
Recent Progress on the Localization of PLK1 to the Kinetochore and Its Role in Mitosis


Department of Biology Education, Pusan National University, 2, Busandaehak-ro 63beon-gil, Geumjeong-gu, Busan 46241, Korea
Int. J. Mol. Sci. 2022, 23(9), 5252; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095252
Submission received: 29 March 2022
/
Revised: 3 May 2022
/
Accepted: 4 May 2022
/
Published: 8 May 2022
(This article belongs to the Special Issue Molecular Control of Chromosome Segregation and Stability in Mitosis)
{kind=link}
{kind=link}
{kind=link}
Abstract
:The accurate distribution of the replicated genome during cell division is essential for cell survival and healthy organismal development. Errors in this process have catastrophic consequences, such as birth defects and aneuploidy, a hallmark of cancer cells. PLK1 is one of the master kinases in mitosis and has multiple functions, including mitotic entry, chromosome segregation, spindle assembly checkpoint, and cytokinesis. To dissect the role of PLK1 in mitosis, it is important to understand how PLK1 localizes in the specific region in cells. PLK1 localizes at the kinetochore and is essential in spindle assembly checkpoint and chromosome segregation. However, how PLK1 localizes at the kinetochore remains elusive. Here, we review the recent literature on the kinetochore recruitment mechanisms of PLK1 and its roles in spindle assembly checkpoint and attachment between kinetochores and spindle microtubules. Together, this review provides an overview of how the local distribution of PLK1 could regulate major pathways in mitosis.
Keywords:
PLK1 kinase; kinetochore; spindle assembly checkpoint; chromosome segregation; PP1; PP2A; cell cycle1. Introduction
During mitosis, it is critical for cells to correctly segregate duplicated genomes into daughter cells. Any errors arising in this process can cause aneuploidy observed in most cancer cells and may lead to birth defects [1,2]. Therefore, it is important to understand how chromosomes are accurately segregated during cell division.
A major component of the machinery required for this process is the kinetochore, a large multiprotein complex that mediates the attachment of chromosomes to spindle microtubule ends. In addition, kinetochores serve as a signaling platform for monitoring errors in attachment and directing progression through the cell cycle. When kinetochores are not attached to spindle microtubules, they delay the progression of anaphase by activating the spindle assembly checkpoint (SAC) to provide sufficient time for the interaction between kinetochores and microtubules. The SAC inhibits activation of the anaphase-promoting complex/cyclosome (APC/C), a large E3 ubiquitin ligase required for sister chromatid separation and execution of the mitotic exit [3,4].
An essential group of mitotic kinases, including CDK1, PLK1, Aurora B, Mps1, and Bub1, and the counteracting phosphatases PP1 and PP2A, are thought to regulate kinetochore assembly and chromosome segregation [5,6]. Polo-like kinase 1 (PLK1), a well-conserved mitotic serine/threonine kinase, is an essential mitotic kinase that localizes to kinetochores, centrosomes, and the central spindle and midbody. It plays important roles in mitotic progression, centrosome assembly, chromosome segregation, spindle elongation, and cytokinesis [7,8,9].
In addition, PLK1 has also been shown to be overexpressed in various types of cancers, including breast cancer, rectal cancer, colorectal cancer, pancreatic cancer, ovarian cancer, and lung cancer [10,11,12]. Overexpression of PLK1 in cancer cells is also associated with poor prognoses [10,11,13,14,15,16]. In fact, many different PLK1 inhibitors have been developed, and more than 10 PLK1 inhibitors are commercially available. Furthermore, some of the PLK1 inhibitors are in clinical trials and have demonstrated promising results in the treatment of many types of cancer [10,13].
However, it is currently unclear why overexpression of PLK1 causes cancer or is a consequence of the overproliferation of cancer cells [17]. If PLK1 is an oncogene, identifying which pathways involve PLK1 and cause carcinogenesis will be critical in developing therapeutic strategies. For example, chromosomal instability is a common feature in cancer cells and can be caused by chromosome missegregation in mitosis [1]. Therefore, elucidating how PLK1 regulates mitosis is an important question to be answered.
Because PLK1 has many diverse functions in mitosis, it is crucial to understand how PLK1 localizes at specific regions during mitosis, which will allow the dissection of the role of PLK1. PLK1 plays a critical role in SAC and chromosome alignment, and many candidates were suggested to recruit PLK1 to the kinetochore. However, the component essential for kinetochore recruitment of PLK1 remains unclear. Here, we first describe the structure and activation mechanism of PLK1 and then elaborate on how PLK1 localizes at the kinetochore. Finally, we discuss the role of PLK1 in SAC, chromosome alignment, and microtubule attachment.
2. Molecular Properties and the Activation of PLK1
PLK1 was initially discovered via genetic screening in budding yeast, in which the temperature mutation failed to progress through the cell cycle. The name Polo-like kinase (Plk) originated from a discovery in Drosophila, where the mutation of the gene resulted in abnormal spindle poles. PLK1 is well conserved among species, and almost all eukaryotes, except plants, have at least one form of the PLK family [18,19,20,21]. While flies, budding yeast, and fission yeast only have a single PLK family member, humans, mice, frogs, and worms have multiple PLK family members [22,23]. In humans, there are five PLKs, from PLK1 to PLK5, and they have little overlap in their localization and function, despite some structural similarity [8,22]. PLK2 is required for centrosome maturation and centriole duplication in the S phase, and PLK3 promotes DNA replication during the G1/S transition and is involved in the stress response pathways [24,25,26,27,28,29,30,31]. PLK4 plays a role in centriole duplication in the S phase, and PLK5 functions in neuron differentiation [31,32,33,34]. PLK1 is mainly expressed in late G2 and M phases and plays critical roles in the regulation of mitosis and cytokinesis [22,31,35,36,37].
PLKs are defined by the presence of an N-terminal catalytic Ser/Thr kinase domain and a C-terminal polo-box domain (PBD) that phosphorylate target proteins and control PLK1 kinase activity and localization, respectively [7,8,9,22]. In humans, kinase domains are highly conserved among all PLKs, except for PLK5, which lacks the kinase domain [22,23,33,38]. In contrast, the PBD region, which consists of two polo-box (PB) motifs, intervening residues, and a part of the linker between the kinase domain and PB1, is less well conserved [23] (Figure 1).
Among all PLK family members, PLK1 is the most highly conserved. PLK1 has also been the most extensively studied because of its importance in cell division. The kinase domain of PLK1 phosphorylates Ser or Thr of substrates that contain the consensus motif [D/N/E/Y]-[X]-S/T-[F/Φ; no P]-[Φ/X] [9,23]. Moreover, the kinase domain contains a T-loop, also called an activation loop, which allows PLK1 to convert ATP to ADP, transferring the phosphate group to PLK downstream phosphorylation substrates [39]. The PBD is required for localization at the target region, such as the centrosome, centromere, and the kinetochore at prometaphase and metaphase, and the central spindle and midbody at anaphase [40,41,42,43]. The expression of the kinase domain alone is not sufficient for PLK1 localization at the centromere and the kinetochore, and the expression of the PBD alone localizes strongly at the centromere and weakly at the kinetochore [44]. PBD recognizes the S-[pS/pT]-[P/X] motif of the target protein, and when X is proline, proline-directed kinases, including the Cdk1 kinase, can phosphorylate Ser or Thr [45,46]. Phosphorylation of the preceding Ser or Thr primes the motif, thereby generating a binding site for PBD docking [9,42]. Alternatively, when the S-[pS/pT]-[P/X] motif is phosphorylated by PLK1 itself, known as self-priming, PBD can interact with PLK1 [42].
Furthermore, PBD is required to control kinase activity. PBD docking partially activates Plk1 by inducing conformational changes [47]. However, to fully activate PLK1 and relieve the intramolecular interaction between the kinase domain and PBD, which stabilizes the inactive conformation of PLK1, phosphorylation of the T-loop in the kinase domain is required [40,47,48,49,50]. The T-loop of the kinase domain can be phosphorylated by Bora, and Aurora A and B, which localize at the centrosomes and kinetochore, respectively [49,50,51,52,53].
3. Function of PLK1 in Mitosis
PLK1 is a master mitotic kinase with diverse and important functions in mitosis, including mitotic entry, a G2/M checkpoint, centrosome assembly, chromosome segregation, a spindle assembly checkpoint, and cytokinesis [54,55,56,57,58,59,60,61,62,63,64,65,66,67]. PLK1 expression levels start to increase at the G2 phase and peak in the metaphase of the M phase. Upon completion of mitosis, PLK1 levels sharply decrease [35,36,37].
PLK1 is essential for mitotic entry and exit. Mitotic entry is controlled by CDK/cyclin B complex activity, which is regulated by phosphorylation. PLK1 activates Cdc25 phosphatase, which in turn inactivates the Wee1 and dephosphorylates CDK/cyclin B, leading to the activation of CDK/cyclin B [68,69,70,71,72]. In addition, PLK1 promotes Wee1 degradation by phosphorylation, which decreases phosphorylation of CDK/cyclin B by Wee1 kinase [73]. PLK1 is also involved in the activation of transcription factor Forkhead Box M1 (FoxM1), which allows the expression of the genes involved in the G2/M transition [74]. Additionally, PLK1 promotes activation of the anaphase-promoting complex (APC/C) and the degradation of the APC/C inhibitor Emi1 to exit mitosis and enter cytokinesis [75,76,77,78,79].
PLK1 also contributes to centriole duplication and centrosome maturation. Other PLK family members, PLK2 and PLK4, also localize to the centrosomes, but the function of PLK1 in centrosome maturation is not redundant with that of PLK2 or PLK4. During the S and M phases, PLK1 promotes centrosome maturation by phosphorylating several centriole proteins and nucleation factors, including pericentrin, cnn, Nlp, Kizuna, and γ-tubulin [80,81,82,83,84]. In addition, PLK1 allows centriole disengagement, leading to centriole duplication [85]. Interestingly, PLK1 was also suggested to contribute to DNA replication during the S phase [86].
During M phases, PLK1 localizes to the kinetochore and centrosomes and controls mitotic progression by regulating the spindle assembly checkpoint and attachment of kinetochores to spindle microtubules, which is discussed below. Moreover, PLK1 is involved in regulating other mitosis kinases. PLK1 increases the expression of Aurora B by phosphorylating FoxM1 [74]. PLK1 also activates Haspin by phosphorylation [87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103].
