

Study of Seed Ageing in lpa1-1 Maize Mutant and Two Possible Approaches to Restore Seed Germination

 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
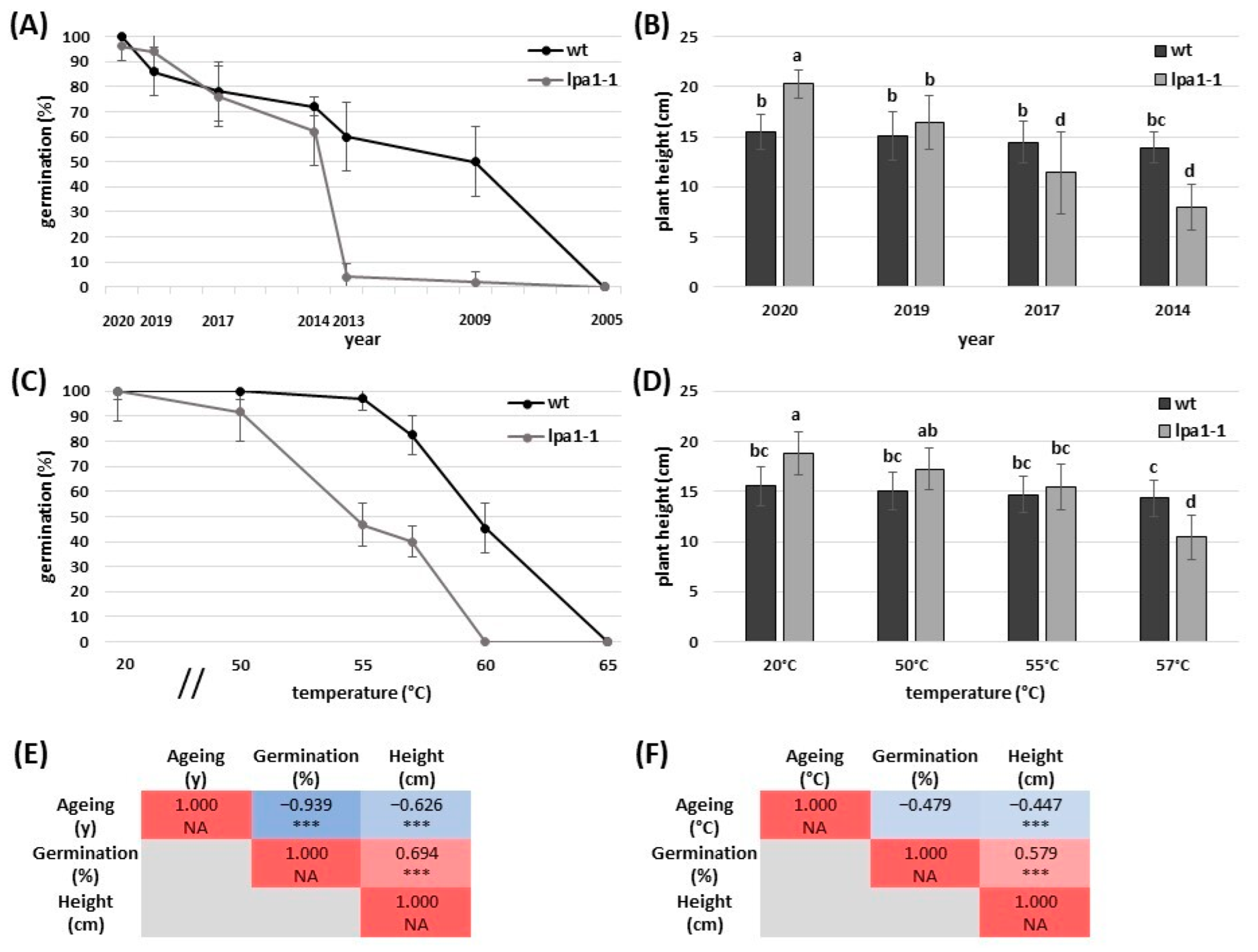
2.1. Accelerated Ageing Mimics the Natural Ageing
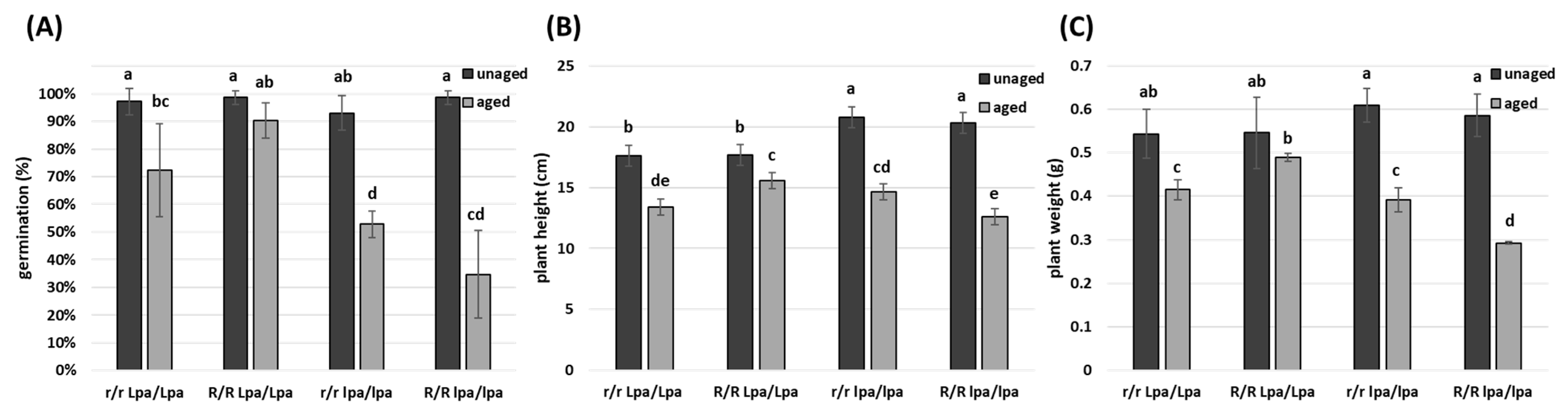
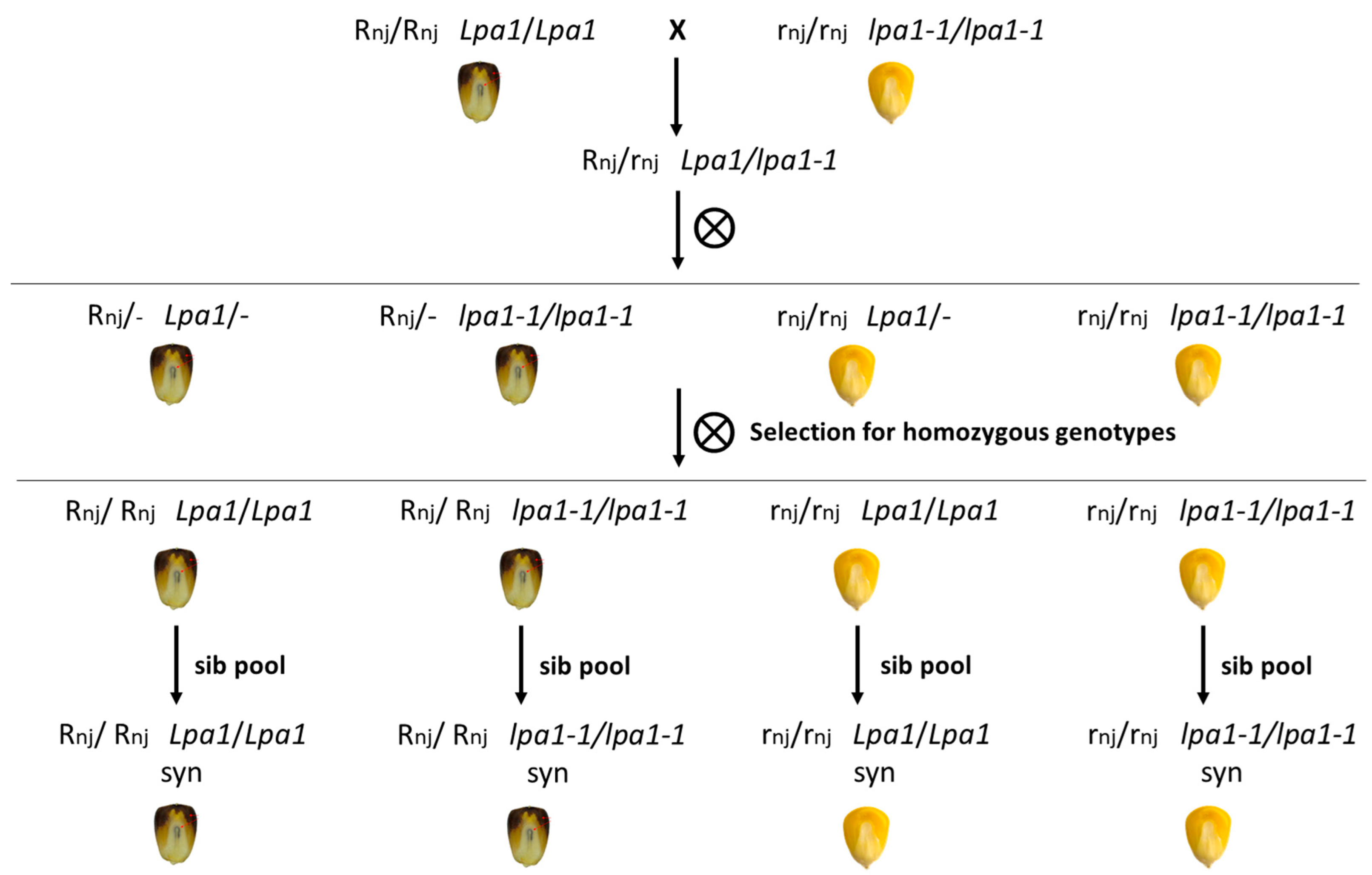
2.2. Genetic Approach to Counteract Seed Ageing
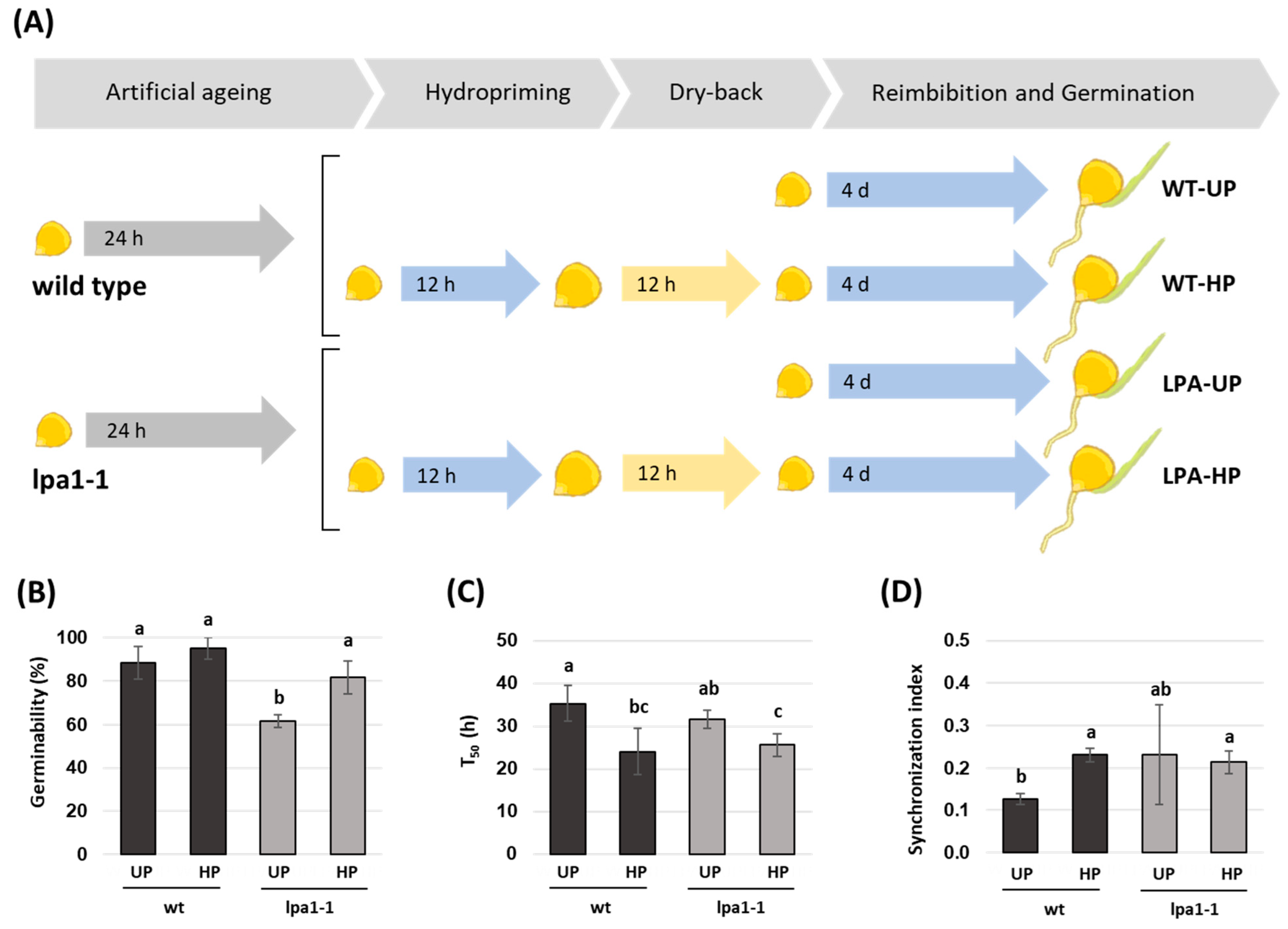
2.3. Hydropriming Improves Germination in Aged Seeds
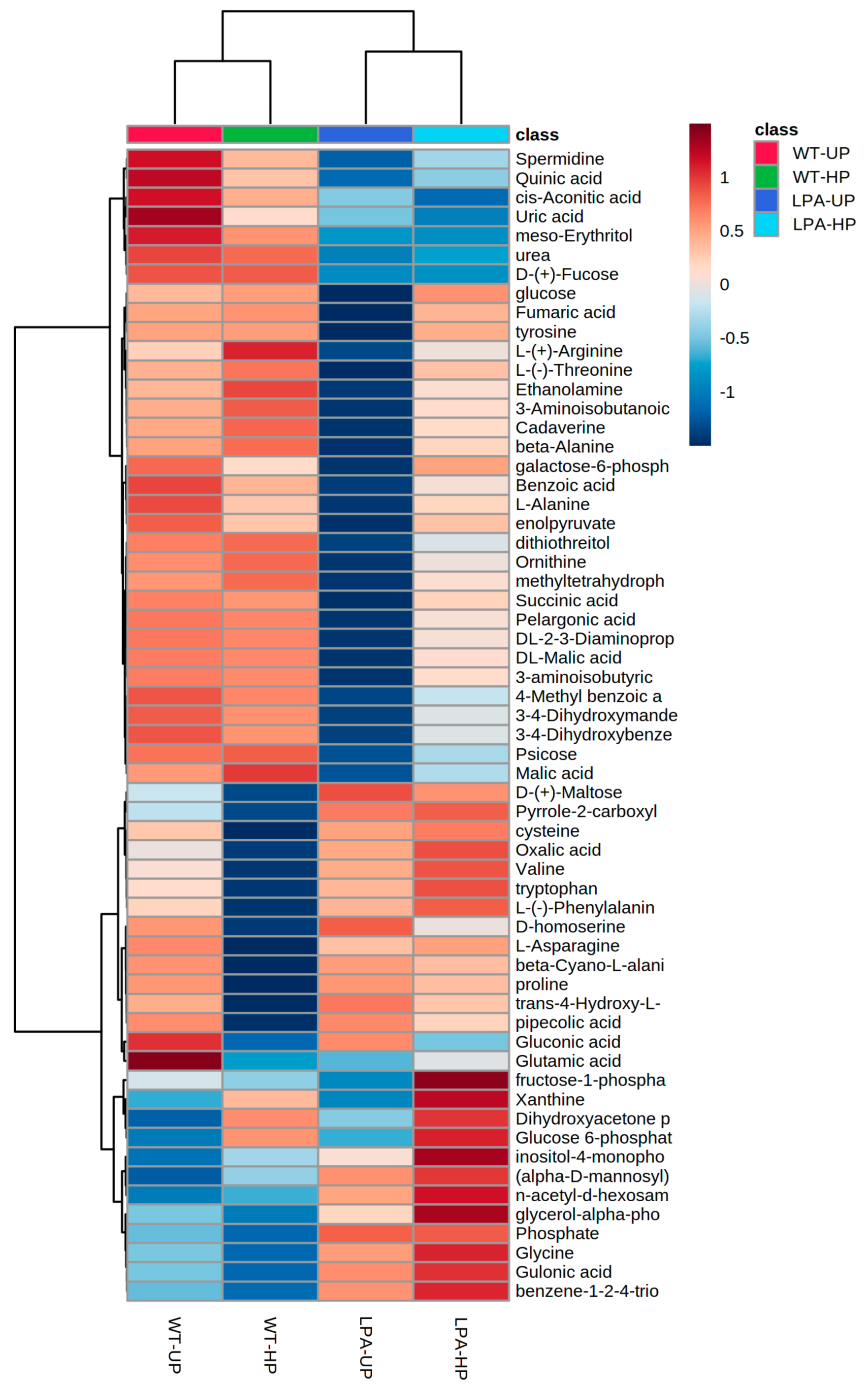
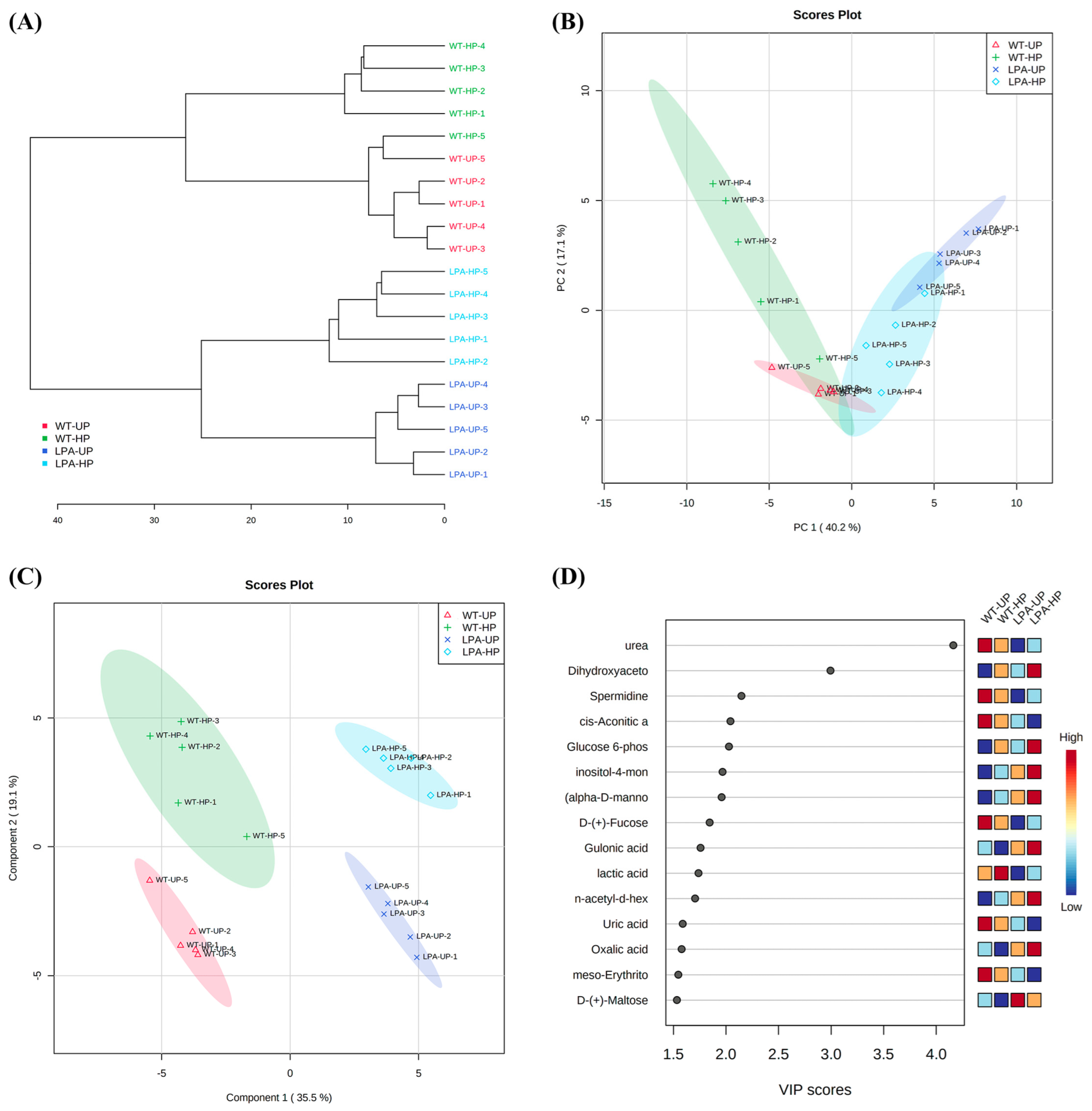
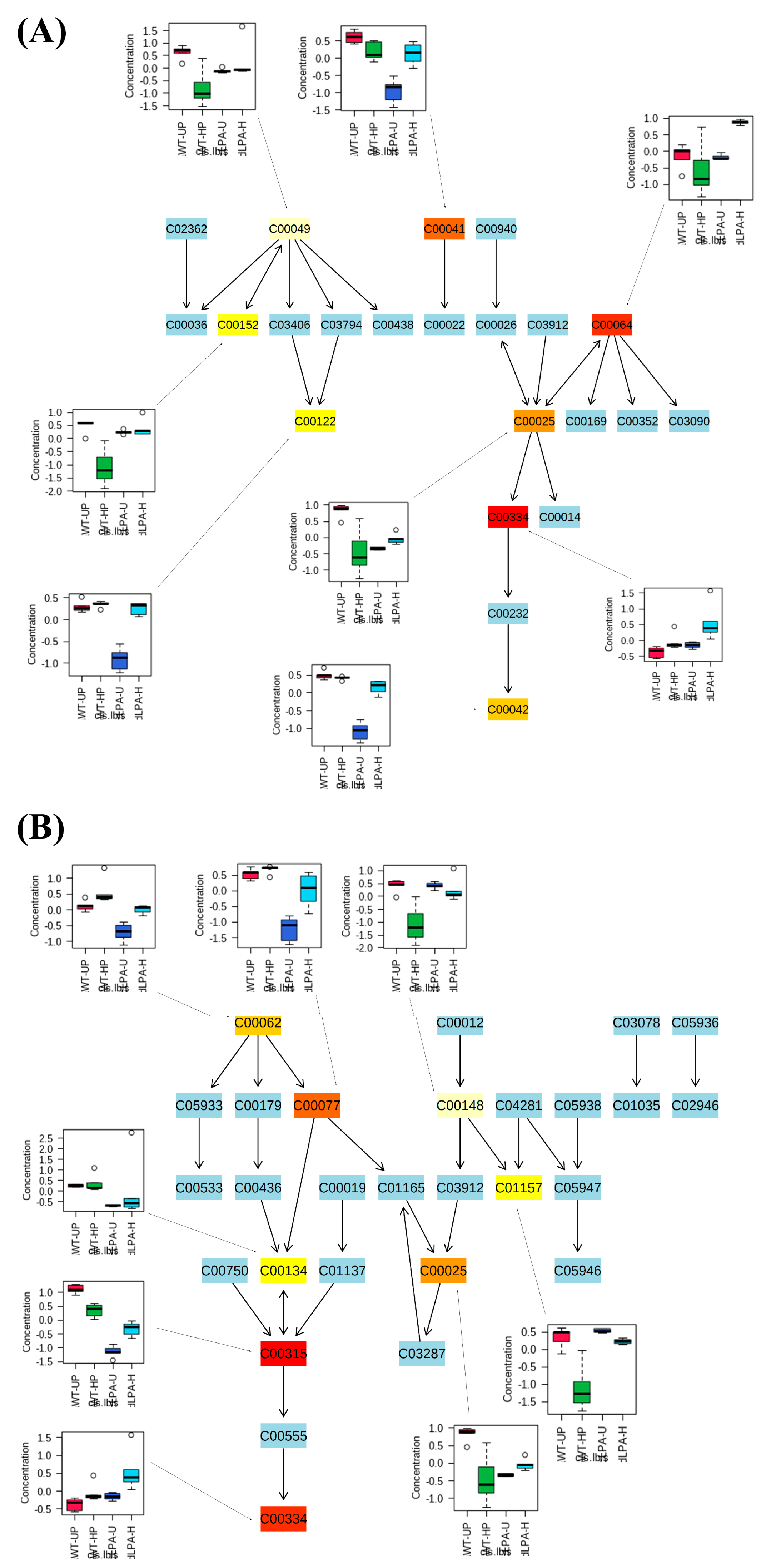
2.4. Untargeted Metabolomic Analysis
3. Discussion
4. Materials and Methods
4.1. Genetic Material
4.2. Ageing Conditions and Germination Tests
4.3. Constitution of the New Genetic Material and Plot Experiment
4.4. Hydropriming Protocol and Germination Test
4.5. Metabolomic Analysis
4.5.1. Samples’ Extraction and Derivatization
4.5.2. GC-MS Analysis
4.5.3. MS-DIAL Analysis
4.5.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raboy, V.; Young, K.A.; Dorsch, J.A.; Cook, A. Genetics and breeding of seed phosphorus and phytic acid. J. Plant Physiol. 2001, 158, 489–497. [Google Scholar] [CrossRef]
- Schlemmer, U.; Frølich, W.; Prieto, R.M.; Grases, F. Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis. Mol. Nutr. Food Res. 2009, 53, S330–S375. [Google Scholar] [CrossRef] [PubMed]
- Raboy, V. Accumulation and Storage of Phosphate and Minerals; Larkins, B.A., Asil, I.K., Eds.; Kluwer Academic: Amsterdam, The Netherlands, 1997. [Google Scholar]
- Raboy, V. Progress in Breeding Low Phytate Crops. Am. Soc. Nutr. Sci. 2002, 132, 503–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laboure, A.M.; Gagnon, J.; Lescure, A.M. Purification and characterization of a phytase (myo-inositol-hexakisphosphate phosphohydrolase) accumulated in maize (Zea mays) seedlings during germination. Biochem. J. 1993, 295, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Graf, E.; Mahoney, J.R.; Bryant, R.G.; Eaton, J.W. Iron-catalyzed hydroxyl radical formation. Stringent requirement for free iron coordination site. J. Biol. Chem. 1984, 259, 3620–3624. [Google Scholar] [CrossRef]
- Graf, E.; Empson, K.L.; Eaton, J.W. Phytic acid. A natural antioxidant. J. Biol. Chem. 1987, 262, 11647–11650. [Google Scholar] [CrossRef]
