
Characterization of the Rate-Limiting Steps in the Dark-To-Light Transitions of Closed Photosystem II: Temperature Dependence and Invariance of Waiting Times during Multiple Light Reactions
 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Temperature Dependence of the Variable Chl-a Fluorescence (Fv) Induced by STSFs
2.2. Temperature Dependence of Δτ1/2
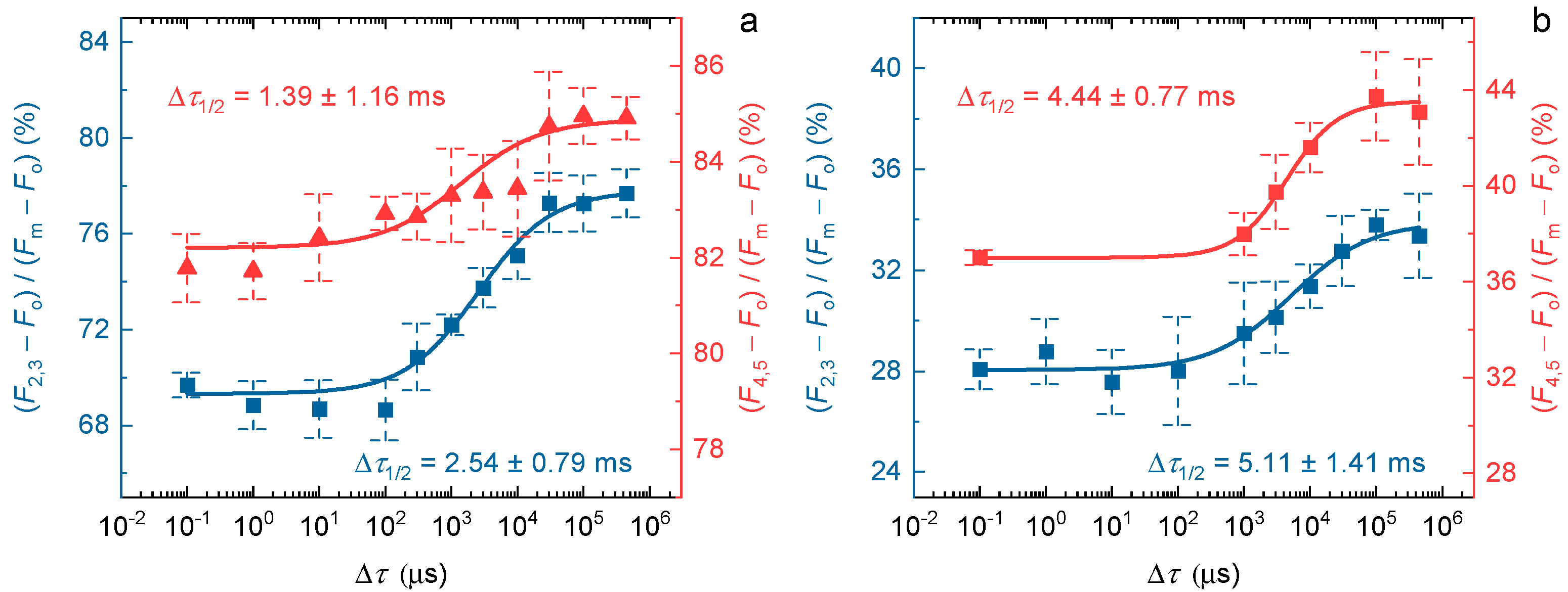
2.3. Temperature Dependence of Δτ1/2 during Multiple Light Reactions
3. Materials and Methods
3.1. Growth Conditions
3.2. Sample Preparation
3.3. Chl-a Relative Fluorescence Yield Measurements
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Magyar, M.; Sipka, G.; Kovacs, L.; Ughy, B.; Zhu, Q.J.; Han, G.Y.; Spunda, V.; Lambrev, P.H.; Shen, J.R.; Garab, G. Rate-limiting steps in the dark-to-light transition of Photosystem II—Revealed by chlorophyll-a fluorescence induction. Sci. Rep. 2018, 8, 2755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sipka, G.; Magyar, M.; Mezzetti, A.; Akhtar, P.; Zhu, Q.; Xiao, Y.; Han, G.; Santabarbara, S.; Shen, J.-R.; Lambrev, P.H.; et al. Light-Adapted Charge-Separated State of Photosystem II: Structural and Functional Dynamics of the Closed Reaction Center. Plant Cell 2021, 33, 1286–1302. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.; Yocum, C.F. Structure and function of photosystems I and II. Annu. Rev. Plant Biol. 2006, 57, 521–565. [Google Scholar] [CrossRef] [Green Version]
- Romero, E.; Novoderezhkin, V.I.; van Grondelle, R. Quantum design of photosynthesis for bio-inspired solar-energy conversion. Nature 2017, 543, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis; Wiley: New York, NY, USA, 2021. [Google Scholar]
- Shen, J.R. The Structure of Photosystem II and the Mechanism of Water Oxidation in Photosynthesis. Annu. Rev. Plant Biol. 2015, 66, 23–48. [Google Scholar] [CrossRef] [Green Version]
- Umena, Y.; Kawakami, K.; Shen, J.R.; Kamiya, N. Crystal structure of oxygen-evolving photosystem II at a resolution of 1.9 angstrom. Nature 2011, 473, 55–60. [Google Scholar] [CrossRef]
- Cardona, T.; Sedoud, A.; Cox, N.; Rutherford, A.W. Charge separation in Photosystem II: A comparative and evolutionary overview. BBA-Bioenerg. 2012, 1817, 26–43. [Google Scholar] [CrossRef] [Green Version]
- Shlyk-Kerner, O.; Samish, I.; Kaftan, D.; Holland, N.; Sai, P.S.M.; Kless, H.; Scherz, A. Protein flexibility acclimatizes photosynthetic energy conversion to the ambient temperature. Nature 2006, 442, 827–830. [Google Scholar] [CrossRef]
- Lubitz, W.; Chrysina, M.; Cox, N. Water oxidation in photosystem II. Photosynth. Res. 2019, 142, 105–125. [Google Scholar] [CrossRef] [Green Version]
- Hasan, S.S.; Cramer, W.A. On rate limitations of electron transfer in the photosynthetic cytochrome b6f complex. Phys. Chem. Chem. Phys. 2012, 14, 13853–13860. [Google Scholar] [CrossRef]