When cells enter anaphase, PLK1 dissipates from the kinetochore, moves to the central spindle and spindle midbody, and plays a role in the initiation of cytokinesis [60,61,63,66,67]. Inhibition of PLK1 abolishes RhoA accumulation at the equatorial cortex and disrupts the localization of Ect2, the GEF protein essential for Rho activation, to the spindle midzone [61]. This suggests that PLK1 is critical for the Ect2-RhoA network, which allows proper cleavage furrow and cytokinetic bridge formation, leading to the initiation of cytokinesis.
4. Kinetochore Recruitment Mechanism of PLK1
Because of the importance of PLK1 during mitosis, the molecular mechanism of the kinetochore localization of PLK1 has been extensively studied. PLK1 has been suggested to be recruited to the kinetochore by interacting with many proteins, including the kinetochore components Bub1, BubR1, CENP-U (also called PBIP1), and other proteins, including nuclear distribution protein C (NUDC), CLIP-170, dynactin subunit p27, INCENP, CLASP2, Survivin, SGO1, NCAPG2, USP16, and RSF1 [87,88,90,93,94,95,97,98,99,100,101,102,103,104,105,106,107,108]. Localization of PLK1 is mediated by the interaction of its PBD with proteins that contain its S-[pT/pS]-P/X consensus-binding motif, which is primed by the master mitotic kinase Cdk1 [45,46]. Accordingly, all the listed proteins, except SGO1, contain the S-[pT/pS]-P/X motif, and inhibition of these proteins or the introduction of mutations in the S-[pT/pS]-P/X motif results in reduced kinetochore recruitment of PLK1 [87,90,93,94,95,96,97,100,101,102,103,104,106,108] (Figure 2). However, it is unclear whether these interacting partners equally contribute to kinetochore recruitment of PLK1 or whether the initial interaction of PLK1 with only one or few proteins facilitates the interaction of PLK1 with other PLK1-binding proteins.
Additionally, Aurora B may contribute to PLK1 kinetochore localization. Aurora B contributes to the kinetochore localization of Bub1, a kinetochore component that recruits PLK1 to the kinetochore [109]. Moreover, the T-loop in the kinase domain of PLK1 can be phosphorylated by Aurora A or B, resulting in the activation and kinetochore recruitment of PLK1 [50,51]. Furthermore, because Aurora B and phosphatase PP2A-B56 counteract each other during mitosis, Aurora B indirectly inhibits dephosphorylation by PP2A-B56, which in turn contributes to the recruitment of PLK1 at the kinetochore [5,110].
Although several proteins were suggested to recruit PLK1 to the kinetochore, three groups recently suggested that CENP-U and Bub1 are the main PLK1 kinetochore receptors during mitosis [90,104,111]. Nguyen et al. and Chen et al. suggested that Bub1 and CENP-U act in parallel pathways to recruit PLK1 to the kinetochore using cell biological approaches. They both showed that depletion of the CENP-O complex, which includes CENP-U or CENP-U, resulted in a significant decrease in kinetochore localization of PLK1. In addition, depletion of only the CENP-U or CENP-O complex compromised chromosome alignment marginally; however, when it was combined with Bub1 depletion, the PLK1 level at the kinetochore was further decreased, and the chromosome alignment defect was significantly increased. These results indicate that both CENP-U and Bub1 work together to reach a certain threshold level for PLK1 at the kinetochore for normal function. Interestingly, Nguyen et al. showed that the requirement of the CENP-O complex for kinetochore localization of PLK1 was variable between cell lines, which suggests that different expression levels of Bub1 and CENP-U could affect the requirement of these components for normal kinetochore recruitment of PLK1.
Singh et al. first tested the role of Aurora B in recruiting PLK1 to the kinetochore. They showed that the contribution of Aurora B to the PLK1 kinetochore is indirect. Inhibition of Aurora B with hesperadin caused a strong decrease in the PLK1 level at the kinetochore. However, this was rescued with phosphatase inhibitor okadaic acid (OA), suggesting that Aurora B indirectly contributes to the PLK1 level at the kinetochore by counteracting the phosphatases PP2A-B56 and PP1. In contrast, Bub1 directly interacts with PLK1-PBD but only in the presence of priming kinases PLK1 and CDK1 in vitro. Although inhibition of kinetochore recruitment of Bub1 by treating human cells with an Mps1 kinase inhibitor significantly decreased the levels of PLK1 at the kinetochore, it left residual levels of PLK1 at the kinetochore, indicating that additional PLK1 receptors were required. Through in vitro reconstitution of the 22 kinetochore components, Singh et al. found that among the inner kinetochore components that comprise constitutive centromere-associated network (CCAN), CENP-U is the primary receptor of PLK1. In addition, ectopic localization of CENP-U increased the PLK1 level at the ectopic site, suggesting that CENP-U is necessary for the kinetochore localization of PLK1. Furthermore, the co-depletion of Bub1 and CENP-OPQUR complex completely abolished the kinetochore recruitment of PLK1, and the PLK1 level was not rescued by OA treatment, indicating that Bub1 and CENP-U are the main receptors for PLK1 at the kinetochore.
As suggested for other PLK1 receptors, interactions between CENP-U, Bub1, and PLK1 are heavily dependent on priming by CDK1-mediated phosphorylation. Although these results indicate that Bub1 and CENP-U are essential for kinetochore recruitment of PLK1, they do not exclude the possibility that other known PLK1 receptors are important in the normal cell progression because all experiments with human cells are conducted using the nocodazole treatment, which removes the spindle microtubules during mitosis.
Once all kinetochores attach to the spindle microtubules at metaphase, PLK1 is ubiquitinated by active APC/C, resulting in its dissociation. Ubiquitin-specific peptidase 16 (Usp16) antagonizes the activity of CUL-3-based ubiquitin ligase by deubiquitinating PLK1 at the kinetochore [94]. Furthermore, Usp16 promotes chromosome alignment by regulating kinetochore localization of PLK1 through phosphorylation of the Cdk1 site of USP16 [94].
In the future, it will be valuable to investigate whether the initial recruitment of PLK1 by CENP-U and Bub1 activates phosphorylation of downstream targets that promote further recruitment of PLK1, thereby clarifying how the PLK1-binding proteins at the kinetochore regulate PLK1 dynamics and function.
5. PLK1 Function in Spindle Checkpoint Activation and Spindle Checkpoint Silencing
Although the kinetochore localization of PLK1 increases at metaphase, how PLK1 functions within the kinetochore is unclear because of the complexity of the interdependence of many kinases and phosphatases and because many PLK1 receptors are distributed along the kinetochore and centromere.
One of the major functions of PLK1 during mitosis is the activation of the spindle assembly checkpoint (SAC). The SAC is a safeguard system that halts anaphase progression by inhibiting APC/C until all kinetochores attach to the spindle microtubules. APC/C inhibition is caused by the accumulation of SAC components at unattached kinetochores, a process that mediates the formation of the mitotic checkpoint complex (MCC) composed of Mad2, Bub3, BubR1, and Cdc20 [3,112].
The accumulation of SAC components involves phosphorylation of SAC components by Mps1 kinase [113,114,115,116,117,118,119,120,121,122]. Both PLK1 and Mps1 share a similar phosphorylation consensus motif D/E-x-S/T-Φ, with a strong preference for D/E residues at position-2 [123,124,125,126,127]. Consequently, PLK1 and Mps1 may share many SAC substrates. Caenorhabditis elegans lacks the Mps1 kinase; thus, PLK1 potentially phosphorylates several known Mps1 substrates, which is discussed later [128]. However, it remains unclear whether PLK1 can completely substitute for Mps1 kinase in other animal systems.
The Bub1/Bub3 complex activates SAC by recruiting Mad1, Mad2, BubR1, and Cdc20 to unattached kinetochores. Kinetochore recruitment of Bub/Bub3 is mediated by the core outer kinetochore component KNL1 via multiple “MELT (Met-Glu-Leu-Thr)” motifs, which are phosphorylated by Mps1 and PLK1 [113,114,115] Additionally, Mps1 phosphorylates the CM1 motif of Bub1, and its phosphorylation is required for its interaction with Mad1 [129,130]. Alternatively, PLK1 is able to phosphorylate the Bub1-CM1 motif in C. elegans [89]. Mps1 is also suggested to phosphorylate MAD-1 C-terminus to promote interaction with the N-terminal region of CDC20 [129,131]. In vitro data using C. elegans proteins showed that phosphorylation of PLK-1 at the MAD-1 C-terminus promoted its interaction with the CDC-20 N-terminus [89].
PLK1 also contributes to SAC activation by phosphorylating the APC/C co-activator Cdc20. PLK1 that interacts with S-[pS/pT]-[P/X] motif of Bub1 at the kinetochore suggested to phosphorylate Cdc20, resulting in the inhibition of the interaction between Cdc20 and APC/C [132].
Collectively, these results suggest that PLK1 may activate SAC by phosphorylating Knl1, Bub1, Mad1, and Cdc20. When kinetochores are unattached, phosphorylation-mediated interaction of Cdc20 with Bub1 and Mad1 can occur at the kinetochores, which is critical for the formation of Mad2-Cdc20, which mediates the formation of MCC (Figure 3). However, which pool of PLK1 phosphorylates these kinetochore components in vivo requires further investigation. It will be enlightening to knock down specific PLK1 pools that interact with Bub1 or CENP-U, or other known PLK1 receptors, to determine the effects of the phosphorylation of one of those PLK1 substrates and the SAC activation.
In addition to substituting for Mps1 activity, PLK1 activates Mps1 by phosphorylating multiple sites of Mps1, although whether such phosphorylation directly activates Mps1 is unclear [134].
Once all kinetochores are attached to spindle microtubules, all SAC components are removed from the kinetochore, which extinguishes the SAC signal and anaphase progression by activating APC/C [3]. For SAC silencing, PP2A-B56 and PP1 are the major phosphatases that dephosphorylate the MELT motifs of KNL1, thereby removing Bub1 from the kinetochore [135,136,137,138,139,140,141,142,143]. Although PLK1 may activate SAC, paradoxically, PLK1 may also help silence SAC by recruiting PP2A to the kinetochore. BubR1 contains a KARD motif that interacts with PP2A-B56, and phosphorylation of the KARD motif by PLK1 enhances the interaction with PP2A-B56 [99,144,145]. However, PP2A-B56 and PP1 may silence the spindle checkpoint by dephosphorylating the polo-binding motif of Bub1 and BubR1, which removes PLK1 from the kinetochore [146]. To clarify the function of PLK1 in the SAC, the functions of phosphatases PP1 and PP2A-B56 in SAC silencing must be studied. Whether the removal of PLK1 from the kinetochore is the major cause of silencing of the spindle checkpoint requires further study.