- Graf, E.; Eaton, J.W. Antioxidant functions of phytic acid. Free Radic. Biol. Med. 1990, 8, 61–69. [Google Scholar] [CrossRef]
- Raboy, V.; Gerbasi, P.F.; Young, K.A.; Stoneberg, S.D.; Pickett, S.G.; Bauman, A.T.; Murthy, P.P.N.; Sheridan, W.F.; Ertl, D.S. Origin and seed phenotype of maize low phytic acid 1-1 and low phytic acid 2-1. Plant Physiol. 2000, 124, 355–368. [Google Scholar] [CrossRef] [Green Version]
- Pilu, R.; Panzeri, D.; Gavazzi, G.; Rasmussen, S.K.; Consonni, G.; Nielsen, E. Phenotypic, genetic and molecular characterization of a maize low phytic acid mutant (lpa241). Theor. Appl. Genet. 2003, 107, 980–987. [Google Scholar] [CrossRef]
- Cerino Badone, F.; Amelotti, M.; Cassani, E.; Pilu, R. Study of low phytic acid1-7 (lpa1-7), a new ZmMRP4 mutation in maize. J. Hered. 2012, 103, 598–605. [Google Scholar] [CrossRef]
- Borlini, G.; Rovera, C.; Landoni, M.; Cassani, E.; Pilu, R. Lpa1-5525: A new lpa1 mutant isolated in a mutagenized population by a novel non-disrupting screening method. Plants 2019, 8, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, S.R.; Young, K.A.; Cook, A.; Blake, T.K.; Raboy, V. Linkage mapping of two mutations that reduce phytic acid content of barley grain. Theor. Appl. Genet. 1998, 97, 141–146. [Google Scholar] [CrossRef]
- Rasmussen, S.K.; Hatzack, F. Identification of two low-phytate barley (Hordeum vulgare L.) grain mutants by TLC and genetic analysis. Hereditas 1998, 129, 107–112. [Google Scholar] [CrossRef]
- Bregitzer, P.; Raboy, V. Effects of four independent low-phytate mutations on barley agronomic performance. Crop Sci. 2006, 46, 1318–1322. [Google Scholar] [CrossRef]
- Larson, S.R.; Rutger, J.N.; Young, K.A.; Raboy, V. Isolation and genetic mapping of a non-lethal rice (Oryza sativa L.) low phytic acid 1 mutation. Crop Sci. 2000, 40, 1397–1405. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.L.; Xu, X.H.; Ren, X.L.; Fu, H.W.; Wu, D.X.; Shu, Q.Y. Generation and characterization of low phytic acid germplasm in rice (Oryza sativa L.). Theor. Appl. Genet. 2007, 114, 803–814. [Google Scholar] [CrossRef]
- Guttieri, M.; Bowen, D.; Dorsch, J.A.; Raboy, V.; Souza, E. Identification and characterization of a low phytic acid wheat. Crop Sci. 2004, 44, 418–424. [Google Scholar] [CrossRef]
