
Ebola Virus Encodes Two microRNAs in Huh7-Infected Cells

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
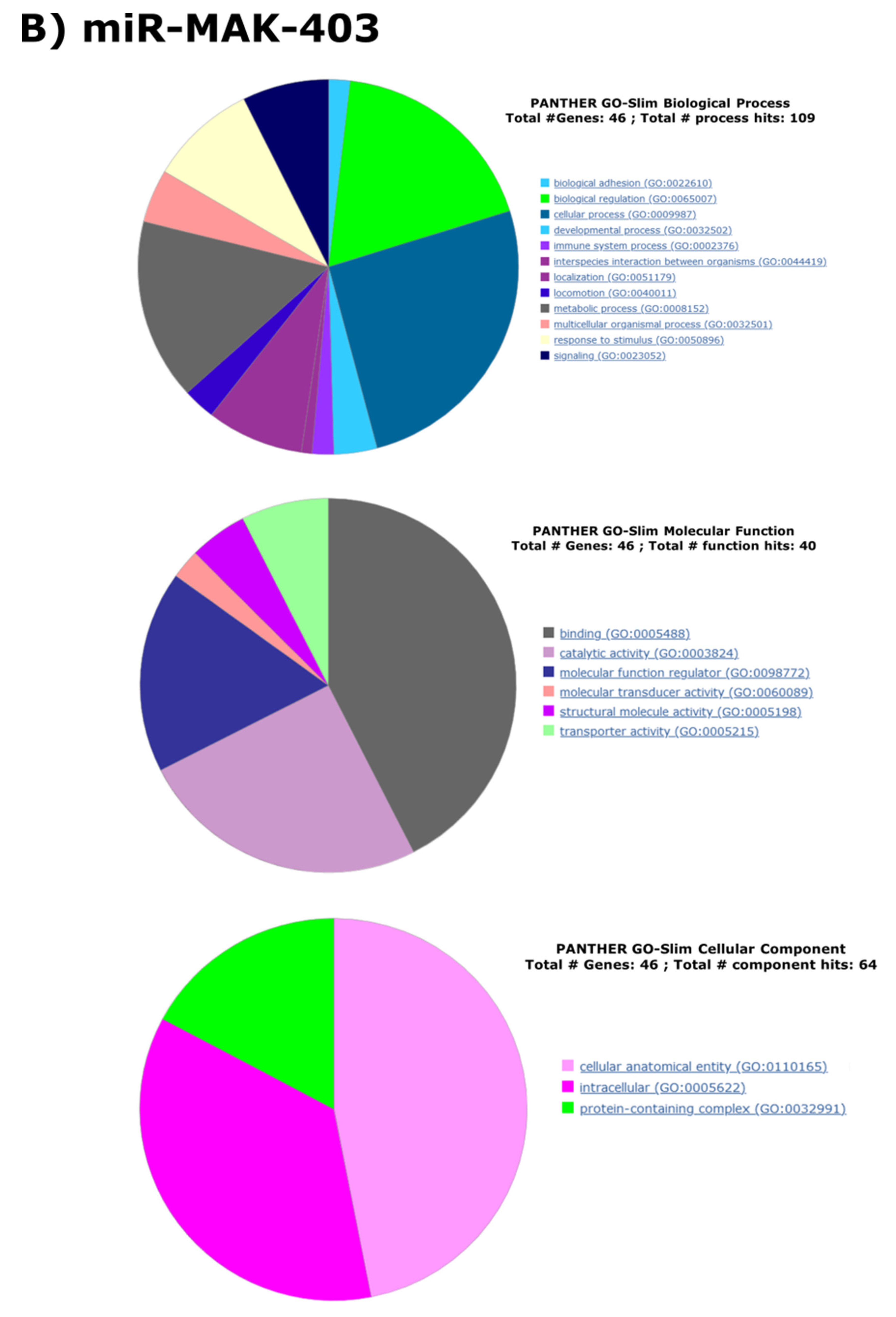{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
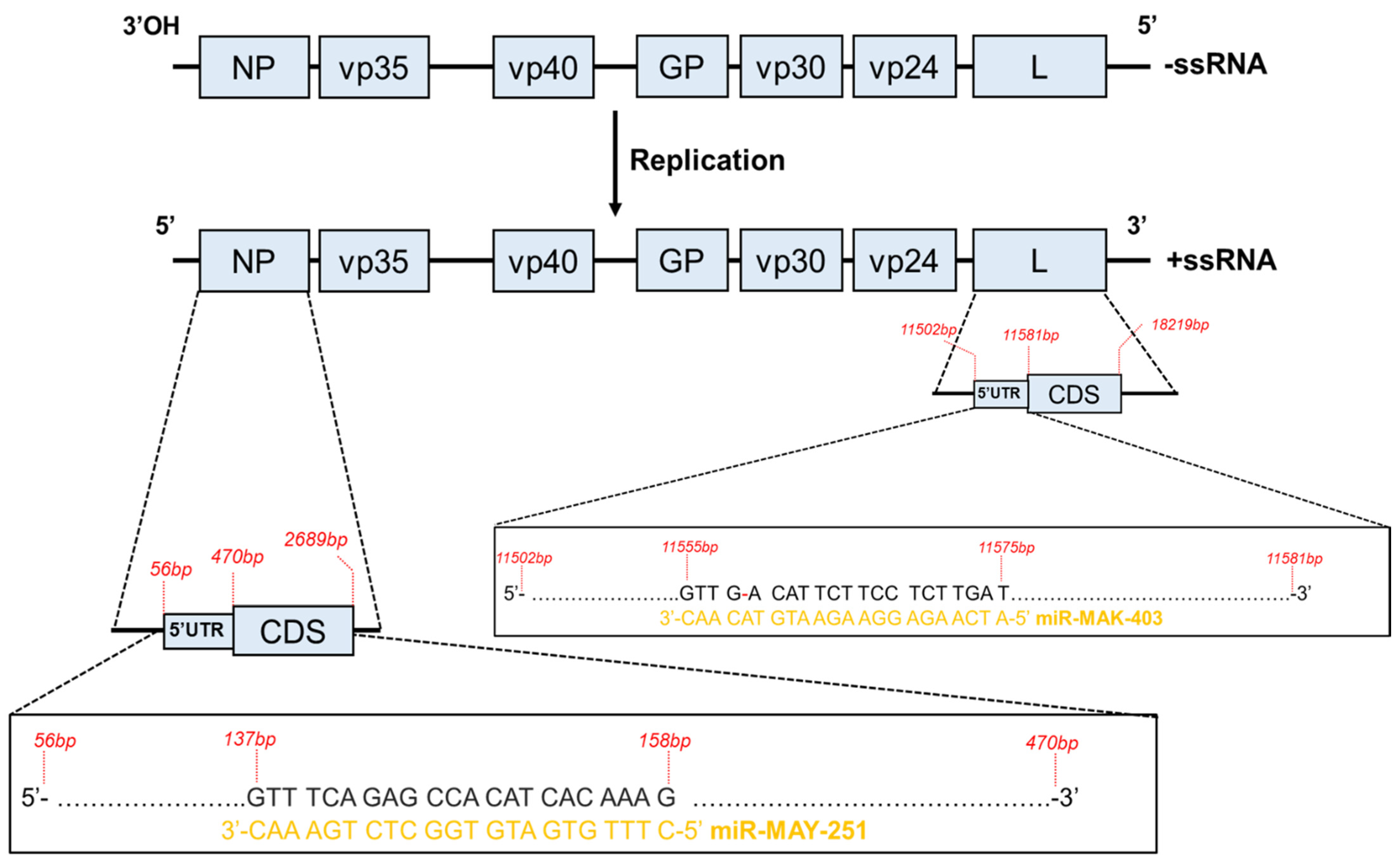
2.1. Description of the Two EBOV-Encoded miRNA Candidates
2.2. miR-MAY-251 and miR-MAK-403 Potential In Silico Human Host Target Candidates
2.3. EBOV miR-MAY-251 May Modulate Selected Host mRNA Targets
2.4. EBOV miR-MAK-403 May Modulate Selected Host mRNA Targets
2.5. miR-MAY-251 and miR-MAK-403 May Directly Regulate DUPS16 and CDK13 through 3′ UTRs, Respectively
3. Discussion
4. Materials and Methods
4.1. Viruses
4.2. Cell Culture Conditions and Transfections
4.3. Plasmid Constructs
4.4. Cell Transfection and Dual Luciferase Assay
4.5. RNA Isolation
4.6. RT-qPCR
4.7. Statical Analysis of qPCR Data
4.8. Illumina Nextseq Sequencing of Cells Infected with EBOV
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ebola Virus Disease. Available online: https://www.who.int/news-room/fact-sheets/detail/ebola-virus-disease (accessed on 1 February 2021).
