
Bempedoic Acid Restores Liver H2S Production in a Female Sprague-Dawley Rat Dietary Model of Non-Alcoholic Fatty Liver
 , , , , , , , and
, , , , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Reduction of H2S Production in the Steatotic Livers of Female Sprague-Dawley Rats Is Reversed by Bempedoic Acid Administration, Restoring CBS and CSE Expression
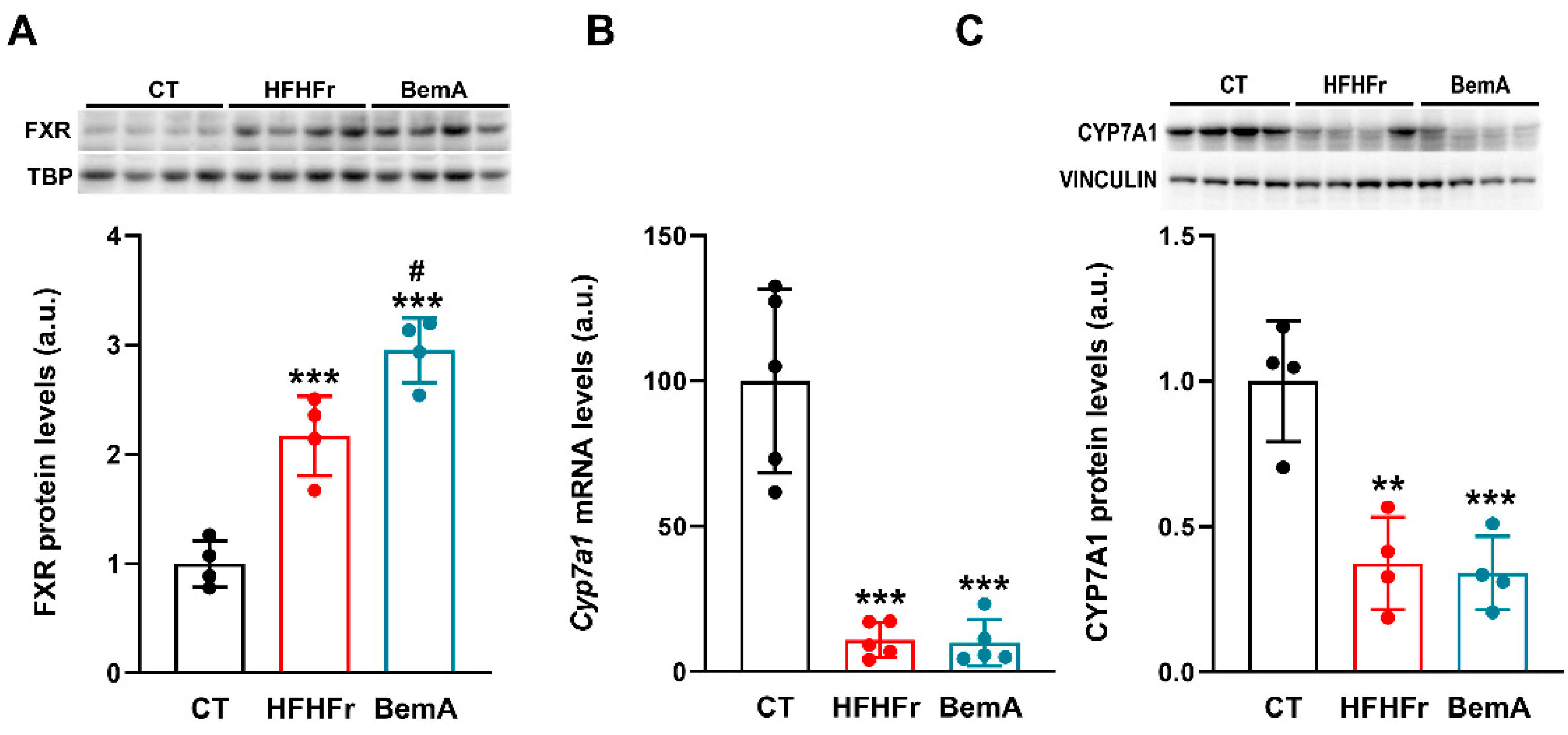
2.2. BemA Administration Increases the Expression and Activity of FXR in the Livers of HFHFr-Fed Female Sprague-Dawley Rats
2.3. BemA Administration Improves the Ratio of Phophor-Ser571/Total PGC1α (Peroxisome Proliferator Activator Receptor Gamma Coactivator-1α) in the Livers of HFHFr-Fed Female Sprague-Dawley Rats
3. Discussion
4. Material and Methods
4.1. Animals and Experimental Design
4.2. Sample Preparation
4.3. Serum Analytes
4.4. Liver Bile Acids Determination
4.5. Determination of H2S Production by Liver Tissue
4.6. RNA Extraction and Quantitative RT-qPCR Analysis
4.7. Protein Extraction and Western Blot
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, Z.L.; Zhao, L.; Ma, T.; Osama, A.; Shen, T.; He, Y.; Fang, J. Progress and perspective on hydrogen sulfide donors and their biomedical applications. Med. Res. Rev. 2022, 42, 1930–1977. [Google Scholar] [CrossRef] [PubMed]
- Comas, F.; Moreno-Navarrete, J.M. The Impact of H2S on Obesity-Associated Metabolic Disturbances. Antioxidants 2021, 10, 633. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Ding, Y.; Liu, H.; Sun, M.; Chen, C.; Yang, Y.; Wang, H. The Role of Hydrogen Sulfide Regulation of Autophagy in Liver Disorders. Int. J. Mol. Sci. 2022, 23, 4035. [Google Scholar] [CrossRef] [PubMed]
- Werge, M.P.; McCann, A.; Galsgaard, E.D.; Holst, D.; Bugge, A.; Wewer Albrechtsen, N.J.; Gluud, L.L. The role of the transsulfuration pathway in non-alcoholic fatty liver disease. J. Clin. Med. 2021, 10, 1081. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.D.; Snyder, S.H. H 2S signalling through protein sulfhydration and beyond. Nat. Rev. Mol. Cell Biol. 2012, 13, 499–507. [Google Scholar] [CrossRef]
- Nguyen, T.T.P.; Kim, D.Y.; Lee, Y.G.; Lee, Y.S.; Truong, X.T.; Lee, J.H.; Song, D.K.; Kwon, T.K.; Park, S.H.; Jung, C.H.; et al. SREBP-1c impairs ULK1 sulfhydration-mediated autophagic flux to promote hepatic steatosis in high-fat-diet-fed mice. Mol. Cell 2021, 81, 3820–3832.e7. [Google Scholar] [CrossRef]
- Renga, B.; Mencarelli, A.; Migliorati, M.; Distrutti, E.; Fiorucci, S. Bile-acid-activated farnesoid X receptor regulates hydrogen sulfide production and hepatic microcirculation. World J. Gastroenterol. 2009, 15, 2097–2108. [Google Scholar] [CrossRef]
- Renga, B.; Bucci, M.; Cipriani, S.; Carino, A.; Monti, M.C.; Zampella, A.; Gargiulo, A.; di Villa Bianca, R.D.; Distrutti, E.; Fiorucci, S. Cystathionine γ-lyase, a H2S-generating enzyme, is a GPBAR1-regulated gene and contributes to vasodilation caused by secondary bile acids. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H114–H126. [Google Scholar] [CrossRef] [Green Version]
