evaluation of Oxidative Injury
4.6. Determination of Antioxidant Enzyme Activities
4.7. Transmission Electron Microscopy Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Dingenen, R.; Dentener, F.J.; Raes, F.; Krol, M.; Emberson, L.; Cofala, J. The global impact of ozone on agricultural crop yields under current and future air quality legislation. Atmos. Environ. 2009, 43, 604–618. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Yendrek, C.R.; Sitch, S.; Collins, W.J.; Emberson, L.D. The effects of tropospheric ozone on net primary productivity and implications of climate change. Annu. Rev. Plant Biol. 2012, 63, 637–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; He, X.; Chen, W.; Huang, Y.; Zhao, Y.; Li, B. Differential sensitivity of four urban tree species to elevated O3. Urban For. Urban Gree. 2015, 14, 1166–1173. [Google Scholar] [CrossRef]
- Liu, N.; Lin, W.; Ma, J.; Xu, W.; Xu, X. Seasonal variation in surface ozone and its regional characteristics at global atmosphere watch stations in China. J. Environ. Sci. 2019, 77, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Wang, P.; Zhang, J.; Zhang, H.; Tang, Y.; Hu, J.; Ying, Q. Spatial-temporal variations and source contributions to forest ozone exposure in China. Sci. Total Environ. 2019, 674, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Hu, E.; Wang, X.; Jiang, L.; Liu, X. Ground-level O3 pollution and its impacts on food crops in China: A Review. Environ. Pollut. 2015, 199, 42–48. [Google Scholar] [CrossRef]
- Dai, L.; Li, P.; Shang, B.; Liu, S.; Yang, A.; Wang, Y.; Feng, Z. Differential responses of peach (Prunus persica) seedlings to elevated ozone are related with leaf mass per area, antioxidant enzymes activity rather than stomatal conductance. Environ. Pollut. 2017, 227, 380–388. [Google Scholar] [CrossRef]
- Iyer, N.J.; Tang, Y.; Mahalingam, R. Physiological, biochemical and molecular responses to a combination of drought and ozone in Medicago truncatula. Plant Cell Environ. 2013, 36, 706–720. [Google Scholar] [CrossRef]
- Xu, S.; He, X.; Chen, W.; Su, D.; Huang, Y. Elevated CO2 ameliorated the adverse effect of elevated O3 in previous-year and current-year needles of Pinus tabulaeformis in urban area. Bull. Environ. Contam. Toxicol. 2014, 92, 733–737. [Google Scholar] [CrossRef]
- Liu, X.; Sui, L.; Huang, Y.; Geng, C.; Yin, B. Physiological and visible injury responses in different growth stages of winter wheat to ozone stress and the protection of spermidine. Atmos. Pollut. Res. 2015, 6, 596–604. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.J.; Gharibeshghi, A.; Mewis, I.; Förster, N.; Beck, W.; Ulrichs, C. Plant responses to ozone: Effects of different ozone exposure durations on plant growth and biochemical quality of Brassica campestris L. ssp. Chinensis. Sci. Hortic. 2020, 262, 108921. [Google Scholar] [CrossRef]
- Shang, B.; Feng, Z.; Gao, F.; Calatayud, V. The ozone sensitivity of five poplar clones is not related to stomatal conductance, constitutive antioxidant levels and morphology of leaves. Sci. Total Environ. 2020, 699, 134402. [Google Scholar] [CrossRef] [PubMed]
- Hayes, F.; Mills, G.; Ashmore, M. Effects of ozone on inter-and intra-species competition and photosynthesis in mesocosms of Lolium perenne and Trifolium repens. Environ. Pollut. 2009, 157, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Pasqualetti, C.B.; Sandrin, C.Z.; Pedroso, A.N.V.; Domingos, M.; Figueiredo-Ribeiro, R.C.L. Fructans, ascorbate peroxidase, and hydrogen peroxide in ryegrass exposed to ozone under contrasting meteorological conditions. Environ. Sci. Pollut. Res. 2015, 22, 4771–4779. [Google Scholar] [CrossRef] [PubMed]
- Calatayud, V.; Cerveró, J.; Calvo, E.; García-Breijo, F.-J.; Reig-Armiñana, J.; Sanz, M.J. Responses of evergreen and deciduous Quercus species to enhanced ozone levels. Environ. Pollut. 2011, 159, 55–63. [Google Scholar] [CrossRef]