- Holland, S.C.; Kappell, A.D.; Burnap, R.L. Redox changes accompanying inorganic carbon limitation in Synechocystis sp PCC 6803. BBA-Bioenerg. 2015, 1847, 355–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavrel, T.; Szabo, M.; Tamburic, B.; Evenhuis, C.; Kuzhiumparambil, U.; Literakova, P.; Larkum, A.W.D.; Raven, J.A.; Cerveny, J.; Ralph, P.J. Effect of carbon limitation on photosynthetic electron transport in Nannochloropsis oculata. J. Photoch. Photobio. B 2018, 181, 31–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suga, M.; Akita, F.; Sugahara, M.; Kubo, M.; Nakajima, Y.; Nakane, T.; Yamashita, K.; Umena, Y.; Nakabayashi, M.; Yamane, T.; et al. Light-induced structural changes and the site of O=O bond formation in PSII caught by XFEL. Nature 2017, 543, 131–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suga, M.; Akita, F.; Yamashita, K.; Nakajima, Y.; Ueno, G.; Li, H.; Yamane, T.; Hirata, K.; Umena, Y.; Yonekura, S.; et al. An oxyl/oxo mechanism for oxygen-oxygen coupling in PSII revealed by an x-ray free-electron laser. Science 2019, 366, 334–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, J.; Chatterjee, R.; Young, I.D.; Fuller, F.D.; Lassalle, L.; Ibrahim, M.; Gul, S.; Fransson, T.; Brewster, A.S.; Alonso-Mori, R.; et al. Structures of the intermediates of Kok’s photosynthetic water oxidation clock. Nature 2018, 563, 421–425. [Google Scholar] [CrossRef]
- Stowell, M.H.; McPhillips, T.M.; Rees, D.C.; Soltis, S.M.; Abresch, E.; Feher, G. Light-induced structural changes in photosynthetic reaction center: Implications for mechanism of electron-proton transfer. Science 1997, 276, 812–816. [Google Scholar] [CrossRef] [Green Version]
- Sugo, Y.; Saito, K.; Ishikita, H. Mechanism of the formation of proton transfer pathways in photosynthetic reaction centers. Proc. Natl. Acad. Sci. USA 2021, 118, e2103203118. [Google Scholar] [CrossRef]
- Wei, R.J.; Zhang, Y.; Mao, J.; Kaur, D.; Khaniya, U.; Gunner, M.R. Comparison of proton transfer paths to the Q(A) and Q(B) sites of the Rb. sphaeroides photosynthetic reaction centers. Photosynth. Res. 2022, 152, 153–165. [Google Scholar] [CrossRef]
- Michel, H.; Deisenhofer, J. Relevance of the Photosynthetic Reaction Center from Purple Bacteria to the Structure of Photosystem-II. Biochemistry 1988, 27, 1–7. [Google Scholar] [CrossRef]
- Heathcote, P.; Fyfe, P.K.; Jones, M.R. Reaction centres: The structure and evolution of biological solar power. Trends Biochem. Sci. 2002, 27, 79–87. [Google Scholar] [CrossRef]
- Krammer, E.M.; Sebban, P.; Ullmann, G.M. Profile Hidden Markov Models for Analyzing Similarities and Dissimilarities in the Bacterial Reaction Center and Photosystem II. Biochemistry 2009, 48, 1230–1243. [Google Scholar] [CrossRef] [PubMed]
- Cardona, T. Photosystem II is a Chimera of Reaction Centers. J. Mol. Evol. 2017, 84, 149–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Mourik, F.; Frese, R.N.; van der Zwan, G.; Cogdell, R.J.; van Grondelle, R. Direct observation of solvation dynamics and dielectric relaxation in the photosynthetic light-harvesting-2 complex of Rhodopseudomonas acidophila. J. Phys. Chem. B 2003, 107, 2156–2161. [Google Scholar] [CrossRef]
- Deshmukh, S.S.; Tang, K.; Kalman, L. Lipid binding to the carotenoid binding site in photosynthetic reaction centers. J. Am. Chem. Soc. 2011, 133, 16309–16316. [Google Scholar] [CrossRef]
- Deshmukh, S.S.; Williams, J.C.; Allen, J.P.; Kalman, L. Light-induced conformational changes in photosynthetic reaction centers: Dielectric relaxation in the vicinity of the dimer. Biochemistry 2011, 50, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Kleinfeld, D.; Okamura, M.Y.; Feher, G. Electron-Transfer Kinetics in Photosynthetic Reaction Centers Cooled to Cryogenic Temperatures in the Charge-Separated State—Evidence for Light-Induced Structural-Changes. Biochemistry 1984, 23, 5780–5786. [Google Scholar] [CrossRef] [PubMed]