- Cucinotta, D.; Vanelli, M. WHO Declares COVID-19 a Pandemic. Acta Biomed. 2020, 91, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Muller, J.E.; Nathan, D.G. COVID-19, Nuclear War, and Global Warming: Lessons for Our Vulnerable World. Lancet 2020, 395, 1967–1968. [Google Scholar] [CrossRef]
- Mühlberger, E. Filovirus Replication and Transcription. Future Virol. 2007, 2, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhn, J.H.; Bào, Y.; Bavari, S.; Becker, S.; Bradfute, S.; Brauburger, K.; Brister, J.R.; Bukreyev, A.A.; Caì, Y.; Chandran, K.; et al. Virus Nomenclature below the Species Level: A Standardized Nomenclature for Filovirus Strains and Variants Rescued from CDNA. Arch. Virol. 2014, 159, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Baseler, L.; Chertow, D.S.; Johnson, K.M.; Feldmann, H.; Morens, D.M. The Pathogenesis of Ebola Virus Disease. Annu. Rev. Pathol. Mech. Dis. 2017, 12, 387–418. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Trobaugh, D.W.; Klimstra, W.B. MicroRNA Regulation of RNA Virus Replication and Pathogenesis. Trends Mol. Med. 2017, 23, 80–93. [Google Scholar] [CrossRef]
- Contreras, J.; Rao, D.S. MicroRNAs in Inflammation and Immune Responses. Leukemia 2012, 26, 404–413. [Google Scholar] [CrossRef] [Green Version]
- Duy, J.; Honko, A.N.; Altamura, L.A.; Bixler, S.L.; Wollen-Roberts, S.; Wauquier, N.; O’Hearn, A.; Mucker, E.M.; Johnson, J.C.; Shamblin, J.D.; et al. Virus-Encoded MiRNAs in Ebola Virus Disease. Sci. Rep. 2018, 8, 6480. [Google Scholar] [CrossRef]
- Teng, Y.; Wang, Y.; Zhang, X.; Liu, W.; Fan, H.; Yao, H.; Lin, B.; Zhu, P.; Yuan, W.; Tong, Y.; et al. Systematic Genome-Wide Screening and Prediction of MicroRNAs in EBOV During the 2014 Ebolavirus Outbreak. Sci. Rep. 2015, 5, 9912. [Google Scholar] [CrossRef] [Green Version]
- Grundhoff, A.; Sullivan, C.S. Virus-Encoded MicroRNAs. Virology 2011, 411, 325–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeffer, S.; Zavolan, M.; Grässer, F.A.; Chien, M.; Russo, J.J.; Ju, J.; John, B.; Enright, A.J.; Marks, D.; Sander, C.; et al. Identification of Virus-Encoded MicroRNAs. Science 2004, 304, 734–736. [Google Scholar] [CrossRef] [PubMed]
- Mishra, R.; Kumar, A.; Ingle, H.; Kumar, H. The Interplay Between Viral-Derived MiRNAs and Host Immunity During Infection. Front. Immunol. 2019, 10, 3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boss, I.W.; Renne, R. Viral MiRNAs: Tools for Immune Evasion. Curr. Opin. Microbiol. 2010, 13, 540–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kincaid, R.P.; Burke, J.M.; Sullivan, C.S. RNA Virus MicroRNA That Mimics a B-Cell OncomiR. Proc. Natl. Acad. Sci. USA 2012, 109, 3077–3082. [Google Scholar] [CrossRef] [Green Version]
- Cullen, B.R. Five Questions about Viruses and MicroRNAs. PLoS Pathog. 2010, 6, e1000787. [Google Scholar] [CrossRef]
- Varble, A.; ten Oever, B.R. Implications of RNA Virus-Produced MiRNAs. RNA Biol. 2011, 8, 190–194. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Asgari, S. MicroRNA-like Viral Small RNA from Dengue Virus 2 Autoregulates Its Replication in Mosquito Cells. Proc. Natl. Acad. Sci. USA 2014, 111, 2746–2751. [Google Scholar] [CrossRef] [Green Version]
- Aguado, L.C.; ten Oever, B. RNA Virus Building Blocks-MiRNAs Not Included. PLoS Pathog. 2018, 14, e1006963. [Google Scholar] [CrossRef]
- Shapiro, J.S.; Varble, A.; Pham, A.M.; Tenoever, B.R. Noncanonical Cytoplasmic Processing of Viral MicroRNAs. RNA 2010, 16, 2068–2074. [Google Scholar] [CrossRef] [Green Version]