- El-Agroudy, N.N.; Kurzbach, A.; Rodionov, R.N.; O’Sullivan, J.; Roden, M.; Birkenfeld, A.L.; Pesta, D.H. Are Lifestyle Therapies Effective for NAFLD Treatment? Trends Endocrinol. Metab. 2019, 30, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Pydyn, N.; Miękus, K.; Jura, J.; Kotlinowski, J. New therapeutic strategies in nonalcoholic fatty liver disease: A focus on promising drugs for nonalcoholic steatohepatitis. Pharmacol. Rep. 2020, 72, 1–12. [Google Scholar] [CrossRef]
- Santhekadur, P.K.; Kumar, D.P.; Sanyal, A.J. Preclinical models of non-alcoholic fatty liver disease. J. Hepatol. 2018, 68, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Fauste, E.; Rodrigo, S.; Aguirre, R.; Donis, C.; Rodríguez, L.; Álvarez-Millán, J.J.; Panadero, M.I.; Otero, P.; Bocos, C. Maternal Fructose Intake Increases Liver H2S Synthesis but Exarcebates its Fructose-Induced Decrease in Female Progeny. Mol. Nutr. Food Res. 2020, 64, e2000628. [Google Scholar] [CrossRef]
- Fauste, E.; Rodrigo, S.; Aguirre, R.; Rodríguez, L.; Álvarez-Millán, J.J.; Panadero, M.I.; Otero, P.; Bocos, C. Liquid carbohydrate intake modifies transsulfuration pathway both in pregnant rats and in their male descendants. Clin. Investig. Arterioscler. 2021, 33, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.; Akamnonu, I.; Yang, T. Bempedoic Acid: A cholesterol lowering agent with a novel mechanism of action. Expert Rev. Clin. Pharmacol. 2021, 14, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhang, L.; Xu, S.; Shen, A.Z. ATP-citrate lyase (ACLY) in lipid metabolism and atherosclerosis: An updated review. Prog. Lipid Res. 2020, 77, 101006. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, A.M.; Bentanachs, R.; Sala-Vila, A.; Lázaro, I.; Rodríguez-Morató, J.; Sánchez, R.M.; Alegret, M.; Roglans, N.; Laguna, J.C. ChREBP-driven DNL and PNPLA3 Expression Induced by Liquid Fructose are Essential in the Production of Fatty Liver and Hypertriglyceridemia in a High-Fat Diet-Fed Rat Model. Mol. Nutr. Food Res. 2022, 66, 2101115. [Google Scholar] [CrossRef] [PubMed]
- Bentanachs, R.; Velázquez, A.M.; Sánchez, R.M.; Alegret, M.; Laguna, J.C.; Roglans, N. Bempedoic acid as a PPARα activator: New perspectives for hepatic steatosis treatment in a female rat experimental model. Clin. Investig. Arterioscler. 2022, 34, 57–67. [Google Scholar] [CrossRef]
- Velázquez, A.M.; Bentanachs, R.; Sala-Vila, A.; Lázaro, I.; Rodríguez-Morató, J.; Sánchez, R.M.; Laguna, J.C.; Roglans, N.; Alegret, M. KHK, PNPLA3 and PPAR as novel targets for the anti-steatotic action of bempedoic acid. Biomedicines 2022, 10, 1517. [Google Scholar] [CrossRef]
- Chiang, J.Y.L.; Ferrell, J.M. Up to date on cholesterol 7 alpha-hydroxylase (CYP7A1) in bile acid synthesis. Liver Res. 2020, 4, 47–63. [Google Scholar] [CrossRef]
- Jannig, P.R.; Dumesic, P.A.; Spiegelman, B.M.; Ruas, J.L. SnapShot: Regulation and biology of PGC-1 alpha. Cell 2022, 185, 1444–1444.e1. [Google Scholar] [CrossRef]
- Liu, G.Y.; Sabatini, D.M. mTOR at the nexus of nutrition, growth, ageing and disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Mateus, I.; Prip-Buus, C. Hydrogen sulphide in liver glucose/lipid metabolism and non-alcoholic fatty liver disease. Eur. J. Clin. Investig. 2022, 52, e13680. [Google Scholar] [CrossRef] [PubMed]