- Fares, S.; Vargas, R.; Detto, M.; Goldstein, A.H.; Karlik, J.; Paoletti, E.; Vitale, M. Tropospheric ozone reduces carbon assimilation in trees: Estimates from analysis of continuous flux measurements. Glob. Chang. Biol. 2013, 19, 2427–2443. [Google Scholar] [CrossRef]
- Kitao, M.; Yasuda, Y.; Kominami, Y.; Yamanoi, K.; Komatsu, M.; Miyama, T.; Mizoguchi, Y.; Kitaoka, S.; Yazaki, K.; Tobita, H.; et al. Increased phytotoxic O3 dose accelerates autumn senescence in an O3-sensitive beech forest even under the present-level O3. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- De Marco, A.; Vitale, M.; Popa, I.; Anav, A.; Badea, O.; Silaghi, D.; Leca, S.; Screpanti, A.; Paoletti, E. Ozone exposure affects tree defoliation in a continental climate. Sci. Total Environ. 2017, 596–597, 396–404. [Google Scholar] [CrossRef]
- Kangasjarvi, J.; Jaspers, P.; Kollist, H. Signalling and cell death in ozone exposed plants. Plant Cell Environ. 2005, 28, 1021–1036. [Google Scholar] [CrossRef]
- Gao, F.; Catalayud, V.; Garcia-Breijo, F.; Reig-Arminana, J.; Feng, Z. Effects of elevated ozone on physiological, anatomical and ultrastructureal characteristics of four common urban tree species in China. Ecol. Indic. 2016, 67, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Krupa, S.; McGrath, M.T.; Andersen, C.P.; Booker, F.L.; Burkey, K.O.; Chappelka, A.H.; Chevone, B.I.; Pell, E.J.; Zilinskas, B.A. Ambient ozone and plant health. Plant Dis. 2001, 85, 4–12. [Google Scholar] [CrossRef] [Green Version]
- Hayes, F.; Williamson, J.; Mills, G. Species-specific responses to ozone and drought in six deciduous trees. Water Air Soil Pollut. 2015, 226, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Heagle, A.S.; Mclughlin, M.R.; Miller, J.E.; Joyner, R.L.; Spruill, S.E. Adaptation of a white clover population to ozone stress. New Phytol. 1991, 119, 61–68. [Google Scholar] [CrossRef]
- Su, H.; Zhou, M.; Xu, H.; Zhang, X.; Li, Y.; Xiang, B. Photosynthesis and biochemical responses to elevated O3 in Plantago major and Sonchus oleraceus growing in a lowland habitat of northern China. J. Environ. Sci. 2017, 53, 113–121. [Google Scholar] [CrossRef]
- Xu, S.; Li, J.; Zhang, X.; Wei, H.; Cui, L. Effects of heat acclimation pretreatment on changes of membrane lipid peroxidation, antioxidant metabolites, and ultrastructure of chloroplasts in two cool-season turfgrass species under heat stress. Environ. Exp. Bot. 2006, 56, 274–285. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, B. Effects of calcium on antioxidant activities and water relations associated with heat tolerance in two cool-season grasses. J. Exp. Bot. 2001, 355, 341–349. [Google Scholar] [CrossRef]
- Plazek, A.; Hura, K.; Rapacz, H. The influence of ozone fumigation on metabolic efficiency and plant resistance to fungal pathogens. J. Appl. Bot. 2001, 75, 8–13. [Google Scholar]
- Guidi, L.; Di Cagno, R.; Soldatini, G. Screening of bean cultivars for their response to ozone as evaluated by visible symptoms and leaf chlorophyll fluorescence. Environ. Pollut. 2000, 107, 349–355. [Google Scholar] [CrossRef]
- Zhang, W.; Feng, Z.; Wang, X.; Niu, J. Responses of native broadleaved woody species to elevated ozone in subtropical China. Environ. Pollut. 2012, 163, 149–157. [Google Scholar] [CrossRef]
- Burkey, K.O.; Carter, T.E. Foliar resistance to ozone injury in the genetic base of U.S. and Canadian soybean and prediction of resistance in descendent cultivars using coefficient of parentage. Field Crops Res. 2009, 111, 207–217. [Google Scholar] [CrossRef]
- Feng, Z.; Sun, J.; Wan, W.; Hu, E.; Calatayud, V. Evidence of widespread ozone-induced visible injury on plants in Beijing, China. Environ. Pollut. 2014, 193, 296–301. [Google Scholar] [CrossRef]
- Díaz-de-Quijano, M.; Schaub, M.; Bassin, S.; Volk, M.; Penuelas, J. Ozone visible symptoms and reduced root biomass in the subalpine species Pinus uncinata after two years of free-air ozone fumigation. Environ. Pollut. 2012, 169, 250–257. [Google Scholar] [CrossRef]