- Varble, A.; Chua, M.A.; Perez, J.T.; Manicassamy, B.; García-Sastre, A.; ten Oever, B.R. Engineered RNA Viral Synthesis of MicroRNAs. Proc. Natl. Acad. Sci. USA 2010, 107, 11519–11524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omoto, S.; Fujii, Y.R. Regulation of Human Immunodeficiency Virus 1 Transcription by Nef MicroRNA. J. Gen. Virol. 2005, 86, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Bennasser, Y.; Le, S.-Y.; Yeung, M.L.; Jeang, K.-T. HIV-1 Encoded Candidate Micro-RNAs and Their Cellular Targets. Retrovirology 2004, 1, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouellet, D.L.; Plante, I.; Landry, P.; Barat, C.; Janelle, M.-E.; Flamand, L.; Tremblay, M.J.; Provost, P. Identification of Functional MicroRNAs Released through Asymmetrical Processing of HIV-1 TAR Element. Nucleic Acids Res. 2008, 36, 2353–2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klase, Z.; Kale, P.; Winograd, R.; Gupta, M.V.; Heydarian, M.; Berro, R.; McCaffrey, T.; Kashanchi, F. HIV-1 TAR Element Is Processed by Dicer to Yield a Viral Micro-RNA Involved in Chromatin Remodeling of the Viral LTR. BMC Mol. Biol. 2007, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Ouellet, D.L.; Vigneault-Edwards, J.; Létourneau, K.; Gobeil, L.-A.; Plante, I.; Burnett, J.C.; Rossi, J.J.; Provost, P. Regulation of Host Gene Expression by HIV-1 TAR MicroRNAs. Retrovirology 2013, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Aso, S.; Kitao, K.; Hashimoto-Gotoh, A.; Sakaguchi, S.; Miyazawa, T. Identification of Feline Foamy Virus-Derived MicroRNAs. Microbes Environ. 2021, 36, ME21055. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, J.; Zhang, H.; Wang, M.; Gao, G.F.; Li, X. Ebola Virus Encodes a MiR-155 Analog to Regulate Importin-A5 Expression. Cell. Mol. Life Sci. 2016, 73, 3733–3744. [Google Scholar] [CrossRef]
- Liang, H.; Zhou, Z.; Zhang, S.; Zen, K.; Chen, X.; Zhang, C. Identification of Ebola Virus MicroRNAs and Their Putative Pathological Function. Sci. China Life Sci. 2014, 57, 973–981. [Google Scholar] [CrossRef] [Green Version]
- Oliver, G.F.; Orang, A.V.; Appukuttan, B.; Marri, S.; Michael, M.Z.; Marsh, G.A.; Smith, J.R. Expression of MicroRNA in Human Retinal Pigment Epithelial Cells Following Infection with Zaire Ebolavirus. BMC Res Notes 2019, 12, 639. [Google Scholar] [CrossRef]
- Prasad, A.N.; Ronk, A.J.; Widen, S.G.; Wood, T.G.; Basler, C.F.; Bukreyev, A. Ebola Virus Produces Discrete Small Noncoding RNAs Independently of the Host MicroRNA Pathway Which Lack RNA Interference Activity in Bat and Human Cells. J. Virol. 2020, 94, e01441-19. [Google Scholar] [CrossRef] [PubMed]
- Diallo, I.; Ho, J.; Laffont, B.; Laugier, J.; Benmoussa, A.; Lambert, M.; Husseini, Z.; Soule, G.; Kozak, R.; Kobinger, G.P.; et al. Altered MicroRNA Transcriptome in Cultured Human Liver Cells upon Infection with Ebola Virus. Int. J. Mol. Sci. 2021, 22, 3792. [Google Scholar] [CrossRef] [PubMed]
- Wong, N.; Wang, X. MiRDB: An Online Resource for MicroRNA Target Prediction and Functional Annotations. Nucleic Acids Res. 2015, 43, D146–D152. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Lai, J.; Lehman, M.L.; Nelson, C.C. MiRDeep *: An Integrated Application Tool for MiRNA Identification from RNA Sequencing Data. Nucleic Acids Res. 2013, 41, 727–737. [Google Scholar] [CrossRef]
- Friedländer, M.R.; Chen, W.; Adamidi, C.; Maaskola, J.; Einspanier, R.; Knespel, S.; Rajewsky, N. Discovering MicroRNAs from Deep Sequencing Data Using MiRDeep. Nat. Biotechnol. 2008, 26, 407–415. [Google Scholar] [CrossRef]
- Friedländer, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. MiRDeep2 Accurately Identifies Known and Hundreds of Novel MicroRNA Genes in Seven Animal Clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-Scale Gene Function Analysis with the PANTHER Classification System. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef]
- Bakre, A.; Andersen, L.E.; Meliopoulos, V.; Coleman, K.; Yan, X.; Brooks, P.; Crabtree, J.; Tompkins, S.M.; Tripp, R.A. Identification of Host Kinase Genes Required for Influenza Virus Replication and the Regulatory Role of MicroRNAs. PLoS ONE 2013, 8, e66796. [Google Scholar] [CrossRef] [Green Version]
- Berro, R.; Pedati, C.; Kehn-Hall, K.; Wu, W.; Klase, Z.; Even, Y.; Genevière, A.-M.; Ammosova, T.; Nekhai, S.; Kashanchi, F. CDK13, a New Potential Human Immunodeficiency Virus Type 1 Inhibitory Factor Regulating Viral MRNA Splicing. J. Virol. 2008, 82, 7155–7166. [Google Scholar] [CrossRef] [Green Version]