- Wilcox, J.R.; Premachandra, G.S.; Young, K.A.; Raboy, V. Isolation of high seed inorganic P, low-phytate soybean mutants. Crop Sci. 2000, 40, 1601–1605. [Google Scholar] [CrossRef] [Green Version]
- Hitz, W.D.; Carlson, T.J.; Kerr, P.S.; Sebastian, S.A. Biochemical and molecular characterization of a mutation that confers a decreased raffinosaccharide and phytic acid phenotype on soybean seeds. Plant Physiol. 2002, 128, 650–660. [Google Scholar] [CrossRef]
- Yuan, F.J.; Zhao, H.J.; Ren, X.L.; Zhu, S.L.; Fu, X.J.; Shu, Q.Y. Generation and characterization of two novel low phytate mutations in soybean (Glycine max L. Merr.). Theor. Appl. Genet. 2007, 115, 945–957. [Google Scholar] [CrossRef]
- Campion, B.; Sparvoli, F.; Doria, E.; Tagliabue, G.; Galasso, I.; Fileppi, M.; Bollini, R.; Nielsen, E. Isolation and characterisation of an lpa (low phytic acid) mutant in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2009, 118, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Cominelli, E.; Confalonieri, M.; Carlessi, M.; Cortinovis, G.; Daminati, M.G.; Porch, T.G.; Losa, A.; Sparvoli, F. Phytic acid transport in Phaseolus vulgaris: A new low phytic acid mutant in the PvMRP1 gene and study of the PvMRPs promoters in two different plant systems. Plant Sci. 2018, 270, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Colombo, F.; Bertagnon, G.; Ghidoli, M.; Pesenti, M.; Giupponi, L.; Pilu, R. Low-Phytate Grains to Enhance Phosphorus Sustainability in Agriculture: Chasing Drought Stress in lpa1-1 Mutant. Agronomy 2022, 12, 721. [Google Scholar] [CrossRef]
- Raboy, V. Seed Phosphate; Springer: New York, NY, USA, 2009. [Google Scholar]
- Shi, J.; Wang, H.; Schellin, K.; Li, B.; Faller, M.; Stoop, J.M.; Meeley, R.B.; Ertl, D.S.; Ranch, J.P.; Glassman, K. Embryo-specific silencing of a transporter reduces phytic acid content of maize and soybean seeds. Nat. Biotechnol. 2007, 25, 930–937. [Google Scholar] [CrossRef] [PubMed]
- Swarbreck, D.; Ripoll, P.J.; Brown, D.A.; Edwards, K.J.; Theodoulou, F. Isolation and characterisation of two multidrug resistance associated protein genes from maize. Gene 2003, 315, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.; Burla, B.; Martinoia, E. The multidrug resistance-associated protein (MRP/ABCC) subfamily of ATP-binding cassette transporters in plants. FEBS Lett. 2006, 580, 1112–1122. [Google Scholar] [CrossRef] [Green Version]
