
Morphological Diversity of Dps Complex with Genomic DNA
 , ,
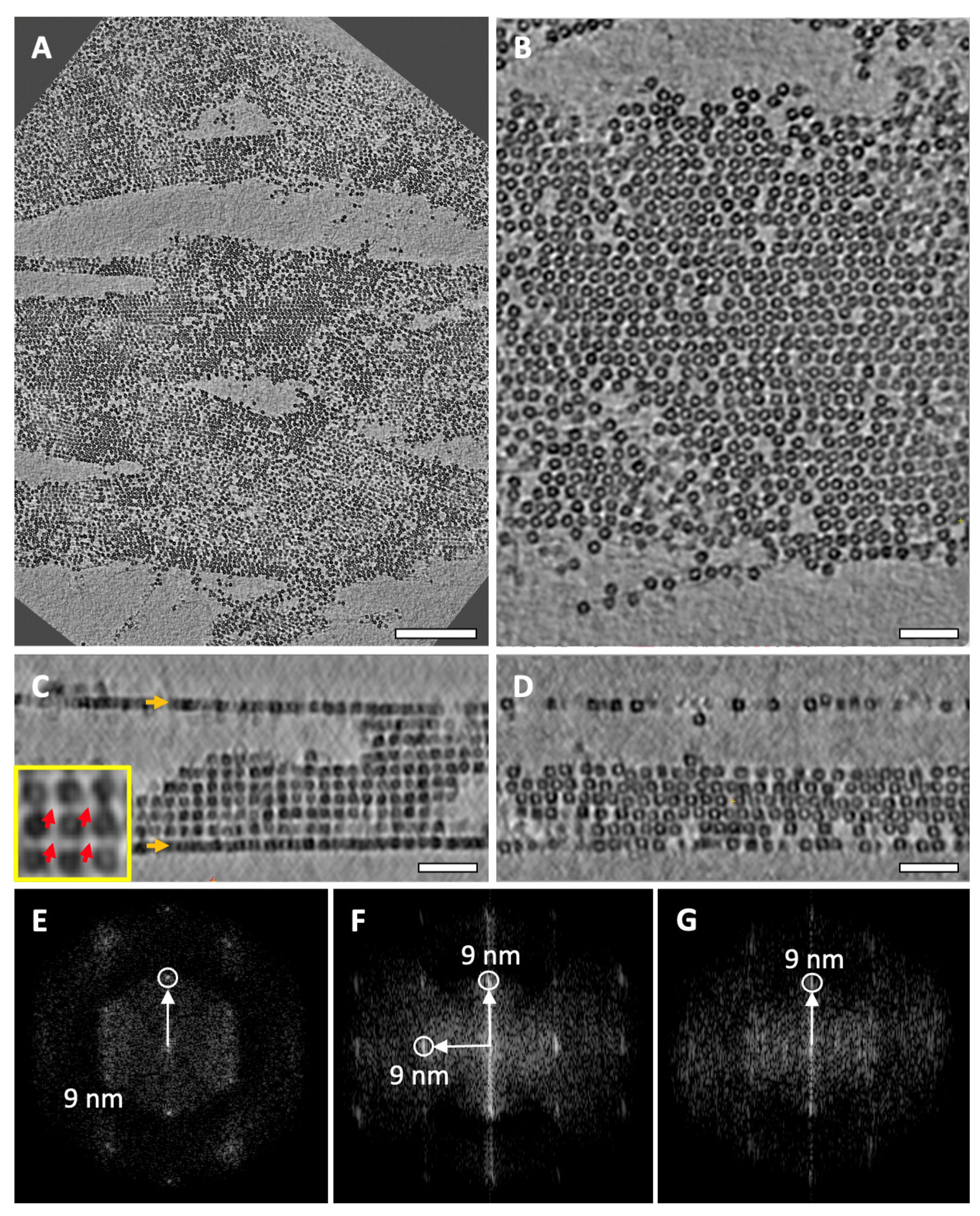
, ,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Conditions for the Formation of Dps Complexes with Genomic DNA
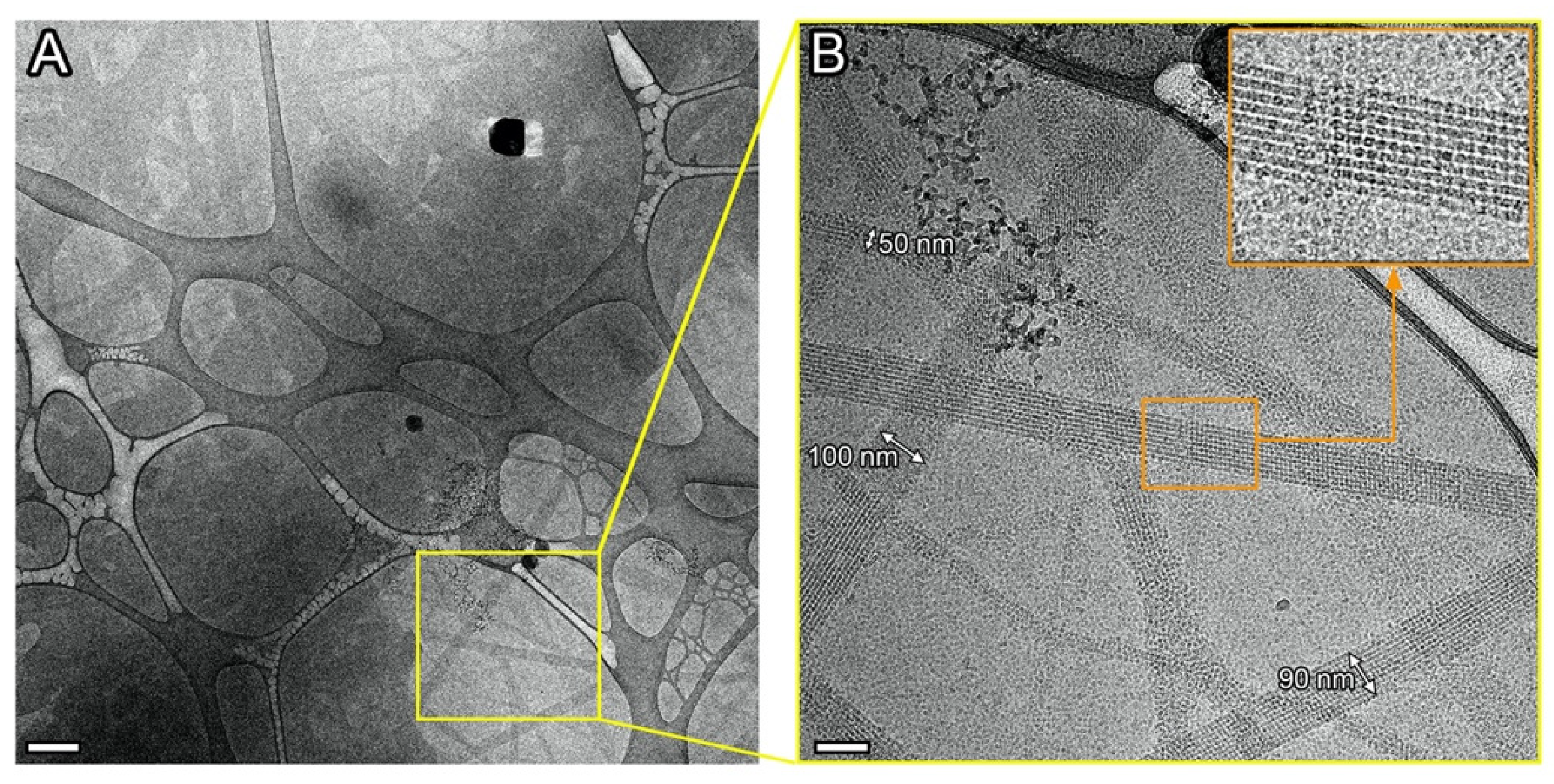
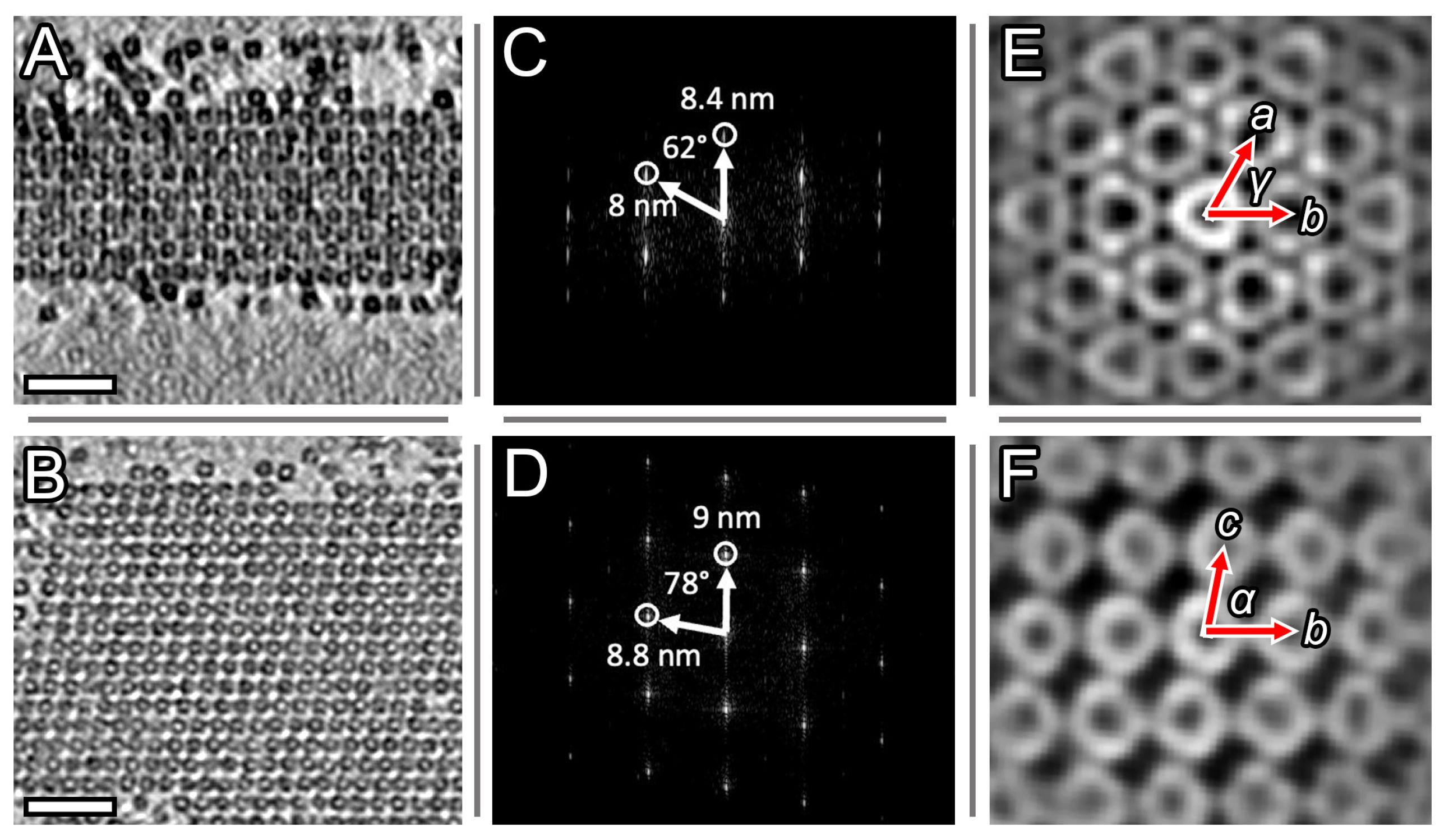
2.2. Elongated Dps–DNA Biocrystals, Filament-like Assemblies and Their Basic Structural Element
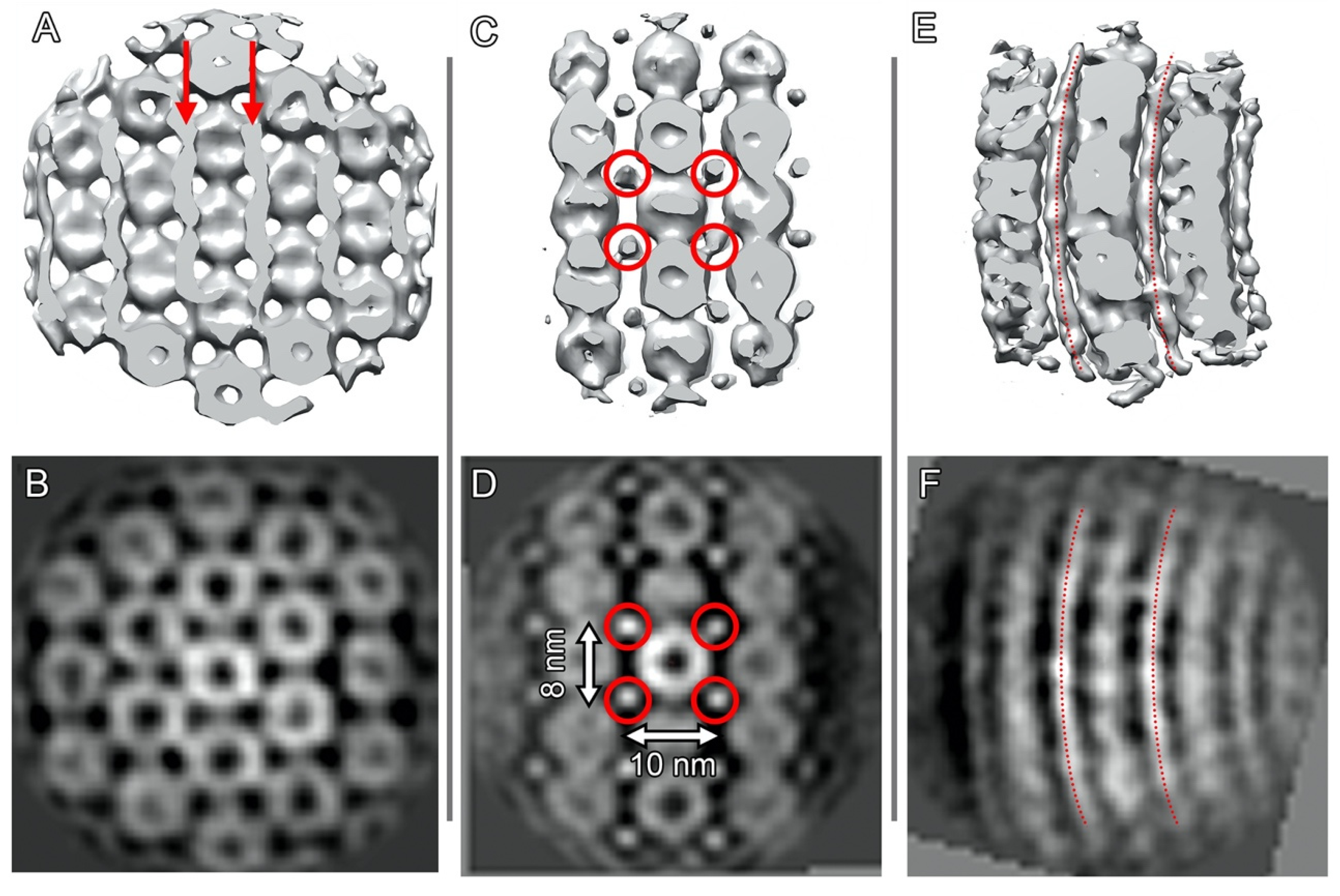
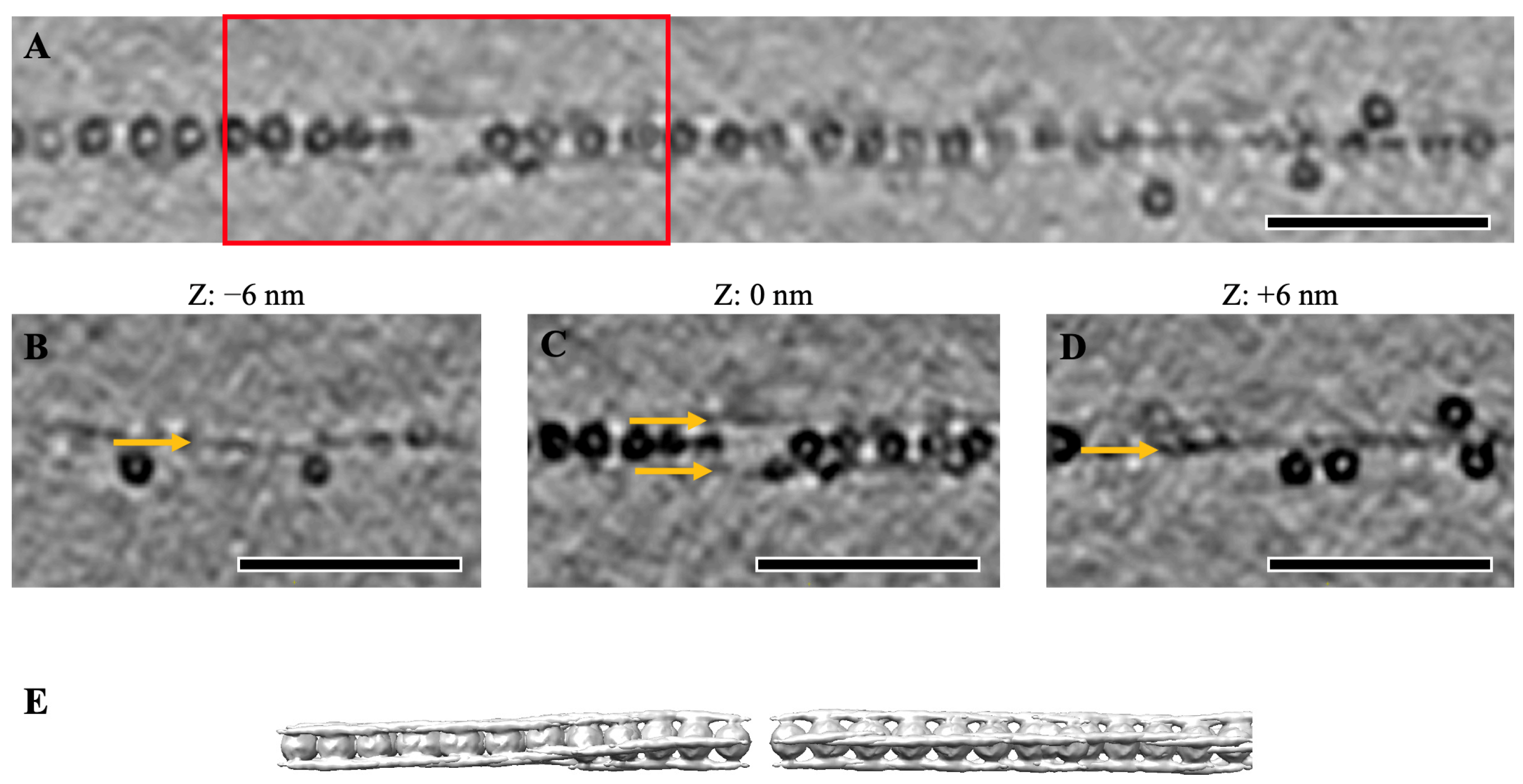
2.3. Cylindrical Assemblies
3. Discussion
4. Materials and Methods
4.1. Dps and DNA Isolation and Purification
4.2. Preparation for the Cryo-EM Study