6. A Role of PLK1 in Stabilizing Kinetochore-Microtubule Attachment
In addition to the function of PLK1 in SAC activation, PLK1 plays a role in stabilizing the kinetochore-microtubule attachment during prometaphase [62]. This is somewhat contradictory because SAC activation delays the onset of anaphase, and stabilizing kinetochore-microtubule attachment promotes the onset of anaphase. Mps1, which can be substituted by PLK1 in some cases, activates the SAC and inhibits the kinetochore-microtubule attachment. Inhibition of PLK1 can destabilize or stabilize kinetochore-microtubule attachment under different experimental conditions [55,77,110]. Potentially, PLK1 initially helps form the kinetochore-microtubule attachment by activating the SAC, and once the kinetochore interacts with the microtubule, PLK1 helps stabilize the kinetochore-microtubule attachment. The dual function of the PLK1 may help cells respond quickly and efficiently to the change in the kinetochore-microtubule dynamics. However, whether or how PLK1 activity is controlled within such a short period needs to be further studied.
Among all suggested PLK1 receptors that localize at the kinetochore or centromere, some may contribute to stabilizing kinetochore-microtubule attachment. CENP-U, BubR1, PICH, CLIP-170, CLASP2, Survivin, USP16, and RSF1, which interact with PBD and are phosphorylated by PLK1, showed defects in chromosome alignment when a phospho-mutant was introduced [62,90,92,94,96,97,99,100,105,106,136,144,145,147]. However, it is unclear whether the defects in the phospho-mutants of PLK1 receptors are caused by reduced localization of PLK1 or other functions of PLK1 receptors. The localization of PLK1 in a specific region by PLK1 receptors may promote phosphorylation of other proteins at close distances that participate in kinetochore-microtubule attachment. For instance, the kinetochore-localized pool of PLK1 binds to CENP-U phosphorylates CENP-Q at multiple sites; mutations of these phosphorylation sites can cause defects in chromosome segregation [148,149]. PLK1 also contributes to the recruitment and phosphorylation of Sgt1, a co-chaperone for Hsp90 [101]. Hsp90 stabilizes the kinetochore complex by interacting with the MIS12 complex, thereby coordinating the recruitment of NDC80, which interacts with spindle microtubules [150]. In addition, PLK1 interacts with and phosphorylates astrin, which is essential for chromosome alignment and microtubule attachment [151].
Finally, PLK1 may promote stable kinetochore-microtubule attachment by controlling the pool of kinases and phosphatases. As described earlier, BubR1 contains a KARD motif that recruits PP2A, which promotes stable kinetochore-microtubule attachment by counteracting Aurora B activity. PLK1 enhances PP2A interactions by phosphorylating the KARD motif [99,144,145]. However, other studies suggest that PLK1 activates Aurora B by phosphorylating the activating loop of Aurora B [88]. Furthermore, Aurora B may contribute to kinetochore localization of PLK1 and activate PLK1 by phosphorylating the activation loop in the kinase domain [152]. Therefore, regulation of the balance of the phosphorylation state by PLK1 is complex and remains to be fully determined.
7. Concluding Remarks
Since the discovery of PLK1 in the early 1980s, it has been extensively studied using structural, biochemical, and cell-biological approaches. PLK1 contains a conserved kinase domain and PBD domains that recognize the S-[pS/pT]-[P/X] motif of the target protein, required for localization and activation. PLK1 is a master kinase with diverse functions in mitosis. PLK1 localizes to the specific regions in cells through interaction with many proteins, including the PBD domain, and locally phosphorylates the substrates to perform diverse functions. How PLK1 localizes at the kinetochore and regulates functions in mitosis is an important question to be resolved. Many kinetochore proteins, which contain a motif that interacts with the PBD domain, have been suggested to recruit PLK1 to the kinetochore. Here, we reviewed the recent studies that suggest the primary receptors of the PLK1 at the kinetochore. Intriguingly, CENP-U and Bub1 are suggested to be the primary receptors of the PLK1 at the kinetochore, although the relationship between PLK1 and other known PLK1 receptors remains to be clarified. PLK1 contributes to SAC by phosphorylating Knl1, Bub1, BubR1, Mad1, Cdc20, and Mps1. In addition, PLK1 contributes to kinetochore-microtubule attachment via multiple pathways involving PLK1 phosphorylation. Significant progress has been recently made, although many important questions remain. For example, understanding the temporal regulation of PLK1 during mitosis is important. How PLK1 and other kinases, Mps1 and Aurora B, and phosphatases, PP1 and PP2A, work together to balance the phosphorylation state of the kinetochore proteins involved in mitosis remains unresolved. Additionally, since PLK1 localizes at the kinetochore, the kinetochore-localized pool of PLK1 might phosphorylate all those substrates involved in the SAC activation and the kinetochore-microtubule attachment. However, whether or when the kinetochore-localized pool of PLK1 phosphorylates these substrates needs to be investigated. Finally, the identification of additional substrates of PLK1 during SAC activation and normal cell cycle progression will herald the development of new approaches for treating diseases such as cancer.
Funding
This work is supported by grants from the National Research Foundation of Korea (No. NRF-2020RC1C1008596 and NRF-2020R1A4A1019322).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Santaguida, S.; Amon, A. Short- and long-term effects of chromosome mis-segregation and aneuploidy. Nat. Rev. Mol. Cell Biol. 2015, 16, 473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funk, L.C.; Zasadil, L.M.; Weaver, B.A. Living in CIN: Mitotic Infidelity and Its Consequences for Tumor Promotion and Suppression. Dev. Cell 2016, 39, 638–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara-Gonzalez, P.; Pines, J.; Desai, A. Spindle assembly checkpoint activation and silencing at kinetochores. Semin. Cell Dev. Biol. 2021, 117, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Musacchio, A.; Desai, A. A Molecular View of Kinetochore Assembly and Function. Biology 2017, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Saurin, A.T. Kinase and Phosphatase Cross-Talk at the Kinetochore. Front. Cell Dev. Biol. 2018, 6, 62. [Google Scholar] [CrossRef] [Green Version]
- Gelens, L.; Qian, J.B.; Bollen, M.; Saurin, A.T. The Importance of Kinase-Phosphatase Integration: Lessons from Mitosis. Trends Cell Biol. 2018, 28, 6–21. [Google Scholar] [CrossRef] [Green Version]
- Colicino, E.G.; Hehnly, H. Regulating a key mitotic regulator, polo-like kinase 1 (PLK1). Cytoskeleton 2018, 75, 481–494. [Google Scholar] [CrossRef] [Green Version]
- Combes, G.; Alharbi, I.; Braga, L.G.; Elowe, S. Playing polo during mitosis: PLK1 takes the lead. Oncogene 2017, 36, 4819–4827. [Google Scholar] [CrossRef]
- Schmucker, S.; Sumara, I. Molecular dynamics of PLK1 during mitosis. Mol. Cell Oncol. 2014, 1, e954507. [Google Scholar] [CrossRef] [Green Version]
- Korns, J.; Liu, X.Q.; Takiar, V. A review of Plks: Thinking outside the (polo) box. Mol. Carcinog. 2022, 61, 254–263. [Google Scholar] [CrossRef]
- Liu, Z.X.; Sun, Q.R.; Wang, X.S. PLK1, A Potential Target for Cancer Therapy. Transl. Oncol. 2017, 10, 22–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mundt, K.E.; Golsteyn, R.M.; Lane, H.A.; Nigg, E.A. On the regulation and function of human polo-like kinase 1 (PLK1): Effects of overexpression on cell cycle progression. Biochem. Bioph. Res. Co. 1997, 239, 377–385. [Google Scholar] [CrossRef]
- Su, S.; Chhabra, G.; Singh, C.K.; Ndiaye, M.A.; Ahmad, N. PLK1 inhibition-based combination therapies for cancer management. Transl. Oncol. 2022, 16, 101332. [Google Scholar] [CrossRef] [PubMed]
- Ramani, P.; Nash, R.; Sowa-Avugrah, E.; Rogers, C. High levels of polo-like kinase 1 and phosphorylated translationally controlled tumor protein indicate poor prognosis in neuroblastomas. J. Neuro-Oncol. 2015, 125, 103–111. [Google Scholar] [CrossRef]