- Pilu, R.; Landoni, M.; Cassani, E.; Doria, E.; Nielsen, E. The maize lpa241 mutation causes a remarkable variability of expression and some pleiotropic effects. Crop Sci. 2005, 45, 2096–2105. [Google Scholar] [CrossRef]
- Pilu, R.; Panzeri, D.; Cassani, E.; Badone, F.C.; Landoni, M.; Nielsen, E. A paramutation phenomenon is involved in the genetics of maize low phytic acid1-241 (lpa1-241) trait. Heredity 2009, 102, 236–245. [Google Scholar] [CrossRef] [Green Version]
- Colombo, F.; Paolo, D.; Cominelli, E.; Sparvoli, F.; Nielsen, E.; Pilu, R. MRP Transporters and Low Phytic Acid Mutants in Major Crops: Main Pleiotropic Effects and Future Perspectives. Front. Plant Sci. 2020, 11, 1301. [Google Scholar] [CrossRef]
- Doria, E.; Galleschi, L.; Calucci, L.; Pinzino, C.; Pilu, R.; Cassani, E.; Nielsen, E. Phytic acid prevents oxidative stress in seeds: Evidence from a maize (Zea mays L.) low phytic acid mutant. J. Exp. Bot. 2009, 60, 967–978. [Google Scholar] [CrossRef]
- Groot, S.P.C.; Surki, A.A.; De Vos, R.C.H.; Kodde, J. Seed storage at elevated partial pressure of oxygen, a fast method for analysing seed ageing under dry conditions. Ann. Bot. 2012, 110, 1149–1159. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, F.; Li, X.; Peng, Y.; Zhang, H.; Hey, S.; Wang, G.; Wang, J.; Gu, R. Comparative analysis of the accelerated aged seed transcriptome profiles of two maize chromosome segment substitution lines. PLoS ONE 2019, 14, e0216977. [Google Scholar] [CrossRef] [PubMed]
- Debeaujon, I.; Léon-Kloosterziel, K.M.; Koornneef, M. Influence of the testa on seed dormancy, germination, and longevity in Arabidopsis. Plant Physiol. 2000, 122, 403–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murthy, U.M.N.; Kumar, P.P.; Sun, W.Q. Mechanisms of seed ageing under different storage conditions for Vigna radiata (L.) Wilczek: Lipid peroxidation, sugar hydrolysis, Maillard reactions and their relationship to glass state transition. J. Exp. Bot. 2003, 54, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Tian, Q.; Yin, G.; Chen, X.; Zhang, J.; Ng, S.; Lu, X. Reduced mitochondrial and ascorbate-glutathione activity after artificial ageing in soybean seed. J. Plant Physiol. 2014, 171, 140–147. [Google Scholar] [CrossRef]