- Ping, Q.; Xu, S.; Li, J.; He, X.; Chen, W.; Huang, Y. Ecophysiological responses of turf-type white clover (Trifolium repens) to elevated O3 concentration. Chin. J. Ecol. 2017, 36, 1234–1242. [Google Scholar]
- Zhao, Y.; Xu, S.; He, X.; Chen, W.; Li, M.; Zhang, N.; Fu, W. Physiological responses of three cool-season types of turfgrass to elevated concentrations. Chin. J. Ecol. 2014, 33, 3203–3208. [Google Scholar]
- Sawada, H.; Kohno, Y. Differential Ozone Sensitivity of Rice Cultivars as Indicated by Visible Injury and Grain Yield. Plant Biol. 2009, 11, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Vollsnes, A.V.; Kruse, O.M.O.; Eriksen, A.B.; Oxaal, U.; Futsaether, C.M. In vivo root growth dynamics of ozone exposed Trifolium subterraneum. Environ. Exp. Bot. 2010, 69, 183–188. [Google Scholar] [CrossRef]
- Menéndez, A.I.; Gundel, P.E.; Lores, L.M.; Martínez-Ghersa, M.A. Assessing the impacts of intra- and interspecific competition between Triticum aestivum and Trifolium repens on the species’ responses to ozone. Botany 2017, 95, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Mills, G.; Hayes, F.; Simpson, D.; Emberson, L.; Norris, D.; Harmens, H.; Buker, P. Evidence of widespread effects of ozone on crops and semi-natural vegetation in Europe (1990–2006) in relation to AOT40-and flux-based risk maps. Glob. Chang. Biol. 2011, 17, 592–613. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Li, B.; Li, P.; He, X.; Chen, W.; Yan, K.; Li, Y.; Wang, Y. Soil high Cd exacerbates the adverse impact of elevated O3 on Populus alba ‘Berolinensis’. Ecotox. Environ. Safe. 2019, 174, 35–42. [Google Scholar] [CrossRef]
- Zhang, L.; Hoshika, Y.; Carrari, E.; Badea, O.; Paoletti, E. Ozone risk assessment is affected by nutrient availability: Evidence from a simulation experiment under free air controlled exposure (FACE). Environ. Pollut. 2018, 238, 812–822. [Google Scholar] [CrossRef]
- Agathokleous, E.; Belz, R.G.; Kitao, M.; Koike, T.; Calabrese, E.J. Does the root to shoot ratio show a hormetic response to stress? An ecological and environmental perspective. J. For. Res. 2019, 30, 1569–1580. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Catalayud, V.; Paoletti, E.; Hoshika, Y.; Feng, Z. Water stress mitigates the negative effects of ozone on photosynthesis and biomass in poplar plants. Environ. Pollut. 2017, 230, 268–279. [Google Scholar] [CrossRef]
- Li, P.; Zhou, H.; Xu, Y.; Shang, B.; Feng, Z. The effects of elevated ozone on the accumulation and allocation of poplar biomass depend strongly on water and nitrogen availability. Sci. Total Environ. 2019, 665, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Fangmeier, A.; Grüters, U.; Hertstein, U.; Sandhage-Hofmann, A.; Vermehren, B.; Jäger, H.-J. Effects of elevated CO2 nitrogen supply and tropospheric ozone on spring wheat. I. Growth and yield. Environ. Pollut. 1996, 91, 381–390. [Google Scholar] [CrossRef]
- Landolt, W.; Buhlmann, U.; Bleuler, P.; Bucher, J.B. Ozone exposure-response relationships for biomass and root/shoot ratio of beech (Fagus sylvatica), ash (Fraxinus excelsior), Norway spruce (Picea abies) and Scots pine (Pinus sylvestris). Environ. Pollut. 2000, 109, 473–478. [Google Scholar] [CrossRef]
- Matyssek, R.; Sandermann, H.; Wieser, G.; Booker, F.; Cieslik, S.; Musselman, R.; Ernst, D. The challenge of making ozone risk assessment for forest trees moremechanistic. Environ. Pollut. 2008, 156, 567–582. [Google Scholar] [CrossRef]
- Dolker, T.; Mukherjee, A.; Agrawal, S.B.; Agrawal, M. Responses of a semi-natural grassland community of tropical region to elevated ozone: An assessment of soil dynamics and biomass accumulation. Sci. Total Environ. 2020, 718, 137141. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yin, R.; Shang, B.; Agathokleous, E.; Zhou, H.; Feng, Z. Interactive effects of ozone exposure and nitrogen addition on tree root traits and biomass allocation pattern: An experimental case study and a literature meta-analysis. Sci. Total Environ. 2020, 710, 136379. [Google Scholar] [CrossRef]