- Abgaryan, G.A.; Christophorov, L.N.; Goushcha, A.O.; Holzwarth, A.R.; Kharkyanen, V.N.; Knox, P.P.; Lukashev, E.A. Effects of mutual influence of photoinduced electron transitions and slow structural rearrangements in bacterial photosynthetic reaction centers. J. Biol. Phys. 1998, 24, 1–17. [Google Scholar] [CrossRef]
- Goushcha, A.O.; Kharkyanen, V.N.; Scott, G.W.; Holzwarth, A.R. Self-regulation phenomena in bacterial reaction centers. I. General theory. Biophys. J. 2000, 79, 1237–1252. [Google Scholar] [CrossRef] [Green Version]
- Barabash, Y.M.; Berezetskaya, N.M.; Christophorov, L.N.; Goushcha, A.O.; Kharkyanen, V.N. Effects of structural memory in protein reactions. J. Chem. Phys. 2002, 116, 4339–4352. [Google Scholar] [CrossRef]
- Christophorov, L.; Holzwarth, A.; Kharkyanen, V. Conformational regulation in single molecule reactions. Ukr. J. Phys. 2003, 48, 672–680. [Google Scholar]
- Goushcha, A.O.; Manzo, A.J.; Scott, G.W.; Christophorov, L.N.; Knox, P.P.; Barabash, Y.M.; Kapoustina, M.T.; Berezetska, N.M.; Kharkyanen, V.N. Self-regulation phenomena applied to bacterial reaction centers 2. Nonequilibrium adiabatic potential: Dark and light conformations revisited. Biophys. J. 2003, 84, 1146–1160. [Google Scholar] [CrossRef] [PubMed]
- Joliot, P.; Joliot, A. Comparative-Study of the Fluorescence Yield and of the C550 Absorption Change at Room-Temperature. Biochim. Biophys. Acta 1979, 546, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Laisk, A.; Oja, V. Variable fluorescence of closed photochemical reaction centers. Photosynth. Res. 2020, 143, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Oja, V.; Laisk, A. Time- and reduction-dependent rise of photosystem II fluorescence during microseconds-long inductions in leaves. Photosynth. Res. 2020, 145, 209–225. [Google Scholar] [CrossRef] [PubMed]
- Sipka, G.; Muller, P.; Brettel, K.; Magyar, M.; Kovacs, L.; Zhu, Q.J.; Xiao, Y.A.; Han, G.Y.; Lambrev, P.H.; Shen, J.R.; et al. Redox transients of P680 associated with the incremental chlorophyll-a fluorescence yield rises elicited by a series of saturating flashes in diuron-treated photosystem II core complex of Thermosynechococcus vulcanus. Physiol. Plant. 2019, 166, 22–32. [Google Scholar] [CrossRef]
- Andreasson, U.; Andreasson, L.E. Characterization of a semi-stable, charge-separated state in reaction centers from Rhodobacter sphaeroides. Photosynth. Res. 2003, 75, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Malferrari, M.; Mezzetti, A.; Francia, F.; Venturoli, G. Effects of dehydration on light-induced conformational changes in bacterial photosynthetic reaction centers probed by optical and differential FTIR spectroscopy. BBA-Bioenerg. 2013, 1827, 328–339. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Govindjee. Polyphasic Chlorophyll-a Fluorescence Transient in Plants and Cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Govindjee, G.; Papageorgiou, G. Chlorophyll A Fluorescence: A Signature of Photosynthesis; Springer: Dordrecht, The Netherlands, 2004; pp. 1–41. [Google Scholar]