- Delouche, J.C.; Baskin, C.C. Accelerated aging techniques for predicting the relative storability of seed lots. Proceedings 1973, 1, 427–452. [Google Scholar]
- TeKrony, D.J.; Ibrahim, A.E.; TeKrony, D.M.; Egli, D.B. Accelerated aging techniques for evaluating Sorghum seed vigor. J. Seed Technol. 1993, 17, 29–37. [Google Scholar]
- Woltz, J.M.; TeKrony, D.M. Accelerated Aging Test for Corn Seed. Seed Technol. 2001, 23, 21–34. [Google Scholar]
- Wattanakulpakin, P.; Photchanachai, S.; Miyagawa, S.; Ratanakhanokchai, K. Loss of maize seed vigor as affected by biochemical changes during hydropriming. Crop Sci. 2012, 52, 2783–2793. [Google Scholar] [CrossRef]
- Frank, T.; Seumo Meuleye, B.; Miller, A.; Shu, Q.Y.; Engel, K.H. Metabolite profiling of two low phytic acid (lpa) rice mutants. J. Agric. Food Chem. 2007, 55, 11011–11019. [Google Scholar] [CrossRef]
- Frank, T.; Nörenberg, S.; Engel, K.H. Metabolite profiling of two novel low phytic acid (lpa) soybean mutants. J. Agric. Food Chem. 2009, 57, 6408–6416. [Google Scholar] [CrossRef] [PubMed]
- Landoni, M.; Cerino Badone, F.; Haman, N.; Schiraldi, A.; Fessas, D.; Cesari, V.; Toschi, I.; Cremona, R.; Delogu, C.; Villa, D.; et al. Low phytic acid 1 mutation in maize modifies density, starch properties, cations, and fiber contents in the seed. J. Agric. Food Chem. 2013, 61, 4622–4630. [Google Scholar] [CrossRef] [PubMed]
- Colombo, F.; Sangiorgio, S.; Abruzzese, A.; Bononi, M.; Tateo, F.; Singh, S.K.; Nocito, F.F.; Pilu, R. The Potential of Low Phytic Acid1-1 Mutant in Maize (Zea mays L.): A Sustainable Solution to Non-Renewable Phosphorus. Front. Biosci. 2022, 27, 284. [Google Scholar] [CrossRef] [PubMed]
- Tekrony, D.M. Accelerated Aging Test: Principles and Procedures. Seed Technol. 2005, 27, 135–146. [Google Scholar]
- Agacka-Mołdoch, M.; Arif, M.A.R.; Lohwasser, U.; Doroszewska, T.; Qualset, C.O.; Börner, A. The inheritance of wheat grain longevity: A comparison between induced and natural ageing. J. Appl. Genet. 2016, 57, 477–481. [Google Scholar] [CrossRef]