- Tut, T.G.; Lim, S.H.S.; Dissanayake, I.U.; Descallar, J.; Chua, W.; Ng, W.; de Souza, P.; Shin, J.S.; Lee, C.S. Upregulated Polo-Like Kinase 1 Expression Correlates with Inferior Survival Outcomes in Rectal Cancer. PLoS ONE 2015, 10, e0129313. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.T.; Shi, H.R.; Ren, F.; Liu, H.N.; Zhang, M.H.; Deng, Y.X.; Li, X. Misregulation of polo-like protein kinase 1, P53 and P21(WAF1) in epithelial ovarian cancer suggests poor prognosis. Oncol. Rep. 2015, 33, 1235–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cholewa, B.D.; Liu, X.; Ahmad, N. The role of polo-like kinase 1 in carcinogenesis: Cause or consequence? Cancer Res. 2013, 73, 6848–6855. [Google Scholar] [CrossRef] [Green Version]
- Wood, J.S.; Hartwell, L.H. A Dependent Pathway of Gene Functions Leading to Chromosome Segregation in Saccharomyces-Cerevisiae. J. Cell Biol. 1982, 94, 718–726. [Google Scholar] [CrossRef] [Green Version]
- Hartwell, L.H.; Smith, D. Altered Fidelity of Mitotic Chromosome Transmission in Cell-Cycle Mutants of S-Cerevisiae. Genetics 1985, 110, 381–395. [Google Scholar] [CrossRef]
- Sunkel, C.E.; Glover, D.M. Polo, a Mitotic Mutant of Drosophila Displaying Abnormal Spindle Poles. J. Cell Sci. 1988, 89, 25–38. [Google Scholar] [CrossRef]
- Llamazares, S.; Moreira, A.; Tavares, A.; Girdham, C.; Spruce, B.A.; Gonzalez, C.; Karess, R.E.; Glover, D.M.; Sunkel, C.E. Polo Encodes a Protein-Kinase Homolog Required for Mitosis in Drosophila. Genes Dev. 1991, 5, 2153–2165. [Google Scholar] [CrossRef] [PubMed]
- de Carcer, G.; Manning, G.; Malumbres, M. From Plk1 to Plk5: Functional evolution of polo-like kinases. Cell Cycle 2011, 10, 2255–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowery, D.M.; Lim, D.; Yaffe, M.B. Structure and function of Polo-like kinases. Oncogene 2005, 24, 248–259. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.; Cizmecioglu, O.; Hoffmann, I.; Rhee, K. PLK2 phosphorylation is critical for CPAP function in procentriole formation during the centrosome cycle. EMBO J. 2010, 29, 2395–2406. [Google Scholar] [CrossRef] [Green Version]
- Cizmecioglu, O.; Warnke, S.; Arnold, M.; Duensing, S.; Hoffmann, I. Plk2 regulated centriole duplication is dependent on its localization to the centrioles and a functional polo-box domain. Cell Cycle 2008, 7, 3548–3555. [Google Scholar] [CrossRef]
- Zimmerman, W.C.; Erikson, R.L. Finding Plk3. Cell Cycle 2007, 6, 1314–1318. [Google Scholar] [CrossRef] [Green Version]
- Warnke, S.; Kemmler, S.; Hames, R.S.; Tsai, H.L.; Hoffmann-Rohrer, U.; Fry, A.M.; Hoffmann, I. Polo-like kinase-2 is required for centriole duplication in mammalian cells. Curr. Biol. 2004, 14, 1200–1207. [Google Scholar] [CrossRef] [Green Version]
- Bahassi, M.; Conn, C.; Myer, D.; Hennigan, R.F.; McGowan, C.H.; Sanchez, Y.; Stambrook, P.J. Mammalian Polo-like kinase 3 (Plk3) is a multifunctional protein involved in stress response pathways. Oncogene 2002, 21, 6633–6640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, D.L.; Neel, B.G.; Stevens, R.; Evett, G.; Erikson, R.L. Identification of an Early-Growth-Response Gene Encoding a Novel Putative Protein-Kinase. Mol. Cell Biol. 1992, 12, 4164–4169. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Gao, J.; Dai, W.; Lu, L. Activation of polo-like kinase 3 by hypoxic stresses. J. Biol. Chem. 2008, 283, 25928–25935. [Google Scholar] [CrossRef] [Green Version]
- Winkles, J.A.; Alberts, G.F. Differential regulation of polo-like kinase 1, 2, 3, and 4 gene expression in mammalian cells and tissues. Oncogene 2005, 24, 260–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habedanck, R.; Stierhof, Y.D.; Wilkinson, C.J.; Nigg, E.A. The Polo kinase Plk4 functions in centriole duplication. Nat. Cell Biol. 2005, 7, 1140–1146. [Google Scholar] [CrossRef] [PubMed]
- de Carcer, G.; Escobar, B.; Higuero, A.M.; Garcia, L.; Anson, A.; Perez, G.; Mollejo, M.; Manning, G.; Melendez, B.; Abad-Rodriguez, J.; et al. Plk5, a Polo Box Domain-Only Protein with Specific Roles in Neuron Differentiation and Glioblastoma Suppression. Mol. Cell Biol. 2011, 31, 1225–1239. [Google Scholar] [CrossRef] [Green Version]
- Ohta, M.; Ashikawa, T.; Nozaki, Y.; Kozuka-Hata, H.; Goto, H.; Inagaki, M.; Oyama, M.; Kitagawa, D. Direct interaction of Plk4 with STIL ensures formation of a single procentriole per parental centriole. Nat. Commun. 2014, 5, 5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.P.; Horlin, A.; Hock, B.; Stutte, H.J.; RubsamenWaigmann, H.; Strebhardt, K. Polo-like kinase, a novel marker for cellular proliferation. Am. J. Pathol. 1997, 150, 1165–1172. [Google Scholar]
- Akopyan, K.; Cascales, H.S.; Hukasova, E.; Saurin, A.T.; Mullers, E.; Jaiswal, H.; Hollman, D.A.A.; Kops, G.J.P.L.; Medema, R.H.; Lindqvist, A. Assessing Kinetics from Fixed Cells Reveals Activation of the Mitotic Entry Network at the S/G2 Transition. Mol. Cell 2014, 53, 843–853. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.S.; Oh, D.Y.; Kang, Y.H.; Park, J.E. Self-regulated mechanism of Plk1 localization to kinetochores: Lessons from the Plk1-PBIP1 interaction. Cell Div. 2008, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The protein kinase complement of the human genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef] [Green Version]
- Kothe, M.; Kohls, D.; Low, S.; Coli, R.; Cheng, A.C.; Jacques, S.L.; Johnson, T.L.; Lewis, C.; Loh, C.; Nonomiya, J.; et al. Structure of the catalytic domain of human polo-like kinase 1. Biochemistry 2007, 46, 5960–5971. [Google Scholar] [CrossRef]
- Jang, Y.J.; Lin, C.Y.; Ma, S.; Erikson, R.L. Functional studies on the role of the C-terminal domain of mammalian polo-like kinase. Proc. Natl. Acad. Sci. USA 2002, 99, 1984–1989. [Google Scholar] [CrossRef] [Green Version]
- Park, J.E.; Soung, N.K.; Johmura, Y.; Kang, Y.H.; Liao, C.; Lee, K.H.; Park, C.H.; Nicklaus, M.C.; Lee, K.S. Polo-box domain: A versatile mediator of polo-like kinase function. Cell Mol. Life Sci. 2010, 67, 1957–1970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.S.; Park, J.E.; Kang, Y.H.; Zimmerman, W.; Soung, N.K.; Seong, Y.S.; Kwak, S.J.; Erikson, R.L. Mechanisms of mammalian polo-like kinase 1 (Plk1) localization: Self-priming. Cell Cycle 2008, 7, 141–145. [Google Scholar] [CrossRef]
- Lee, K.S.; Grenfell, T.Z.; Yarm, F.R.; Erikson, R.L. Mutation of the polo-box disrupts localization and mitotic functions of the mammalian polo kinase Plk. Proc. Natl. Acad. Sci. USA 1998, 95, 9301–9306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishi, K.; van Vugt, M.A.T.M.; Okamoto, K.; Hayashi, Y.; Yaffe, M.B. Functional Dynamics of Polo-Like Kinase 1 at the Centrosome. Mol. Cell Biol. 2009, 29, 3134–3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elia, A.E.H.; Cantley, L.C.; Yaffe, M.B. Proteomic screen finds pSer/pThr-binding domain localizing Plk1 to mitotic substrates. Science 2003, 299, 1228–1231. [Google Scholar] [CrossRef] [PubMed]
- Elia, A.E.H.; Rellos, P.; Haire, L.F.; Chao, J.W.; Ivins, F.J.; Hoepker, K.; Mohammad, D.; Cantley, L.C.; Smerdon, S.J.; Yaffe, M.B. The molecular basis for phosphodependent substrate targeting and regulation of Plks by the Polo-box domain. Cell 2003, 115, 83–95. [Google Scholar] [CrossRef]
- Xu, J.; Shen, C.; Wang, T.; Quan, J.M. Structural basis for the inhibition of Polo-like kinase 1. Nat. Struct. Mol. Biol. 2013, 20, 1047–1053. [Google Scholar] [CrossRef]
- Paschal, C.R.; Maciejowski, J.; Jallepalli, P.V. A stringent requirement for Plk1 T210 phosphorylation during K-fiber assembly and chromosome congression. Chromosoma 2012, 121, 565–572. [Google Scholar] [CrossRef] [Green Version]
- Seki, A.; Coppinger, J.A.; Jang, C.Y.; Yates, J.R.; Fang, G.W. Bora and the kinase Aurora A cooperatively activate the kinase Plk1 and control mitotic entry. Science 2008, 320, 1655–1658. [Google Scholar] [CrossRef] [Green Version]
- Macurek, L.; Lindqvist, A.; Lim, D.; Lampson, M.A.; Klompmaker, R.; Freire, R.; Clouin, C.; Taylor, S.S.; Yaffe, M.B.; Medema, R.H. Polo-like kinase-1 is activated by aurora A to promote checkpoint recovery. Nature 2008, 455, U119–U188. [Google Scholar] [CrossRef]
- Carmena, M.; Pinson, X.; Platani, M.; Salloum, Z.; Xu, Z.J.; Clark, A.; MacIsaac, F.; Ogawa, H.; Eggert, U.; Glover, D.M.; et al. The Chromosomal Passenger Complex Activates Polo Kinase at Centromeres. PLoS Biol. 2012, 10, e1001250. [Google Scholar] [CrossRef]
- Bruinsma, W.; Macurek, L.; Freire, R.; Lindqvist, A.; Medema, R.H. Bora and Aurora-A continue to activate Plk1 in mitosis. J. Cell Sci. 2014, 127, 801–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavernier, N.; Thomas, Y.; Vigneron, S.; Maisonneuve, P.; Orlicky, S.; Mader, P.; Regmi, S.G.; Van Hove, L.; Levinson, N.M.; Gasmi-Seabrook, G.; et al. Bora phosphorylation substitutes in trans for T-loop phosphorylation in Aurora A to promote mitotic entry. Nat. Commun. 2021, 12, 1899. [Google Scholar] [CrossRef] [PubMed]