- Sipka, G.; Nagy, L.; Magyar, M.; Akhtar, P.; Shen, J.-R.; Holzwarth, A.R.; Lambrev, P.H.; Garab, G. Light-induced reversible reorganizations in closed Type-II reaction centre complexes. Physiological roles and physical mechanisms. Open Biol. 2022, 12, 220297. [Google Scholar] [CrossRef]
- Magyar, M.; Akhtar, P.; Sipka, G.; Han, W.; Li, X.; Han, G.; Shen, J.R.; Lambrev, P.H.; Garab, G. Dependence of the rate-limiting steps in the dark-to-light transition of photosystem II on the lipidic environment of the reaction center. Photosynthetica 2022, 60, 147–156. [Google Scholar] [CrossRef]
- Jaenicke, R.; Bohm, G. The stability of proteins in extreme environments. Curr. Opin. Struct. Biol. 1998, 8, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Garbers, A.; Reifarth, F.; Kurreck, J.; Renger, G.; Parak, F. Correlation between protein flexibility and electron transfer from QA to QB in PSII membrane fragments from spinach. Biochemistry 1998, 37, 11399–11404. [Google Scholar] [CrossRef] [PubMed]
- Pieper, J.; Hauss, T.; Buchsteiner, A.; Baczynski, K.; Adamiak, K.; Lechner, R.E.; Renger, G. Temperature- and hydration-dependent protein dynamics in photosystem II of green plants studied by quasielastic neutron scattering. Biochemistry 2007, 46, 11398–11409. [Google Scholar] [CrossRef]
- Pieper, J.; Trapp, M.; Skomorokhov, A.; Natkaniec, I.; Peters, J.; Renger, G. Temperature-dependent vibrational and conformational dynamics of photosystem II membrane fragments from spinach investigated by elastic and inelastic neutron scattering. BBA-Bioenerg. 2012, 1817, 1213–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, M.; Sokolov, A.P. Dielectric Spectroscopy of Hydrated Biomacromolecules. In Dielectric Relaxation in Biological Systems: Physical Principles, Methods, and Applications; Raicu, V., Feldman, Y., Eds.; Oxford University Press: Oxford, UK, 2015; pp. 248–275. [Google Scholar]
- Katona, G.; Snijder, A.; Gourdon, P.; Andreasson, U.; Hansson, O.; Andreasson, L.E.; Neutze, R. Conformational regulation of charge recombination reactions in a photosynthetic bacterial reaction center. Nat. Struct. Mol. Biol. 2005, 12, 630–631. [Google Scholar] [CrossRef] [PubMed]
- Iwata, T.; Paddock, M.L.; Okamura, M.Y.; Kandori, H. Identification of FTIR Bands Due to Internal Water Molecules around the Quinone Binding Sites in the Reaction Center from Rhodobacter sphaeroides. Biochemistry 2009, 48, 1220–1229. [Google Scholar] [CrossRef] [Green Version]
- Malferrari, M.; Turina, P.; Francia, F.; Mezzetti, A.; Leibl, W.; Venturoli, G. Dehydration affects the electronic structure of the primary electron donor in bacterial photosynthetic reaction centers: Evidence from visible-NIR and light-induced difference FTIR spectroscopy. Photoch. Photobio. Sci. 2015, 14, 238–251. [Google Scholar] [CrossRef] [Green Version]
- Koike, H.; Inoue, Y. Preparation of oxygen-evolving photosystem II particles from a thermophilic blue-green alga. In The Oxygen Evolving System of Photosynthesis; Inoue, Y., Crofts, A.R., Govindjee, M.N., Renger, G., Satoh, K., Eds.; Academic Press: Cambridge, MA, USA, 1983; pp. 257–263. [Google Scholar]