- Petroni, K.; Pilu, R.; Tonelli, C. Anthocyanins in corn: A wealth of genes for human health. Planta 2014, 240, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Goodman, C.D.; Casati, P.; Walbot, V. A multidrug resistance-associated protein involved in anthocyanin transport in Zea mays. Plant Cell 2004, 16, 1812–1826. [Google Scholar] [CrossRef] [Green Version]
- Badone, F.C.; Cassani, E.; Landoni, M.; Doria, E.; Panzeri, D.; Lago, C.; Mesiti, F.; Nielsen, E.; Pilu, R. The low phytic acid1-241 (lpa1-241) maize mutation alters the accumulation of anthocyanin pigment in the kernel. Planta 2010, 231, 1189–1199. [Google Scholar] [CrossRef]
- Tilden, R.L.; West, S.H. Reversal of the effects of aging in soybean seeds. Plant Physiol. 1985, 77, 584–586. [Google Scholar] [CrossRef]
- Yan, H.; Jia, S.; Mao, P. Melatonin priming alleviates aging-induced germination inhibition by regulating β-oxidation, protein translation, and antioxidant metabolism in oat (Avena sativa L.) seeds. Int. J. Mol. Sci. 2020, 21, 1898. [Google Scholar] [CrossRef] [Green Version]
- Hazebroek, J.; Harp, T.; Shi, J.; Wang, H. Metabolomic Analysis of Low Phytic Acid Maize Kernels. In Concepts in Plant Metabolomics; Springer: Dordrecht, The Netherlands, 2007; pp. 221–238. [Google Scholar]
- Sharma, S.; Hari, L.; Anandkumar, D.; Tyagi, A.; Muthumilarasan, M. An insight into phytic acid biosynthesis and its reduction strategies to improve mineral bioavailability. Nucleus 2022, 65, 255–267. [Google Scholar] [CrossRef]
- Shen, W.; Wei, Y.; Dauk, M.; Zheng, Z.; Zou, J. Identification of a mitochondrial glycerol-3-phosphate dehydrogenase from Arabidopsis thaliana: Evidence for a mitochondrial glycerol-3-phosphate shuttle in plants. FEBS Lett. 2003, 536, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.; Rangarajan, P.; Donahue, J.L.; Williams, S.P.; Land, E.S.; Mandal, M.K.; Phillippy, B.Q.; Perera, I.Y.; Raboy, V.; Gillaspy, G.E. Two inositol hexakisphosphate kinases drive inositol pyrophosphate synthesis in plants. Plant J. 2014, 80, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Zanganeh, R.; Jamei, R.; Rahmani, F. Impacts of seed priming with salicylic acid and sodium hydrosulfide on possible metabolic pathway of two amino acids in maize plant under lead stress. Mol. Biol. Res. Commun. 2018, 7, 83–88. [Google Scholar] [CrossRef]