- van Vugt, M.A.T.M.; Bras, A.; Medema, R.H. Polo-like kinase-1 controls recovery from a G2 DNA damage-induced arrest in mammalian cells. Mol. Cell 2004, 15, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Sumara, I.; Gimenez-Abian, J.F.; Gerlich, D.; Hirota, T.; Kraft, C.; de la Torre, C.; Ellenberg, J.; Peters, J.M. Roles of polo-like kinase 1 in the assembly of functional mitotic spindles. Curr. Biol. 2004, 14, 1712–1722. [Google Scholar] [CrossRef] [Green Version]
- Arnaud, L.; Pines, J.; Nigg, E.A. GFP tagging reveals human Polo-like kinase 1 at the kinetochore/centromere region of mitotic chromosomes. Chromosoma 1998, 107, 424–429. [Google Scholar] [CrossRef]
- Abrieu, A.; Brassac, T.; Galas, S.; Fisher, D.; Labbe, J.C.; Doree, M. The polo-like kinase Plx1 is a component of the MPF amplification loop at the G(2)/M-phase transition of the cell cycle in Xenopus eggs. J. Cell Sci. 1998, 111, 1751–1757. [Google Scholar] [CrossRef]
- Lane, H.A.; Nigg, E.A. Antibody microinjection reveals an essential role for human polo-like kinase 1 (Plk1) in the functional maturation of mitotic centrosomes. J. Cell Biol. 1996, 135, 1701–1713. [Google Scholar] [CrossRef] [Green Version]
- Godinho, S.; Tavares, A.A. A role for Drosophila polo protein in chromosome resolution and segregation during mitosis. Cell Cycle 2008, 7, 2529–2534. [Google Scholar] [CrossRef] [Green Version]
- Petronczki, M.; Glotzer, M.; Kraut, N.; Peters, J.M. Polo-like kinase 1 triggers the initiation of cytokinesis in human cells by promoting recruitment of the RhoGEF Ect2 to the central spindle. Dev. Cell 2007, 12, 713–725. [Google Scholar] [CrossRef] [Green Version]
- Burkard, M.E.; Randall, C.L.; Larochelle, S.; Zhang, C.; Shokat, K.M.; Fisher, R.P.; Jallepalli, P.V. Chemical genetics reveals the requirement for Polo-like kinase 1 activity in positioning RhoA and triggering cytokinesis in human cells. Proc. Natl. Acad. Sci. USA 2007, 104, 4383–4388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.H.; Park, J.E.; Yu, L.R.; Soung, N.K.; Yun, S.M.; Bang, J.K.; Seong, Y.S.; Yu, H.T.; Garfield, S.; Veenstra, T.D.; et al. Self-regulated Plk1 recruitment to kinetochores by the Plk1-PBIP1 interaction is critical for proper chromosome segregation. Mol. Cell 2006, 24, 409–422. [Google Scholar] [CrossRef] [PubMed]
- Neef, R.; Preisinger, C.; Sutcliffe, J.; Kopajtich, R.; Nigg, E.A.; Mayer, T.U.; Barr, F.A. Phosphorylation of mitotic kinesin-like protein 2 by polo-like kinase 1 is required for cytokinesis. J. Cell Biol. 2003, 162, 863–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seong, Y.S.; Kamijo, K.; Lee, J.S.; Fernandez, E.; Kuriyama, R.; Miki, T.; Lee, K.S. A spindle checkpoint arrest and a cytokinesis failure by the dominant-negative polo-box domain of plk1 in u-2 OS cells. J. Biol. Chem. 2002, 277, 32282–32293. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.W.; Erikson, E.; Maller, J.L. Mitotic effects of a constitutively active mutant of the Xenopus polo-like kinase Plx1. Mol. Cell Biol. 1999, 19, 8625–8632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, B.A.; Takaki, T.; Petronczki, M.; Glotzer, M. Polo-Like Kinase 1 Directs Assembly of the HsCyk-4 RhoGAP/Ect2 RhoGEF Complex to Initiate Cleavage Furrow Formation. PLoS Biol. 2009, 7, e1000110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkard, M.E.; Maciejowski, J.; Rodriguez-Bravo, V.; Repka, M.; Lowery, D.M.; Clauser, K.R.; Zhang, C.; Shokat, K.M.; Carr, S.A.; Yaffe, M.B.; et al. Plk1 Self-Organization and Priming Phosphorylation of HsCYK-4 at the Spindle Midzone Regulate the Onset of Division in Human Cells. PLoS Biol. 2009, 7, e1000111. [Google Scholar] [CrossRef] [Green Version]
- Lobjois, V.; Jullien, D.; Bouche, J.P.; Ducommun, B. The polo-like kinase 1 regulates CDC25B-dependent mitosis entry. Biochim. Biophy Acta Mol. Cell Res. 2009, 1793, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Lobjois, V.; Froment, C.; Braud, E.; Grimal, F.; Burlet-Schiltz, O.; Ducommun, B.; Bouche, J.P. Study of the docking-dependent PLK1 phosphorylation of the CDC25B phosphatase. Biochem. Biophy Res. Comm. 2011, 410, 87–90. [Google Scholar] [CrossRef]
- Liu, K.; Zheng, M.Y.; Lu, R.; Du, J.X.; Zhao, Q.; Li, Z.G.; Li, Y.W.; Zhang, S.W. The role of CDC25C in cell cycle regulation and clinical cancer therapy: A systematic review. Cancer Cell Int. 2020, 20, 213. [Google Scholar] [CrossRef]
- Gheghiani, L.; Loew, D.; Lombard, B.; Mansfeld, J.; Gavet, O. PLK1 Activation in Late G2 Sets Up Commitment to Mitosis. Cell Rep. 2017, 19, 2060–2073. [Google Scholar] [CrossRef] [PubMed]
- Toyoshima-Morimoti, F.; Taniguchi, E.; Shinya, N.; Iwamatsu, A.; Nishida, E. Polo-like kinase 1 phosphorylates cyclin B1 and targets it to the nucleus during prophase. Nature 2001, 410, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Arai, H.; Nishihara, Y.; Taniguchi, M.; Watanabe, N.; Hunter, T.; Osada, H. M-phase kinases induce phospho-dependent ubiquitination of somatic Wee1 by SCF beta-TrCP. Proc. Natl. Acad. Sci. USA 2004, 101, 4419–4424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.; Malureanu, L.; Huang, J.; Wang, W.; Li, H.; Van Deursen, J.M.; Tindall, D.J.; Chen, J. Plk1-dependent phosphorylation of FoxM1 regulates a transcriptional programme required for mitotic progression. Nat. Cell Biol. 2008, 10, 1076–1082. [Google Scholar] [CrossRef]
- Moshe, Y.; Boulaire, J.; Pagano, M.; Hershko, A. Role of Polo-like kinase in the degradation of early mitotic inhibitor 1, a regulator of the anaphase promoting complex/cyclosome. Proc. Natl. Acad. Sci. USA 2004, 101, 7937–7942. [Google Scholar] [CrossRef] [Green Version]
- Hansen, D.V.; Loktev, A.V.; Ban, K.H.; Jackson, P.K. Plk1 regulates activation of the anaphase promoting complex by phosphorylating and triggering SCF beta TrCP-dependent destruction of the APC inhibitor Emi1. Mol. Biol. Cell 2004, 15, 5623–5634. [Google Scholar] [CrossRef] [Green Version]
- Lenart, P.; Petronczki, M.; Steegmaier, M.; Di Fiore, B.; Lipp, J.J.; Hoffmann, M.; Rettig, W.J.; Kraut, N.; Peters, J.M. The small-molecule inhibitor BI 2536 reveals novel insights into mitotic roles of polo-like kinase 1. Curr. Biol. 2007, 17, 304–315. [Google Scholar] [CrossRef] [Green Version]
- Kraft, C.; Herzog, F.; Gieffers, C.; Mechtler, K.; Hagting, A.; Pines, J.; Peters, J.M. Mitotic regulation of the human anaphase-promoting complex by phosphorylation. EMBO J. 2003, 22, 6598–6609. [Google Scholar] [CrossRef] [Green Version]
- Golan, A.; Yudkovsky, Y.; Hershko, A. The cyclin-ubiquitin ligase activity of cyclosome/APC is jointly activated by protein kinases Cdk1-cyclin B and Plk. J. Biol. Chem. 2002, 277, 15552–15557. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Rhee, K. PLK1 phosphorylation of pericentrin initiates centrosome maturation at the onset of mitosis. J. Cell Biol. 2011, 195, 1093–1101. [Google Scholar] [CrossRef] [Green Version]
- Haren, L.; Stearns, T.; Luders, J. PLK1-DEPENDENT RECRUITMENT OF GAMMA-TUBULIN COMPLEXES TO MITOTIC CENTROSOMES INVOLVES MULTIPLE PCM COMPONENTS. PLoS ONE 2009, 4, e5976. [Google Scholar] [CrossRef]
- Conduit, P.T.; Feng, Z.; Richens, J.H.; Baumbach, J.; Wainman, A.; Bakshi, S.D.; Dobbelaere, J.; Johnson, S.; Lea, S.M.; Raff, J.W. The Centrosome-Specific Phosphorylation of Cnn by Polo/Plk1 Drives Cnn Scaffold Assembly and Centrosome Maturation. Dev. Cell 2014, 28, 659–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casenghi, M.; Barr, F.A.; Nigg, E.A. Phosphorylation of Nlp by Plk1 negatively regulates its dynein-dynactin-dependent targeting to the centrosome. J. Cell Sci. 2005, 118, 5101–5108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshimori, N.; Ohsugi, M.; Yamamoto, T. The Plk1 target Kizuna stabilizes mitotic centrosomes to ensure spindle bipolarity. Nat. Cell Biol. 2006, 8, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Tsou, M.F.B.; Wang, W.J.; George, K.A.; Uryu, K.; Stearns, T.; Jallepalli, P.V. Polo Kinase and Separase Regulate the Mitotic Licensing of Centriole Duplication in Human Cells. Dev. Cell 2009, 17, 344–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, B.; Liu, X.S.; Davis, K.; Liu, X.Q. Plk1 phosphorylation of Orc2 promotes DNA replication under conditions of stress. Mol. Cell Biol. 2011, 31, 4844–4856. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Ali, M.; Duan, W.; Yuan, X.; Garba, F.; Mullen, M.; Sun, B.; Poser, I.; Duan, H.; Lu, J.; et al. Feedback control of PLK1 by Apolo1 ensures accurate chromosome segregation. Cell Rep. 2021, 36, 109343. [Google Scholar] [CrossRef]