- Shen, J.R.; Kawakami, K.; Koike, H. Purification and crystallization of oxygen-evolving photosystem II core complex from thermophilic cyanobacteria. Methods Mol. Biol. 2011, 684, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Chylla, R.A.; Garab, G.; Whitmarsh, J. Evidence for Slow Turnover in a Fraction of Photosystem-II Complexes in Thylakoid Membranes. Biochim. Biophys. Acta 1987, 894, 562–571. [Google Scholar] [CrossRef]
- Shen, J.R.; Inoue, Y. Binding and functional properties of two new extrinsic components, cytochrome c-550 and a 12-kDa protein, in cyanobacterial photosystem II. Biochemistry 1993, 32, 1825–1832. [Google Scholar] [CrossRef]
- Shen, J.R.; Kamiya, N. Crystallization and the crystal properties of the oxygen-evolving photosystem II from Synechococcus vulcanus. Biochemistry 2000, 39, 14739–14744. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Shen, J.R. Purification of fully active and crystallizable photosystem II from thermophilic cyanobacteria. Methods Enzymol. 2018, 613, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, U.; Krieger, A. Two fundamentally different types of variable chlorophyll fluorescence in vivo. FEBS Lett. 1996, 397, 131–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, J.P.; Chamberlain, K.D.; Williams, J.C. Identification of amino acid residues in a proton release pathway near the bacteriochlorophyll dimer in reaction centers from Rhodobacter sphaeroides. Photosynth. Res. 2022, 1–12. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | PSII CC Δτ1/2 (ms) | Logistic Fit P | Fv/Fm | Thylakoid Δτ1/2 (ms) | Logistic Fit P | Fv/Fm |
|---|---|---|---|---|---|---|
| 23 | 1.16 ± 0.40 | 0.73 * | 0.80 ± 0.01 | - | - | - |
| 5 | 1.77 ± 0.39 | 0.73 * | 0.85 ± 0.02 | 0.20 ± 0.04 | 0.73 * | 0.65 ± 0.00 |
| −20 | 1.37 ± 0.49 | 0.73 * | 0.83 ± 0.01 | 0.42 ± 0.12 | 0.73 * | 0.60 ± 0.01 |
| −40 | 2.01 ± 0.45 | 0.73 * | 0.83 ± 0.01 | 0.36 ± 0.09 | 0.73 * | 0.56 ± 0.01 |
| −60 | 3.29 ± 0.93 | 0.73 * | 0.83 ± 0.00 | 0.80 ± 0.09 | 1.3 | 0.53 ± 0.02 |
| −80 | 4.18 ± 0.89 | 1.15 | 0.83 ± 0.01 | 1.14 ± 0.18 | 1.03 | 0.50 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magyar, M.; Sipka, G.; Han, W.; Li, X.; Han, G.; Shen, J.-R.; Lambrev, P.H.; Garab, G. Characterization of the Rate-Limiting Steps in the Dark-To-Light Transitions of Closed Photosystem II: Temperature Dependence and Invariance of Waiting Times during Multiple Light Reactions. Int. J. Mol. Sci. 2023, 24, 94. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010094
Magyar M, Sipka G, Han W, Li X, Han G, Shen J-R, Lambrev PH, Garab G. Characterization of the Rate-Limiting Steps in the Dark-To-Light Transitions of Closed Photosystem II: Temperature Dependence and Invariance of Waiting Times during Multiple Light Reactions. International Journal of Molecular Sciences. 2023; 24(1):94. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010094
Chicago/Turabian StyleMagyar, Melinda, Gábor Sipka, Wenhui Han, Xingyue Li, Guangye Han, Jian-Ren Shen, Petar H. Lambrev, and Győző Garab. 2023. "Characterization of the Rate-Limiting Steps in the Dark-To-Light Transitions of Closed Photosystem II: Temperature Dependence and Invariance of Waiting Times during Multiple Light Reactions" International Journal of Molecular Sciences 24, no. 1: 94. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010094