- Li, Y.; Liang, L.; Li, W.; Ashraf, U.; Ma, L.; Tang, X.; Pan, S.; Tian, H.; Mo, Z. ZnO nanoparticle-based seed priming modulates early growth and enhances physio-biochemical and metabolic profiles of fragrant rice against cadmium toxicity. J. Nanobiotechnol. 2021, 19, 75. [Google Scholar] [CrossRef]
- Zhou, X.; Jia, X.; Zhang, Z.; Chen, K.; Wang, L.; Chen, H.; Yang, Z.; Li, C.; Zhao, L. AgNPs seed priming accelerated germination speed and altered nutritional profile of Chinese cabbage. Sci. Total Environ. 2022, 808, 151896. [Google Scholar] [CrossRef]
- Ruan, Y.; Cai, Z.; Deng, Y.; Pan, D.; Zhou, C.; Cao, J.; Chen, X.; Xia, Q. An untargeted metabolomic insight into the high-pressure stress effect on the germination of wholegrain Oryza sativa L. Food Res. Int. 2021, 140, 109984. [Google Scholar] [CrossRef]
- Fait, A.; Angelovici, R.; Less, H.; Ohad, I.; Urbanczyk-Wochniak, E.; Fernie, A.R.; Galili, G. Arabidopsis seed development and germination is associated with temporally distinct metabolic switches. Plant Physiol. 2006, 142, 839–854. [Google Scholar] [CrossRef] [Green Version]
- Angelovici, R.; Fait, A.; Fernie, A.R.; Galili, G. A seed high-lysine trait is negatively associated with the TCA cycle and slows down Arabidopsis seed germination. New Phytol. 2011, 189, 148–159. [Google Scholar] [CrossRef]
- Gerdes, J.T.; Behr, C.F.; Coors, J.G.; Tracy, W.F. Compilation of North American Maize Breeding Germplasm; Crop Science Society of America: Madison, WI, USA, 1993; ISBN 9780891185758. [Google Scholar]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Ranal, M.A.; De Santana, D.G. How and why to measure the germination process? Rev. Bras. Bot. 2006, 29, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Assaad, H.I.; Hou, Y.; Zhou, L.; Carroll, R.J.; Wu, G. Rapid publication-ready MS-Word tables for two-way ANOVA. Springerplus 2015, 4, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, B.B.; Das, V.; Landi, M.; Abenavoli, M.R.; Araniti, F. Short-term effects of the allelochemical umbelliferone on Triticum durum L. metabolism through GC–MS based untargeted metabolomics. Plant Sci. 2020, 298, 110548. [Google Scholar] [CrossRef] [PubMed]
- Sansone, S.-A.; Fan, T.; Goodacre, R.; Griffin, J.L.; Hardy, N.W.; Kaddurah-Daouk, R.; Kristal, B.S.; Lindon, J.; Mendes, P.; Morrison, N.; et al. The metabolomics standards initiative (MSI). Nat. Biotechnol. 2007, 25, 864. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Total Cmpd | Hits | p-Value | FDR | Impact |