- Xie, M.; Steitz, J.A. Versatile MicroRNA Biogenesis in Animals and Their Viruses. RNA Biol. 2014, 11, 673–681. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.; Wang, Y.; Zhao, Z.; Wu, W.; Su, Y.; Zhang, Z.; Shen, M.; Wu, P.; Qian, H.; Guo, X. Two Putative Cypovirus-Encoded MiRNAs Co-Regulate the Host Gene of GTP-Binding Nuclear Protein Ran and Facilitate Virus Replication. Front. Physiol. 2021, 12, 663482. [Google Scholar] [CrossRef] [PubMed]
- Ouellet, D.L.; Perron, M.P.; Gobeil, L.-A.; Plante, P.; Provost, P. MicroRNAs in Gene Regulation: When the Smallest Governs It All. J. Biomed. Biotechnol. 2006, 2006, 69616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, W.; Legendre, M.; Gautheret, D. RNA Stem-Loops: To Be or Not to Be Cleaved by RNAse III. RNA 2007, 13, 457–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kincaid, R.P.; Sullivan, C.S. Virus-Encoded MicroRNAs: An Overview and a Look to the Future. PLoS Pathog. 2012, 8, e1003018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeshita, A.; Hirabayashi, N.; Ichihara, M.; Miwa, Y. Treatment of Chronic Granulocytic Leukemia in the Accelerated Phase by Transfusion of Autologous Buffy-Coat Cells—A Case Report. Nihon Ketsueki Gakkai Zasshi J. Jpn. Haematol. Soc. 1990, 53, 51–56. [Google Scholar]
- Murphy, E.; Vaníček, J.; Robins, H.; Shenk, T.; Levine, A.J. Suppression of Immediate-Early Viral Gene Expression by Herpesvirus-Coded MicroRNAs: Implications for Latency. Proc. Natl. Acad. Sci. USA 2008, 105, 5453–5458. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-K.; Kim, B.; Kim, V.N. Re-Evaluation of the Roles of DROSHA, Exportin 5, and DICER in MicroRNA Biogenesis. Proc. Natl. Acad. Sci. USA 2016, 113, E1881–E1889. [Google Scholar] [CrossRef] [Green Version]
- Ammosova, T.; Pietzsch, C.A.; Saygideğer, Y.; Ilatovsky, A.; Lin, X.; Ivanov, A.; Kumari, N.; Jerebtsova, M.; Kulkarni, A.; Petukhov, M.; et al. Protein Phosphatase 1–Targeting Small-Molecule C31 Inhibits Ebola Virus Replication. J Infect. Dis. 2018, 218, S627–S635. [Google Scholar] [CrossRef] [Green Version]
- Takamatsu, Y.; Krähling, V.; Kolesnikova, L.; Halwe, S.; Lier, C.; Baumeister, S.; Noda, T.; Biedenkopf, N.; Becker, S. Serine-Arginine Protein Kinase 1 Regulates Ebola Virus Transcription. mBio 2020, 11, e02565-19. [Google Scholar] [CrossRef] [Green Version]
- Batra, J.; Hultquist, J.F.; Liu, D.; Shtanko, O.; Dollen, J.V.; Satkamp, L.; Jang, G.M.; Luthra, P.; Schwarz, T.M.; Small, G.I.; et al. Protein Interaction Mapping Identifies RBBP6 as a Negative Regulator of Ebola Virus Replication. Cell 2018, 175, 1917–1930.e13. [Google Scholar] [CrossRef] [Green Version]
- Stewart, C.M.; Phan, A.; Bo, Y.; LeBlond, N.D.; Smith, T.K.T.; Laroche, G.; Giguère, P.M.; Fullerton, M.D.; Pelchat, M.; Kobasa, D.; et al. Ebola Virus Triggers Receptor Tyrosine Kinase-Dependent Signaling to Promote the Delivery of Viral Particles to Entry-Conducive Intracellular Compartments. PLoS Pathog. 2021, 17, e1009275. [Google Scholar] [CrossRef] [PubMed]
- Schnittler, H.J.; Feldmann, H. Molecular Pathogenesis of Filovirus Infections: Role of Macrophages and Endothelial Cells. Curr. Top. Microbiol. Immunol. 1999, 235, 175–204. [Google Scholar] [CrossRef] [PubMed]
- Wong, G.; Leung, A.; He, S.; Cao, W.; De La Vega, M.-A.; Griffin, B.D.; Soule, G.; Kobinger, G.P.; Kobasa, D.; Qiu, X. The Makona Variant of Ebola Virus Is Highly Lethal to Immunocompromised Mice and Immunocompetent Ferrets. J. Infect. Dis. 2018, 218, S466–S470. [Google Scholar] [CrossRef] [PubMed]
- Keragala, C.B.; Medcalf, R.L. Plasminogen: An Enigmatic Zymogen. Blood 2021, 137, 2881–2889. [Google Scholar] [CrossRef]
- Sidhu, G.; Soff, G.A. The Coagulation System and Angiogenesis. Cancer Treat Res. 2009, 148, 67–80. [Google Scholar] [CrossRef]
- Gadomska, G.; Ziołkowska, K.; Boinska, J.; Filipiak, J.; Rość, D. Activation of TF-Dependent Blood Coagulation Pathway and VEGF-A in Patients with Essential Thrombocythemia. Medicina (Kaunas) 2019, 55, 54. [Google Scholar] [CrossRef] [Green Version]