- Eckhardt, U.; Grimm, B.; Hörtensteiner, S. Recent advances in chlorophyll biosynthesis and breakdown in higher plants. Plant Mol. Biol. 2004, 56, 1–14. [Google Scholar] [CrossRef]
- Kobayashi, K.; Fujii, S.; Sasaki, D.; Baba, S.; Ohta, H.; Masuda, T.; Wada, H. Transcriptional regulation of thylakoid galactolipid biosynthesis coordinated with chlorophyll biosynthesis during the development of chloroplasts in Arabidopsis. Front. Plant Sci. 2014, 5, 272. [Google Scholar] [CrossRef] [Green Version]
- Leitao, L.; Moret, J.J.; Biolley, J.P. Changes in PEP carboxylase, rubisco and rubisco activase mRNA levels from maize (Zea mays) exposed to a chronic ozone stress. Biol. Res. 2007, 40, 137–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calatayud, A.; Barreno, E. Response to ozone in two lettuce varieties on chlorophyll a fluorescence, photosynthetic pigments and lipid peroxidation. Plant Physiol. Biochem. 2004, 42, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Ommen, O.; Donnelly, A.; Vanhoutvin, S.; Van Oijen, M.; Manderscheid, R. Chlorophyll content of spring wheat flag leaves grown under elevated CO2 concentrations and other environmental stresses within the ‘ESPACE-wheat’ project. Eur. J. Agron. 1999, 10, 197–203. [Google Scholar] [CrossRef]
- Yan, K.; Chen, W.; He, X.; Zhang, G.; Xu, S.; Wang, L. Responses of photosynthesis, lipid peroxidation and antioxidant system in leaves of Quercus mongolica to elevated O3. Environ. Exp. Bot. 2010, 69, 198–204. [Google Scholar] [CrossRef]
- Harmens, H.; Hayes, F.; Sharps, K.; Mills, G.; Calatayud, V. Leaf traits and photosynthetic responses of Betula pendula saplings to a range of ground-level ozone concentrations at a range of nitrogen loads. J. Plant Physiol. 2017, 211, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Calatayud, V.; García-Breijo, F.J.; Cervero, J.; Reig-Armiñana, J.; Sanz, M.J. Physiological, anatomical and biomass partitioning responses to ozone in the Mediterranean endemic plant Lamottea dianae. Ecotoxicol. Environ. Safe 2011, 74, 1131–1138. [Google Scholar] [CrossRef]
- Kivimäenpää, M.; Sutinen, S.; Calatayud, V.; Sanz, M.J. Visible and microscopic needle alterations of mature Aleppo Pine (Pinus halepensis) trees growing on anozone gradient in eastern Spain. Tree Physiol. 2010, 30, 541–554. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.A.; Chaurasia, M.; Gupta, V.; Agrawal, M.; Agrawal, S.B. Responses of Zea mays L. cultivars ‘Buland’ and ‘Prakash’ to an antiozonant ethylene diurea grown under ambient and elevated levels of ozone. Acta Physiol. Plant. 2018, 40, 92. [Google Scholar] [CrossRef]
- Podda, A.; Pisuttu, C.; Hoshika, Y.; Pellegrini, E.; Carrari, E.; Lorenzini, G.; Nali, C.; Cotrozzi, L.; Zhang, L.; Baraldi, R.; et al. Can nutrient fertilization mitigate the effects of ozone exposure on an ozone-sensitive poplar clone? Sci. Total Environ. 2019, 657, 340–350. [Google Scholar] [CrossRef]
- Rao, M.V.; Davis, K. The physiology of ozone induced cell death. Planta 2001, 213, 682–690. [Google Scholar] [CrossRef]
- Lu, T.; He, X.; Chen, W.; Yan, K.; Zhao, T. Effects of elevated O3 and/or elevated CO2 on lipid peroxidation and antioxidant systems in Ginkgo biloba leaves. Bull. Environ. Contam. Toxicol. 2009, 83, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Booker, F.; Muntifering, R.; McGrath, M.; Burkey, K.; Decoteau, D.; Fiscus, E.; Manning, W.; Krupa, S.; Chappelka, A.; Grantz, D. The ozone component of global change: Potential effects on agricultural and horticultural plant yield, product quality and interactions with invasive species. J. Integr. Plant Biol. 2009, 51, 337–351. [Google Scholar] [CrossRef]