- Lee, H.S.; Min, S.; Jung, Y.E.; Chae, S.; Heo, J.; Lee, J.H.; Kim, T.; Kang, H.C.; Nakanish, M.; Cha, S.S.; et al. Spatiotemporal coordination of the RSF1-PLK1-Aurora B cascade establishes mitotic signaling platforms. Nat. Commun. 2021, 12, 5931. [Google Scholar] [CrossRef]
- Lara-Gonzalez, P.; Kim, T.; Oegema, K.; Corbett, K.; Desai, A. A tripartite mechanism catalyzes Mad2-Cdc20 assembly at unattached kinetochores. Science 2021, 371, 64–67. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, M.; Pan, X.; Yuan, X.; Zhou, L.; Yan, L.; Zeng, L.H.; Xu, J.; Yang, B.; Zhang, L.; et al. Bub1 and CENP-U redundantly recruit Plk1 to stabilize kinetochore-microtubule attachments and ensure accurate chromosome segregation. Cell Rep. 2021, 36, 109740. [Google Scholar] [CrossRef]
- Lera, R.F.; Norman, R.X.; Dumont, M.; Dennee, A.; Martin-Koob, J.; Fachinetti, D.; Burkard, M.E. Plk1 protects kinetochore-centromere architecture against microtubule pulling forces. EMBO Rep. 2019, 20, e48711. [Google Scholar] [CrossRef] [PubMed]
- Jones, O.A.; Tiwari, A.; Olukoga, T.; Herbert, A.; Chan, K.L. PLK1 facilitates chromosome biorientation by suppressing centromere disintegration driven by BLM-mediated unwinding and spindle pulling. Nat. Commun. 2019, 10, 2861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, M.; Tanaka, K. Plk1 bound to Bub1 contributes to spindle assembly checkpoint activity during mitosis. Sci. Rep. 2017, 7, 8794. [Google Scholar] [CrossRef]
- Zhuo, X.; Guo, X.; Zhang, X.; Jing, G.; Wang, Y.; Chen, Q.; Jiang, Q.; Liu, J.; Zhang, C. Usp16 regulates kinetochore localization of Plk1 to promote proper chromosome alignment in mitosis. J. Cell Biol. 2015, 210, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Shim, J.; Ji, M.J.; Jung, Y.; Bong, S.M.; Jang, Y.J.; Yoon, E.K.; Lee, S.J.; Kim, K.G.; Kim, Y.H.; et al. The condensin component NCAPG2 regulates microtubule-kinetochore attachment through recruitment of Polo-like kinase 1 to kinetochores. Nat. Commun. 2014, 5, 4588. [Google Scholar] [CrossRef] [Green Version]
- Kakeno, M.; Matsuzawa, K.; Matsui, T.; Akita, H.; Sugiyama, I.; Ishidate, F.; Nakano, A.; Takashima, S.; Goto, H.; Inagaki, M.; et al. Plk1 phosphorylates CLIP-170 and regulates its binding to microtubules for chromosome alignment. Cell Struct. Funct. 2014, 39, 45–59. [Google Scholar] [CrossRef] [Green Version]
- Amin, M.A.; Itoh, G.; Iemura, K.; Ikeda, M.; Tanaka, K. CLIP-170 recruits PLK1 to kinetochores during early mitosis for chromosome alignment. J. Cell Sci. 2014, 127 Pt 13, 2818–2824. [Google Scholar] [CrossRef] [Green Version]
- Yeh, T.Y.; Kowalska, A.K.; Scipioni, B.R.; Cheong, F.K.; Zheng, M.; Derewenda, U.; Derewenda, Z.S.; Schroer, T.A. Dynactin helps target Polo-like kinase 1 to kinetochores via its left-handed beta-helical p27 subunit. EMBO J. 2013, 32, 1023–1035. [Google Scholar] [CrossRef] [Green Version]
- Suijkerbuijk, S.J.; Vleugel, M.; Teixeira, A.; Kops, G.J. Integration of kinase and phosphatase activities by BUBR1 ensures formation of stable kinetochore-microtubule attachments. Dev. Cell 2012, 23, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Maia, A.R.; Garcia, Z.; Kabeche, L.; Barisic, M.; Maffini, S.; Macedo-Ribeiro, S.; Cheeseman, I.M.; Compton, D.A.; Kaverina, I.; Maiato, H. Cdk1 and Plk1 mediate a CLASP2 phospho-switch that stabilizes kinetochore-microtubule attachments. J. Cell Biol. 2012, 199, 285–301. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.S.; Song, B.; Tang, J.B.; Liu, W.Y.; Kuang, S.H.; Liu, X.Q. Plk1 Phosphorylates Sgt1 at the Kinetochores To Promote Timely Kinetochore-Microtubule Attachment. Mol. Cell Biol. 2012, 32, 4053–4067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishino, M.; Kurasawa, Y.; Evans, R.; Lin, S.H.; Brinkley, B.R.; Yu-Lee, L.Y. NudC is required for Plk1 targeting to the kinetochore and chromosome congression. Curr. Biol. 2006, 16, 1414–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, H.; Kiyono, T.; Tomono, Y.; Kawajiri, A.; Urano, T.; Furukawa, K.; Nigg, E.A.; Inagaki, M. Complex formation of Plk1 and INCENP required for metaphase-anaphase transition. Nat. Cell Biol. 2006, 8, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Pesenti, M.E.; Maffini, S.; Carmignani, S.; Hedtfeld, M.; Petrovic, A.; Srinivasamani, A.; Bange, T.; Musacchio, A. BUB1 and CENP-U, Primed by CDK1, Are the Main PLK1 Kinetochore Receptors in Mitosis. Mol. Cell 2021, 81, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Park, Y.Y.; Cho, M.Y.; Chae, S.; Yoo, Y.S.; Kwon, M.H.; Lee, C.W.; Cho, H. The chromatin remodeller RSF1 is essential for PLK1 deposition and function at mitotic kinetochores. Nat. Commun. 2015, 6, 7904. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, X.S.; Yang, X.; Wang, Y.; Wang, Y.; Turner, J.R.; Liu, X. Phosphorylation of CLIP-170 by Plk1 and CK2 promotes timely formation of kinetochore-microtubule attachments. EMBO J. 2010, 29, 2953–2965. [Google Scholar] [CrossRef] [Green Version]
- Pouwels, J.; Kukkonen, A.M.; Lan, W.; Daum, J.R.; Gorbsky, G.J.; Stukenberg, T.; Kallio, M.J. Shugoshin 1 plays a central role in kinetochore assembly and is required for kinetochore targeting of Plk1. Cell Cycle 2007, 6, 1579–1585. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Aumais, J.P.; Liu, X.Q.; Yu-Lee, L.Y.; Erikson, R.L. A Role for Plk1 phosphorylation of NudC in cytokinesis. Dev. Cell 2003, 5, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Morrow, C.J.; Tighe, A.; Johnson, V.L.; Scott, M.I.; Ditchfield, C.; Taylor, S.S. Bub1 and aurora B cooperate to maintain BubR1-mediated inhibition of APC/CCdc20. J. Cell Sci. 2005, 118 Pt 16, 3639–3652. [Google Scholar] [CrossRef] [Green Version]
- Foley, E.A.; Maldonado, M.; Kapoor, T.M. Formation of stable attachments between kinetochores and microtubules depends on the B56-PP2A phosphatase. Nat. Cell Biol. 2011, 13, 1265–1271. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.L.; Fadel, M.D.; Cheeseman, I.M. Differential requirements for the CENP-O complex reveal parallel PLK1 kinetochore recruitment pathways. Mol. Biol. Cell 2021, 32, 712–721. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Gartner, A. Bub1 kinase in the regulation of mitosis. Anim. Cells Syst. 2021, 25, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, Y.; Yang, C.H.; Tanno, Y.; Watanabe, Y. MPS1/Mph1 phosphorylates the kinetochore protein KNL1/Spc7 to recruit SAC components. Nat. Cell Biol. 2012, 14, 746–752. [Google Scholar] [CrossRef]
- Shepperd, L.A.; Meadows, J.C.; Sochaj, A.M.; Lancaster, T.C.; Zou, J.; Buttrick, G.J.; Rappsilber, J.; Hardwick, K.G.; Millar, J.B.A. Phosphodependent Recruitment of Bub1 and Bub3 to Spc7/KNL1 by Mph1 Kinase Maintains the Spindle Checkpoint. Curr. Biol. 2012, 22, 891–899. [Google Scholar] [CrossRef] [Green Version]
- London, N.; Ceto, S.; Ranish, J.A.; Biggins, S. Phosphoregulation of Spc105 by Mps1 and PP1 Regulates Bub1 Localization to Kinetochores. Curr. Biol. 2012, 22, 900–906. [Google Scholar] [CrossRef] [Green Version]
- Faesen, A.C.; Thanasoula, M.; Maffini, S.; Breit, C.; Muller, F.; van Gerwen, S.; Bange, T.; Musacchio, A. Basis of catalytic assembly of the mitotic checkpoint complex. Nature 2017, 542, 498–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santaguida, S.; Tighe, A.; D’Alise, A.M.; Taylor, S.S.; Musacchio, A. Dissecting the role of MPS1 in chromosome biorientation and the spindle checkpoint through the small molecule inhibitor reversine. J. Cell Biol. 2010, 190, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Maciejowski, J.; George, K.A.; Terret, M.E.; Zhang, C.; Shokat, K.M.; Jallepalli, P.V. Mps1 directs the assembly of Cdc20 inhibitory complexes during interphase and mitosis to control M phase timing and spindle checkpoint signaling. J. Cell Biol. 2010, 190, 89–100. [Google Scholar] [CrossRef]