- Gebhard, C.; Akhmedov, A.; Mocharla, P.; Angstenberger, J.; Sahbai, S.; Camici, G.G.; Lüscher, T.F.; Tanner, F.C. PDGF-CC Induces Tissue Factor Expression: Role of PDGF Receptor Alpha/Beta. Basic Res. Cardiol. 2010, 105, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Wadhwa, M.; Dilger, P.; Tubbs, J.; Mire-Sluis, A.; Barrowcliffe, T.; Thorpe, R. Identification of Transforming Growth Factor-Beta as a Contaminant in Factor VIII Concentrates: A Possible Link with Immunosuppressive Effects in Hemophiliacs. Blood 1994, 84, 2021–2030. [Google Scholar] [CrossRef] [Green Version]
- Gregg, D.; de Carvalho, D.D.; Kovacic, H. Integrins and Coagulation: A Role for ROS/Redox Signaling? Antioxid. Redox Signal. 2004, 6, 757–764. [Google Scholar] [CrossRef]
- Grignani, G.; Maiolo, A. Cytokines and Hemostasis. Haematologica 2000, 85, 967–972. [Google Scholar]
- Li, B.; Clohisey, S.M.; Chia, B.S.; Wang, B.; Cui, A.; Eisenhaure, T.; Schweitzer, L.D.; Hoover, P.; Parkinson, N.J.; Nachshon, A.; et al. Genome-Wide CRISPR Screen Identifies Host Dependency Factors for Influenza A Virus Infection. Nat. Commun. 2020, 11, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, H.-D.; Graber, T.E.; Jia, J.-J.; Vaidya, N.; Gilchrist, V.H.; Xiang, X.; Li, W.; Cowan, K.N.; Gkogkas, C.G.; Jaramillo, M.; et al. Induction of an Alternative MRNA 5’ Leader Enhances Translation of the Ciliopathy Gene Inpp5e and Resistance to Oncolytic Virus Infection. Cell Rep. 2019, 29, 4010–4023.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Tai, A.W. Nir2 Is an Effector of VAPs Necessary for Efficient Hepatitis C Virus Replication and Phosphatidylinositol 4-Phosphate Enrichment at the Viral Replication Organelle. J. Virol. 2019, 93, e00742-19. [Google Scholar] [CrossRef] [PubMed]
- McCune, B.T.; Tang, W.; Lu, J.; Eaglesham, J.B.; Thorne, L.; Mayer, A.E.; Condiff, E.; Nice, T.J.; Goodfellow, I.; Krezel, A.M.; et al. Noroviruses Co-Opt the Function of Host Proteins VAPA and VAPB for Replication via a Phenylalanine-Phenylalanine-Acidic-Tract-Motif Mimic in Nonstructural Viral Protein NS1/2. mBio 2017, 8, e00668-17. [Google Scholar] [CrossRef] [Green Version]
- Amini-Bavil-Olyaee, S.; Choi, Y.J.; Lee, J.H.; Shi, M.; Huang, I.-C.; Farzan, M.; Jung, J.U. The Antiviral Effector IFITM3 Disrupts Intracellular Cholesterol Homeostasis to Block Viral Entry. Cell Host Microbe 2013, 13, 452–464. [Google Scholar] [CrossRef] [Green Version]
- Dantoft, W.; Robertson, K.A.; Watkins, W.J.; Strobl, B.; Ghazal, P. Metabolic Regulators Nampt and Sirt6 Serially Participate in the Macrophage Interferon Antiviral Cascade. Front. Microbiol. 2019, 10, 355. [Google Scholar] [CrossRef] [Green Version]
- Schoggins, J.W.; Wilson, S.J.; Panis, M.; Murphy, M.Y.; Jones, C.T.; Bieniasz, P.; Rice, C.M. A Diverse Range of Gene Products Are Effectors of the Type I Interferon Antiviral Response. Nature 2011, 472, 481–485. [Google Scholar] [CrossRef]
- Heer, C.D.; Sanderson, D.J.; Voth, L.S.; Alhammad, Y.M.O.; Schmidt, M.S.; Trammell, S.A.J.; Perlman, S.; Cohen, M.S.; Fehr, A.R.; Brenner, C. Coronavirus Infection and PARP Expression Dysregulate the NAD Metabolome: An Actionable Component of Innate Immunity. J. Biol. Chem. 2020, 295, 17986–17996. [Google Scholar] [CrossRef]
- Narita, R.; Takahasi, K.; Murakami, E.; Hirano, E.; Yamamoto, S.P.; Yoneyama, M.; Kato, H.; Fujita, T. A Novel Function of Human Pumilio Proteins in Cytoplasmic Sensing of Viral Infection. PLoS Pathog. 2014, 10, e1004417. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Yan, Y.; He, H.; Wang, L.; Zhang, N.; Zhang, J.; Huang, H.; Wu, N.; Ren, H.; Qian, M.; et al. IFN-Stimulated P2Y13 Protects Mice from Viral Infection by Suppressing the CAMP/EPAC1 Signaling Pathway. J. Mol. Cell Biol. 2019, 11, 395–407. [Google Scholar] [CrossRef] [Green Version]