- Scalet, M.; Federico, R.; Guido, M.C.; Manes, F. Peroxidase activity and polyamine changes in response to ozone and simulated acid rain in Aleppo pine needles. Environ. Exp. Bot. 1995, 35, 417–425. [Google Scholar] [CrossRef]
- Xu, S.; He, X.Y.; Du, Z.; Chen, W.; Li, B.; Li, Y.; Li, M.H.; Schaub, M. Tropospheric ozone and cadmium do not have interactive effects on growth, photosynthesis and mineral nutrients of Catalpa ovata seedlings in the urban areas of Northeast China. Sci. Total Environ. 2020, 704, 135307. [Google Scholar] [CrossRef]
- Xu, W.; He, X.; Chen, W.; Hu, J.; Wen, H. Responses of Shenyang urban tree phenology to climate warming. Chin. J. Appl. Ecol. 2006, 17, 1777–1781. [Google Scholar]
- Xu, S.; Fu, W.; He, X.; Chen, W.; Zhang, W.; Li, B.; Huang, Y. Drought alleviated the negative effects of elevated O3 on Lonicera maackii in urban area. Bull. Environ. Contam. Toxicol. 2017, 99, 648–653. [Google Scholar] [CrossRef] [PubMed]
- Heagle, A.S.; Body, D.E.; Heck, W.W. An open top field chamber to assess the impact of air pollution on plants. J. Environ. Qual. 1973, 2, 365–368. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic membranes. Method Enzym. 1987, 148, 350–382. [Google Scholar]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreases levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammonium chloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhuri, M.A. Implication of hydrogen peroxide-ascorbate system on membrane permeability of water stressed Vigna seedlings. New Phytol. 1985, 99, 355–360. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: II. Purification and quantitative relationship with water-soluble protein in seedlings. Plant Physiol. 1977, 59, 315–318. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, R.A.; Alas, R.M.; Smith, R.J.; Lea, P.J. Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation, in the leaves and roots of wild-type and a catalase-deficient mutant of barley. Physiol. Plant. 1998, 104, 280–292. [Google Scholar] [CrossRef]
- Zhou, W.; Leul, M. Uniconazole-induced tolerance of rape plants to heat stress in relation to changes in hormonal levels, enzyme activities and lipid peroxidation. Plant Growth Regul. 1999, 27, 99–104. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Reinolds, S.S. The use of lead citrate of high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | O3 | Species × O3 | |
|---|---|---|---|
| Shoot biomass | 0.003 | <0.001 | 0.240 |
| Root biomass | 0.077 | 0.001 | 0.742 |
| Total biomass | 0.010 | <0.001 | 0.418 |
| R/S ratio | 0.406 | 0.065 | 0.960 |
| MDA | 0.852 | <0.001 | 0.430 |
| O2−. | <0.001 | <0.001 | 0.145 |
| H2O2 | <0.001 | <0.001 | <0.001 |
| SOD | <0.001 | <0.001 | 0.004 |
| CAT | 0.006 | <0.001 | 0.004 |
| POD | <0.001 | <0.001 | <0.001 |
| APX | 0.081 | <0.001 | 0.473 |
| Chl a | 0.033 | <0.001 | 0.016 |
| Chl b | 0.810 | 0.058 | 0.256 |
| Chl a + b | 0.270 | 0.001 | 0.074 |
| Car | 0.002 | <0.001 | 0.875 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, S.; Li, Y.; Li, B.; He, X.; Chen, W.; Yan, K. Responses of Growth, Oxidative Injury and Chloroplast Ultrastructure in Leaves of Lolium perenne and Festuca arundinacea to Elevated O3 Concentrations. Int. J. Mol. Sci. 2022, 23, 5153. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095153
Xu S, Li Y, Li B, He X, Chen W, Yan K. Responses of Growth, Oxidative Injury and Chloroplast Ultrastructure in Leaves of Lolium perenne and Festuca arundinacea to Elevated O3 Concentrations. International Journal of Molecular Sciences. 2022; 23(9):5153. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095153
Chicago/Turabian StyleXu, Sheng, Yan Li, Bo Li, Xingyuan He, Wei Chen, and Kun Yan. 2022. "Responses of Growth, Oxidative Injury and Chloroplast Ultrastructure in Leaves of Lolium perenne and Festuca arundinacea to Elevated O3 Concentrations" International Journal of Molecular Sciences 23, no. 9: 5153. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095153