- Hewitt, L.; Tighe, A.; Santaguida, S.; White, A.M.; Jones, C.D.; Musacchio, A.; Green, S.; Taylor, S.S. Sustained Mps1 activity is required in mitosis to recruit O-Mad2 to the Mad1-C-Mad2 core complex. J. Cell Biol. 2010, 190, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Abrieu, A.; Magnaghi-Jaulin, L.; Kahana, J.A.; Peter, M.; Castro, A.; Vigneron, S.; Lorca, T.; Cleveland, D.W.; Labbe, J.C. Mps1 is a kinetochore associated kinase essential for the vertebrate mitotic checkpoint. Cell 2001, 106, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Sacristan, C.; Ahmad, M.U.D.; Keller, J.; Fermie, J.; Groenewold, V.; Tromer, E.; Fish, A.; Melero, R.; Carazo, J.M.; Klumperman, J.; et al. Dynamic kinetochore size regulation promotes microtubule capture and chromosome biorientation in mitosis. Nat. Cell Biol. 2018, 20, 800–810. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Rodriguez, J.A.; Lewis, C.; McKinley, K.L.; Sikirzhytski, V.; Corona, J.; Maciejowski, J.; Khodjakov, A.; Cheeseman, I.M.; Jallepalli, P.V. Distinct Roles of RZZ and Bub1-KNL1 in Mitotic Checkpoint Signaling and Kinetochore Expansion. Curr. Biol. 2018, 28, 3422–3429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, Z.; von Schubert, C.; Korner, R.; Santamaria, A.; Elowe, S.; Nigg, E.A. Quantitative Mass Spectrometry Analysis Reveals Similar Substrate Consensus Motif for Human Mps1 Kinase and Plk1. PLoS ONE 2011, 6, e18793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennrich, M.L.; Marino, F.; Groenewold, V.; Kops, G.J.P.L.; Mohammed, S.; Heck, A.J.R. Universal Quantitative Kinase Assay Based on Diagonal SCX Chromatography and Stable Isotope Dimethyl Labeling Provides High-definition Kinase Consensus Motifs for PKA and Human Mps1. J. Proteome Res. 2013, 12, 2214–2224. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Toyoshima-Morimoto, F.; Taniguchi, E.; Nishida, E. Identification of a consensus motif for Plk (Polo-like kinase) phosphorylation reveals Myt1 as a Plk1 substrate. J. Biol. Chem. 2003, 278, 25277–25280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppermann, F.S.; Grundner-Culemann, K.; Kumar, C.; Gruss, O.J.; Jallepalli, P.V.; Daub, H. Combination of Chemical Genetics and Phosphoproteomics for Kinase Signaling Analysis Enables Confident Identification of Cellular Downstream Targets. Mol. Cell Proteom 2012, 11, O111.012351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamaria, A.; Wang, B.; Elowe, S.; Malik, R.; Zhang, F.; Bauer, M.; Schmidt, A.; Sillje, H.H.; Korner, R.; Nigg, E.A. The Plk1-dependent phosphoproteome of the early mitotic spindle. Mol. Cell Proteom. 2011, 10, M110.004457. [Google Scholar] [CrossRef] [Green Version]
- Espeut, J.; Lara-Gonzalez, P.; Sassine, M.; Shiau, A.K.; Desai, A.; Abrieu, A. Natural Loss of Mps1 Kinase in Nematodes Uncovers a Role for Polo-like Kinase 1 in Spindle Checkpoint Initiation. Cell Rep. 2015, 12, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Ji, Z.J.; Gao, H.S.; Jia, L.Y.; Li, B.; Yu, H.T. A sequential multi-target Mps1 phosphorylation cascade promotes spindle checkpoint signaling. Elife 2017, 6, e22513. [Google Scholar] [CrossRef]
- London, N.; Biggins, S. Mad1 kinetochore recruitment by Mps1-mediated phosphorylation of Bub1 signals the spindle checkpoint. Genes Dev. 2014, 28, 140–152. [Google Scholar] [CrossRef] [Green Version]
- Piano, V.; Alex, A.; Stege, P.; Maffini, S.; Stoppiello, G.A.; Veld, P.J.H.I.; Vetter, I.R.; Musacchio, A. CDC20 assists its catalytic incorporation in the mitotic checkpoint complex. Science 2021, 371, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.Y.; Li, B.; Yu, H.T. The Bub1-Plk1 kinase complex promotes spindle checkpoint signalling through Cdc20 phosphorylation. Nat. Commun. 2016, 7, 10818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T. Delay in the Cell Cycle by a Single Unattached Kinetochore. J. Life Sci. 2022, 32, 161–166. [Google Scholar]
- von Schubert, C.; Cubizolles, F.; Bracher, J.M.; Sliedrecht, T.; Kops, G.J.P.L.; Nigg, E.A. Plk1 and Mps1 Cooperatively Regulate the Spindle Assembly Checkpoint in Human Cells. Cell Rep. 2015, 12, 66–78. [Google Scholar] [CrossRef] [Green Version]
- Espert, A.; Uluocak, P.; Bastos, R.N.; Mangat, D.; Graab, P.; Gruneberg, U. PP2A-B56 opposes Mps1 phosphorylation of Knl1 and thereby promotes spindle assembly checkpoint silencing. J. Cell Biol. 2014, 206, 833–842. [Google Scholar] [CrossRef]
- Braga, L.G.; Cisneros, A.F.; Mathieu, M.M.; Clerc, M.; Garcia, P.; Lottin, B.; Garand, C.; Thebault, P.; Landry, C.R.; Elowe, S. BUBR1 Pseudokinase Domain Promotes Kinetochore PP2A-B56 Recruitment, Spindle Checkpoint Silencing, and Chromosome Alignment. Cell Rep. 2020, 33, 108397. [Google Scholar] [CrossRef]
- Nijenhuis, W.; Vallardi, G.; Teixeira, A.; Kops, G.J.P.L.; Saurin, A.T. Negative feedback at kinetochores underlies a responsive spindle checkpoint signal. Nat. Cell Biol. 2014, 16, 1257–1269. [Google Scholar] [CrossRef] [Green Version]
- Espeut, J.; Cheerambathur, D.K.; Krenning, L.; Oegema, K.; Desai, A. Microtubule binding by KNL-1 contributes to spindle checkpoint silencing at the kinetochore. J. Cell Biol. 2012, 196, 469–482. [Google Scholar] [CrossRef]
- Rosenberg, J.S.; Cross, F.R.; Funabiki, H. KNL1/Spc105 Recruits PP1 to Silence the Spindle Assembly Checkpoint. Curr. Biol. 2011, 21, 942–947. [Google Scholar] [CrossRef] [Green Version]
- Meadows, J.C.; Shepperd, L.A.; Vanoosthuyse, V.; Lancaster, T.C.; Sochaj, A.M.; Buttrick, G.J.; Hardwick, K.G.; Millar, J.B.A. Spindle Checkpoint Silencing Requires Association of PP1 to Both Spc7 and Kinesin-8 Motors. Dev. Cell 2011, 20, 739–750. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Vleugel, M.; Backer, C.B.; Hori, T.; Fukagawa, T.; Cheeseman, I.M.; Lampson, M.A. Regulated targeting of protein phosphatase 1 to the outer kinetochore by KNL1 opposes Aurora B kinase. J. Cell Biol. 2010, 188, 809–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanoosthuyse, V.; Hardwick, K.G. A Novel Protein Phosphatase 1-Dependent Spindle Checkpoint Silencing Mechanism. Curr. Biol. 2009, 19, 1176–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinsky, B.A.; Nelson, C.R.; Biggins, S. Protein Phosphatase 1 Regulates Exit from the Spindle Checkpoint in Budding Yeast. Curr. Biol. 2009, 19, 1182–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruse, T.; Zhang, G.; Larsen, M.S.Y.; Lischetti, T.; Streicher, W.; Nielsen, T.K.; Bjorn, S.P.; Nilsson, J. Direct binding between BubR1 and B56-PP2A phosphatase complexes regulate mitotic progression. J. Cell Sci. 2013, 126, 1086–1092. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Raetz, E.A.; Kitagawa, M.; Virshup, D.M.; Lee, S.H. BUBR1 recruits PP2A via the B56 family of targeting subunits to promote chromosome congression. Biol. Open 2013, 2, 479–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordeiro, M.H.; Smith, R.J.; Saurin, A.T. Kinetochore phosphatases suppress autonomous Polo-like kinase 1 activity to control the mitotic checkpoint. J. Cell Biol. 2020, 219, e202002020. [Google Scholar] [CrossRef] [PubMed]
- Colnaghi, R.; Wheatley, S.P. Liaisons between survivin and Plk1 during cell division and cell death. J. Biol. Chem. 2010, 285, 22592–22604. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.H.; Park, C.H.; Kim, T.S.; Soung, N.K.; Bang, J.K.; Kim, B.Y.; Park, J.E.; Lee, K.S. Mammalian Polo-like Kinase 1-dependent Regulation of the PBIP1-CENP-Q Complex at Kinetochores. J. Biol. Chem. 2011, 286, 19744–19757. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Park, J.E.; Kim, T.S.; Kang, Y.H.; Soung, N.K.; Zhou, M.; Kim, N.H.; Bang, J.K.; Lee, K.S. Mammalian Polo-like Kinase 1 (Plk1) Promotes Proper Chromosome Segregation by Phosphorylating and Delocalizing the PBIP1.CENP-Q Complex from Kinetochores. J. Biol. Chem. 2015, 290, 8569–8581. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.E.; Kaplan, K.B. Hsp90-Sgt1 and Skp1 target human Mis12 complexes to ensure efficient formation of kinetochore-microtubule binding sites. J. Cell Biol. 2010, 189, 261–274. [Google Scholar] [CrossRef]
- Geraghty, Z.; Barnard, C.; Uluocak, P.; Gruneberg, U. The association of Plk1 with the astrin-kinastrin complex promotes formation and maintenance of a metaphase plate. J. Cell Sci. 2021, 134, jcs251025. [Google Scholar] [CrossRef] [PubMed]
- Kachaner, D.; Pinson, X.; Ben El Kadhi, K.; Normandin, K.; Talje, L.; Lavoie, H.; Lepine, G.; Carreno, S.; Kwok, B.H.; Hickson, G.R.; et al. Interdomain allosteric regulation of Polo kinase by Aurora B and Map205 is required for cytokinesis. J. Cell Biol. 2014, 207, 201–211. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Domain organization of PLK1. A schematic diagram of human PLK1 protein and its domain organization. PLK1 contains the serine/threonine kinase domain (KD: orange), the polo-box domain (PBD: green), and the linker between two domains. The PBD consists of polo-box 1 and 2 (PB1 and PB2: yellow). PLK1 activation requires the phosphorylation of conserved Thr residues within the T-Loop.
Figure 1.
Domain organization of PLK1. A schematic diagram of human PLK1 protein and its domain organization. PLK1 contains the serine/threonine kinase domain (KD: orange), the polo-box domain (PBD: green), and the linker between two domains. The PBD consists of polo-box 1 and 2 (PB1 and PB2: yellow). PLK1 activation requires the phosphorylation of conserved Thr residues within the T-Loop.