|---|---|---|---|---|---|
| Alanine aspartate and glutamate metabolism | 22 | 8 | 1.39 × 10−8 | 3.97 × 10−7 | 0.77698 |
| Arginine and proline metabolism | 28 | 8 | 4.00 × 10−7 | 2.07 × 10−6 | 0.63167 |
| Tryptophan metabolism | 23 | 2 | 1.58 × 10−5 | 3.45 × 10−5 | 0.5862 |
| Glycine serine and threonine metabolism | 33 | 6 | 7.72 × 10−9 | 3.97 × 10−7 | 0.57964 |
| Phenylalanine metabolism | 12 | 1 | 0.000234 | 0.000333 | 0.42308 |
| Starch and sucrose metabolism | 22 | 5 | 2.41 × 10−7 | 1.53 × 10−6 | 0.4125 |
| Isoquinoline alkaloid biosynthesis | 6 | 2 | 0.007983 | 0.009286 | 0.41176 |
| beta-Alanine metabolism | 18 | 6 | 1.01 × 10−6 | 4.80 × 10−6 | 0.4008 |
| Cyanoamino acid metabolism | 26 | 6 | 2.82 × 10−6 | 8.91 × 10−6 | 0.375 |
| Arginine biosynthesis | 18 | 7 | 1.87 × 10−6 | 6.97 × 10−6 | 0.34079 |
| Tyrosine metabolism | 18 | 3 | 1.65 × 10−7 | 1.42 × 10−6 | 0.27568 |
| Pantothenate and CoA biosynthesis | 23 | 5 | 1.92 × 10−5 | 4.05 × 10−5 | 0.24406 |
| Citrate cycle (TCA cycle) | 20 | 5 | 1.36 × 10−6 | 5.55 × 10−6 | 0.23269 |
| Glyoxylate and dicarboxylate metabolism | 29 | 8 | 9.20 × 10−8 | 1.42 × 10−6 | 0.20626 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Colombo, F.; Pagano, A.; Sangiorgio, S.; Macovei, A.; Balestrazzi, A.; Araniti, F.; Pilu, R. Study of Seed Ageing in lpa1-1 Maize Mutant and Two Possible Approaches to Restore Seed Germination. Int. J. Mol. Sci. 2023, 24, 732. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010732
Colombo F, Pagano A, Sangiorgio S, Macovei A, Balestrazzi A, Araniti F, Pilu R. Study of Seed Ageing in lpa1-1 Maize Mutant and Two Possible Approaches to Restore Seed Germination. International Journal of Molecular Sciences. 2023; 24(1):732. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010732
Chicago/Turabian StyleColombo, Federico, Andrea Pagano, Stefano Sangiorgio, Anca Macovei, Alma Balestrazzi, Fabrizio Araniti, and Roberto Pilu. 2023. "Study of Seed Ageing in lpa1-1 Maize Mutant and Two Possible Approaches to Restore Seed Germination" International Journal of Molecular Sciences 24, no. 1: 732. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010732