- Morrow, M.R.; Batchuluun, B.; Wu, J.; Ahmadi, E.; Leroux, J.M.; Mohammadi-Shemirani, P.; Desjardins, E.M.; Wang, Z.; Tsakiridis, E.E.; Lavoie, D.C.T.; et al. Inhibition of ATP-citrate lyase improves NASH, liver fibrosis, and dyslipidemia. Cell Metab. 2022, 34, 919–936.e8. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, N.; Fu, C.; Pappin, D.J.; Tonks, N.K. Biochemistry: H 2S-induced sulfhydration of the phosphatase PTP1B and its role in the endoplasmic reticulum stress response. Sci. Signal. 2011, 4, ra86. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Marcos, P.J.; Auwerx, J. Regulation of PGC-1α, a nodal regulator of mitochondrial biogenesis. Am. J. Clin. Nutr. 2011, 93, 884–890. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Liao, C.; Quan, J.; Cheng, C.; Zhao, X.; Bode, A.M.; Cao, Y. Posttranslational regulation of PGC-1α and its implication in cancer metabolism. Int. J. Cancer 2019, 145, 1475–1483. [Google Scholar] [CrossRef]
- Green, C.L.; Lamming, D.W.; Fontana, L. Molecular mechanisms of dietary restriction promoting health and longevity. Nat. Rev. Mol. Cell Biol. 2022, 23, 56–73. [Google Scholar] [CrossRef]
- Baena, M.; Sangüesa, G.; Hutter, N.; Sánchez, R.M.; Roglans, N.; Laguna, J.C.; Alegret, M. Fructose supplementation impairs rat liver autophagy through mTORC activation without inducing endoplasmic reticulum stress. Biochim. Biophys. Acta 2015, 1851, 107–116. [Google Scholar] [CrossRef]
- Hine, C.; Mitchell, J. Endpoint or Kinetic Measurement of Hydrogen Sulfide Production Capacity in Tissue Extracts. Bio-Protocol 2017, 7, e2382. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Minkler, P.; Grove, D.; Wang, R.; Willard, B.; Dweik, R.; Hine, C. Non-enzymatic hydrogen sulfide production from cysteine in blood is catalyzed by iron and vitamin B6. Commun. Biol. 2019, 2, 194. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roglans, N.; Fauste, E.; Bentanachs, R.; Velázquez, A.M.; Pérez-Armas, M.; Donis, C.; Panadero, M.I.; Alegret, M.; Otero, P.; Bocos, C.; et al. Bempedoic Acid Restores Liver H2S Production in a Female Sprague-Dawley Rat Dietary Model of Non-Alcoholic Fatty Liver. Int. J. Mol. Sci. 2023, 24, 473. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010473
Roglans N, Fauste E, Bentanachs R, Velázquez AM, Pérez-Armas M, Donis C, Panadero MI, Alegret M, Otero P, Bocos C, et al. Bempedoic Acid Restores Liver H2S Production in a Female Sprague-Dawley Rat Dietary Model of Non-Alcoholic Fatty Liver. International Journal of Molecular Sciences. 2023; 24(1):473. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010473
Chicago/Turabian StyleRoglans, Núria, Elena Fauste, Roger Bentanachs, Ana M. Velázquez, Madelin Pérez-Armas, Cristina Donis, María I. Panadero, Marta Alegret, Paola Otero, Carlos Bocos, and et al. 2023. "Bempedoic Acid Restores Liver H2S Production in a Female Sprague-Dawley Rat Dietary Model of Non-Alcoholic Fatty Liver" International Journal of Molecular Sciences 24, no. 1: 473. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010473