- Kachuri, L.; Francis, S.S.; Morrison, M.L.; Wendt, G.A.; Bossé, Y.; Cavazos, T.B.; Rashkin, S.R.; Ziv, E.; Witte, J.S. The Landscape of Host Genetic Factors Involved in Immune Response to Common Viral Infections. Genome Med. 2020, 12, 93. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Li, R.; Meng, J.-L.; Mao, H.-T.; Zhang, Y.; Zhang, J. Smurf2 Negatively Modulates RIG-I–Dependent Antiviral Response by Targeting VISA/MAVS for Ubiquitination and Degradation. J. Immunol. 2014, 192, 4758–4764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verga-Gérard, A.; Porcherot, M.; Meyniel-Schicklin, L.; André, P.; Lotteau, V.; Perrin-Cocon, L. Hepatitis C Virus/Human Interactome Identifies SMURF2 and the Viral Protease as Critical Elements for the Control of TGF-β Signaling. FASEB J. 2013, 27, 4027–4040. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Barouch-Bentov, R.; Xiao, F.; Schor, S.; Pu, S.; Biquand, E.; Lu, A.; Lindenbach, B.D.; Jacob, Y.; Demeret, C.; et al. MARCH8 Ubiquitinates the Hepatitis C Virus Nonstructural 2 Protein and Mediates Viral Envelopment. Cell Rep. 2019, 26, 1800–1814.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, S.; Diep, J.; Feng, N.; Ren, L.; Li, B.; Ooi, Y.S.; Wang, X.; Brulois, K.F.; Yasukawa, L.L.; Li, X.; et al. STAG2 Deficiency Induces Interferon Responses via CGAS-STING Pathway and Restricts Virus Infection. Nat. Commun. 2018, 9, 1485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.-M.; Zhou, J.; Xu, Z.; Huang, H.-J.; Chen, M.-J.; Ji, J.-S. MicroRNA-27a-3p Inhibits Cell Viability and Migration through down-Regulating DUSP16 in Hepatocellular Carcinoma. J. Cell Biochem. 2018, 119, 5143–5152. [Google Scholar] [CrossRef] [PubMed]
- Niedzielska, M.; Bodendorfer, B.; Münch, S.; Eichner, A.; Derigs, M.; da Costa, O.; Schweizer, A.; Neff, F.; Nitschke, L.; Sparwasser, T.; et al. Gene Trap Mice Reveal an Essential Function of Dual Specificity Phosphatase Dusp16/MKP-7 in Perinatal Survival and Regulation of Toll-like Receptor (TLR)-Induced Cytokine Production. J. Biol. Chem. 2014, 289, 2112–2126. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Guo, Z.; Hu, Z.; Liu, M.; Chen, Y.; Chen, S.; Peng, B.; Zhang, P.; Wu, Z.; Luo, H.; et al. FGD1 Exhibits Oncogenic Properties in Hepatocellular Carcinoma through Regulating Cell Morphology, Autophagy and Mitochondrial Function. Biomed. Pharm. 2020, 125, 110029. [Google Scholar] [CrossRef]
- Wang, H.; Wu, Q.; Li, S.; Zhang, B.; Chi, Z.; Hao, L. Unc5D Regulates P53-Dependent Apoptosis in Neuroblastoma Cells. Mol. Med. Rep. 2014, 9, 2411–2416. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Huang, K.; Zeng, X.; Liu, Z.; Liao, X.; Yang, C.; Yu, T.; Han, C.; Zhu, G.; Qin, W.; et al. Diagnostic and Prognostic Value of MRNA Expression of Phospholipase C β Family Genes in Hepatitis B Virus-associated Hepatocellular Carcinoma. Oncol. Rep. 2019, 41, 2855–2875. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.K. MiRNAs Target Databases: Developmental Methods and Target Identification Techniques with Functional Annotations. Cell Mol. Life Sci. 2017, 74, 2239–2261. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Buchan, R.J.; Cook, S.A. MicroRNA-223 Regulates Glut4 Expression and Cardiomyocyte Glucose Metabolism. Cardiovasc. Res. 2010, 86, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S. Posttranscriptional Upregulation by MicroRNAs. Wiley Interdiscip. Rev. RNA 2012, 3, 311–330. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.-W.; Wang, F.; Wei, Q.; Zhao, Y.-F.; Liu, M.; Li, X.; Tang, H. MiR-20a Promotes Migration and Invasion by Regulating TNKS2 in Human Cervical Cancer Cells. FEBS Lett. 2012, 586, 897–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, F.; Liu, X.; Li, D.; Wang, P.; Li, N.; Lu, L.; Cao, X. MicroRNA-466l Upregulates IL-10 Expression in TLR-Triggered Macrophages by Antagonizing RNA-Binding Protein Tristetraprolin-Mediated IL-10 MRNA Degradation. J. Immunol. 2010, 184, 6053–6059. [Google Scholar] [CrossRef]