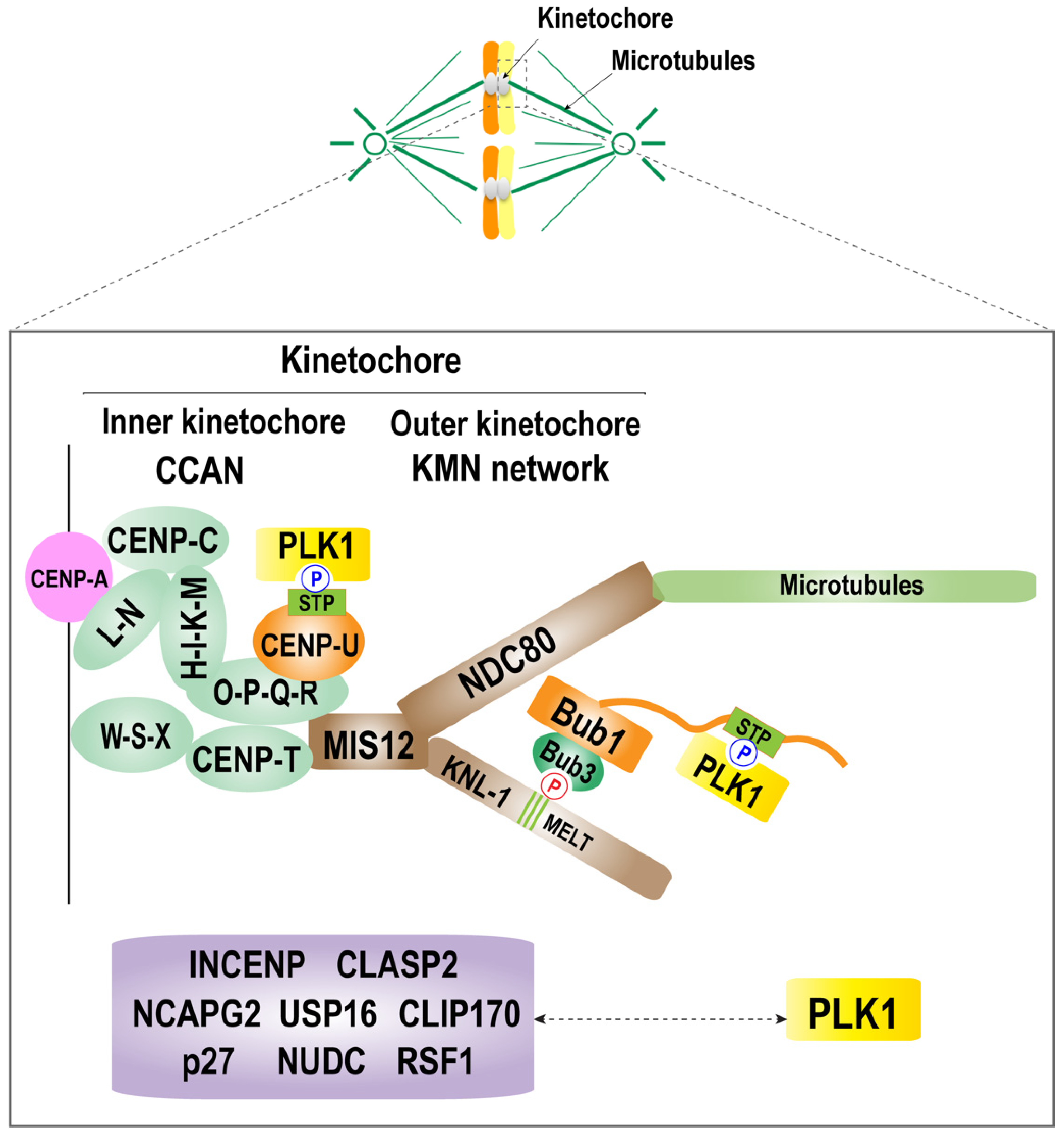
Figure 2.
Molecular mechanism of PLK1 localization at the kinetochore. A schematic diagram of the kinetochore structure and the mechanism of kinetochore recruitment of PLK1. The kinetochore consists of an inner and an outer kinetochore. The inner kinetochore is composed of a complex of CCAN, CENP-C, CENP-HIK-LN, CENP-T, and CENP-QU. The outer kinetochore is composed of KNL1 complex, Mis12 complex, and the NDC80 complex (KMN network). PLK1 is recruited to the kinetochore by interacting with CENP-U and Bub1, which are located in the inner kinetochore and outer kinetochore, respectively. There are other known PLK1 receptors, including the kinetochore components Bub1, BubR1, and CENP-U (also called PBIP1), and other proteins, including nuclear distribution protein C (NUDC), CLIP-170, dynactin subunit p27, INCENP, CLASP2, Survivin, NCAPG2, USP16, and RSF1. The relationship between PLK1 and these components during mitosis requires further characterization.
Figure 2.
Molecular mechanism of PLK1 localization at the kinetochore. A schematic diagram of the kinetochore structure and the mechanism of kinetochore recruitment of PLK1. The kinetochore consists of an inner and an outer kinetochore. The inner kinetochore is composed of a complex of CCAN, CENP-C, CENP-HIK-LN, CENP-T, and CENP-QU. The outer kinetochore is composed of KNL1 complex, Mis12 complex, and the NDC80 complex (KMN network). PLK1 is recruited to the kinetochore by interacting with CENP-U and Bub1, which are located in the inner kinetochore and outer kinetochore, respectively. There are other known PLK1 receptors, including the kinetochore components Bub1, BubR1, and CENP-U (also called PBIP1), and other proteins, including nuclear distribution protein C (NUDC), CLIP-170, dynactin subunit p27, INCENP, CLASP2, Survivin, NCAPG2, USP16, and RSF1. The relationship between PLK1 and these components during mitosis requires further characterization.

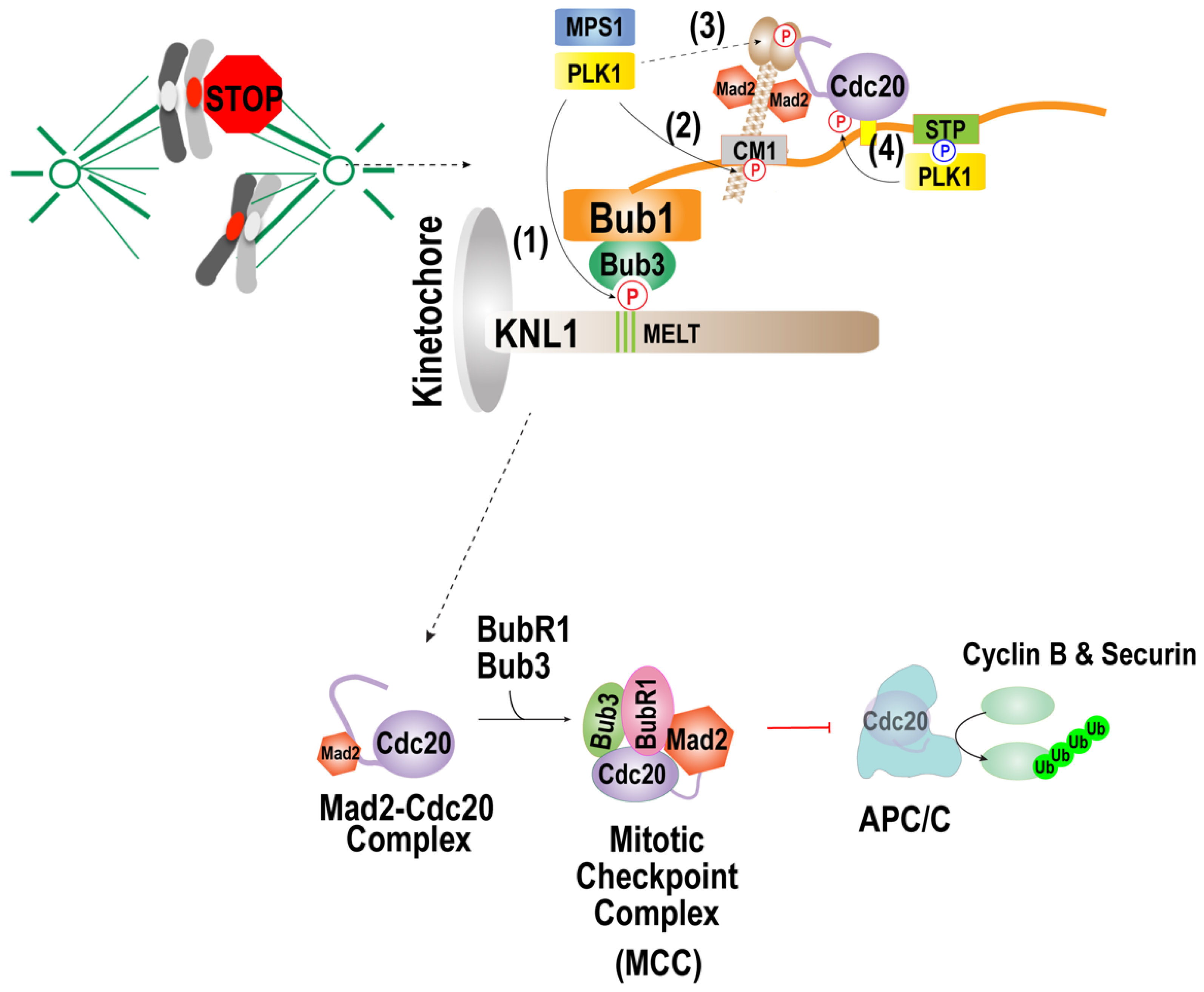
Figure 3.
Function of the PLK1 in spindle assembly checkpoint activation. Reprinted/adapted with permission from Ref. [133], 2021, Taekyung Kim. When kinetochores are not attached to the microtubules, PLK1 activates the checkpoint by phosphorylating (1) MELT motifs of KNL1 and (2) the CM1 motif of Bub1, (3) C-terminal region of Mad1, and (4) Cdc20. The PLK1 can substitute Mps1 kinase in phosphorylating MELT motifs of KNL1, which promotes kinetochore localization of the Bub1/Bub3 complex and the CM1 motif of bub1, mediating the interaction with Mad1. PLK-1 in C. elegans was shown to phosphorylate the C-terminal region of Mad1, which promotes the interaction with Mad1. PLK1 that interacts with Bub1 activates the MCC formation by phosphorylating Cdc20.
Figure 3.
Function of the PLK1 in spindle assembly checkpoint activation. Reprinted/adapted with permission from Ref. [133], 2021, Taekyung Kim. When kinetochores are not attached to the microtubules, PLK1 activates the checkpoint by phosphorylating (1) MELT motifs of KNL1 and (2) the CM1 motif of Bub1, (3) C-terminal region of Mad1, and (4) Cdc20. The PLK1 can substitute Mps1 kinase in phosphorylating MELT motifs of KNL1, which promotes kinetochore localization of the Bub1/Bub3 complex and the CM1 motif of bub1, mediating the interaction with Mad1. PLK-1 in C. elegans was shown to phosphorylate the C-terminal region of Mad1, which promotes the interaction with Mad1. PLK1 that interacts with Bub1 activates the MCC formation by phosphorylating Cdc20.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, T. Recent Progress on the Localization of PLK1 to the Kinetochore and Its Role in Mitosis. Int. J. Mol. Sci. 2022, 23, 5252. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095252
AMA Style
Kim T. Recent Progress on the Localization of PLK1 to the Kinetochore and Its Role in Mitosis. International Journal of Molecular Sciences. 2022; 23(9):5252. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095252
Chicago/Turabian StyleKim, Taekyung. 2022. "Recent Progress on the Localization of PLK1 to the Kinetochore and Its Role in Mitosis" International Journal of Molecular Sciences 23, no. 9: 5252. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23095252
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.