- Zhang, M.-M.; Sun, F.; Cui, B.; Zhang, L.-L.; Fang, Y.; Li, Y.; Zhang, R.-J.; Ye, X.-P.; Ma, Y.-R.; Han, B.; et al. Tumor-Suppressive Function of UNC5D in Papillary Thyroid Cancer. Oncotarget 2017, 8, 96126–96138. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, B.; Gu, M.; Li, S.; Chi, Z.; Hao, L. Overexpression of the Dependence Receptor UNC5H4 Inhibits Cell Migration and Invasion, and Triggers Apoptosis in Neuroblastoma Cell. Tumour Biol. 2014, 35, 5417–5425. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, Y.; Haraguchi, S.; Yu, M.; Ohira, M.; Ozaki, T.; Nakagawa, A.; Ushijima, T.; Isogai, E.; Koseki, H.; et al. Dependence Receptor UNC5D Mediates Nerve Growth Factor Depletion-Induced Neuroblastoma Regression. J. Clin. Investig. 2013, 123, 2935–2947. [Google Scholar] [CrossRef] [Green Version]
- Plissonnier, M.-L.; Lahlali, T.; Michelet, M.; Lebossé, F.; Cottarel, J.; Beer, M.; Neveu, G.; Durantel, D.; Bartosch, B.; Accardi, R.; et al. Epidermal Growth Factor Receptor-Dependent Mutual Amplification between Netrin-1 and the Hepatitis C Virus. PLoS Biol. 2016, 14, e1002421. [Google Scholar] [CrossRef]
- Lara-Pezzi, E.; Pezzi, N.; Prieto, I.; Barthelemy, I.; Carreiro, C.; Martínez, A.; Maldonado-Rodríguez, A.; López-Cabrera, M.; Barbero, J.L. Evidence of a Transcriptional Co-Activator Function of Cohesin STAG/SA/Scc3. J. Biol. Chem. 2004, 279, 6553–6559. [Google Scholar] [CrossRef] [Green Version]
- Pleet, M.L.; DeMarino, C.; Lepene, B.; Aman, M.J.; Kashanchi, F. The Role of Exosomal VP40 in Ebola Virus Disease. DNA Cell Biol. 2017, 36, 243–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepley-McTaggart, A.; Schwoerer, M.P.; Sagum, C.A.; Bedford, M.T.; Jaladanki, C.K.; Fan, H.; Cassel, J.; Harty, R.N. Ubiquitin Ligase SMURF2 Interacts with Filovirus VP40 and Promotes Egress of VP40 VLPs. Viruses 2021, 13, 288. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Chen, S.-P.; Zhang, X.-P.; Wang, H.; Zhu, C.; Lin, H.-Y. Smurf2 Participates in Human Trophoblast Cell Invasion by Inhibiting TGF-Beta Type I Receptor. J. Histochem. Cytochem. 2009, 57, 605–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C. Roles of E3 Ubiquitin Ligases in Cell Adhesion and Migration. Cell Adh. Migr. 2010, 4, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brenner, C.; Galluzzi, L.; Kepp, O.; Kroemer, G. Decoding Cell Death Signals in Liver Inflammation. J. Hepatol. 2013, 59, 583–594. [Google Scholar] [CrossRef] [Green Version]
- Longo, P.A.; Kavran, J.M.; Kim, M.-S.; Leahy, D.J. Transient Mammalian Cell Transfection with Polyethylenimine (PEI). Methods Enzymol. 2013, 529, 227–240. [Google Scholar] [CrossRef] [Green Version]
- Dalby, B.; Cates, S.; Harris, A.; Ohki, E.C.; Tilkins, M.L.; Price, P.J.; Ciccarone, V.C. Advanced Transfection with Lipofectamine 2000 Reagent: Primary Neurons, SiRNA, and High-Throughput Applications. Methods 2004, 33, 95–103. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A Tool to Design Target-Specific Primers for Polymerase Chain Reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Vester, B.; Wengel, J. LNA (Locked Nucleic Acid): High-Affinity Targeting of Complementary RNA and DNA. Biochemistry 2004, 43, 13233–13241. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diallo, I.; Husseini, Z.; Guellal, S.; Vion, E.; Ho, J.; Kozak, R.A.; Kobinger, G.P.; Provost, P. Ebola Virus Encodes Two microRNAs in Huh7-Infected Cells. Int. J. Mol. Sci. 2022, 23, 5228. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095228
Diallo I, Husseini Z, Guellal S, Vion E, Ho J, Kozak RA, Kobinger GP, Provost P. Ebola Virus Encodes Two microRNAs in Huh7-Infected Cells. International Journal of Molecular Sciences. 2022; 23(9):5228. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095228
Chicago/Turabian StyleDiallo, Idrissa, Zeinab Husseini, Sara Guellal, Elodie Vion, Jeffrey Ho, Robert A. Kozak, Gary P. Kobinger, and Patrick Provost. 2022. "Ebola Virus Encodes Two microRNAs in Huh7-Infected Cells" International Journal of Molecular Sciences 23, no. 9: 5228. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095228