4.3. Cryo-Electron Tomography Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A




References
- Luijsterburg, M.S.; Noom, M.C.; Wuite, G.J.L.; Dame, R.T. The Architectural Role of Nucleoid-Associated Proteins in the Organization of Bacterial Chromatin: A Molecular Perspective. J. Struct. Biol. 2006, 156, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, G.-W.; Chen, C.; Xie, X.S.; Zhuang, X. Chromosome Organization by a Nucleoid-Associated Protein in Live Bacteria. Science 2011, 333, 1445–1449. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, H.M.; Schroeder, J.; Freddolino, P.L. Nucleoid-Associated Proteins Shape Chromatin Structure and Transcriptional Regulation across the Bacterial Kingdom. Transcription 2021, 12, 182–218. [Google Scholar] [CrossRef] [PubMed]
- Norris, V.; Kayser, C.; Muskhelishvili, G.; Konto-Ghiorghi, Y. The Roles of Nucleoid-Associated Proteins and Topoisomerases in Chromosome Structure, Strand Segregation, and the Generation of Phenotypic Heterogeneity in Bacteria. FEMS Microbiol. Rev. 2022, 46, fuac049. [Google Scholar] [CrossRef] [PubMed]
- Hołówka, J.; Zakrzewska-Czerwińska, J. Nucleoid Associated Proteins: The Small Organizers That Help to Cope with Stress. Front. Microbiol. 2020, 11, 590. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.H.; de la Tour, C.B.; Toueille, M.; Vannier, F.; Sommer, S.; Servant, P. The Essential Histone-like Protein HU Plays a Major Role in Deinococcus Radiodurans Nucleoid Compaction. Mol. Microbiol. 2009, 73, 240–252. [Google Scholar] [CrossRef]
- Salerno, P.; Larsson, J.; Bucca, G.; Laing, E.; Smith, C.P.; Flärdh, K. One of the Two Genes Encoding Nucleoid-Associated HU Proteins in Streptomyces Coelicolor Is Developmentally Regulated and Specifically Involved in Spore Maturation. J. Bacteriol. 2009, 191, 6489–6500. [Google Scholar] [CrossRef]
- Pandey, S.D.; Choudhury, M.; Yousuf, S.; Wheeler, P.R.; Gordon, S.V.; Ranjan, A.; Sritharan, M. Iron-Regulated Protein HupB of Mycobacterium Tuberculosis Positively Regulates Siderophore Biosynthesis and Is Essential for Growth in Macrophages. J. Bacteriol. 2014, 196, 1853–1865. [Google Scholar] [CrossRef]
- Crosby, H.A.; Schlievert, P.M.; Merriman, J.A.; King, J.M.; Salgado-Pabón, W.; Horswill, A.R. The Staphylococcus Aureus Global Regulator MgrA Modulates Clumping and Virulence by Controlling Surface Protein Expression. PLoS Pathog. 2016, 12, e1005604. [Google Scholar] [CrossRef]
- Odermatt, N.T.; Sala, C.; Benjak, A.; Cole, S.T. Essential Nucleoid Associated Protein MIHF (Rv1388) Controls Virulence and Housekeeping Genes in Mycobacterium Tuberculosis. Sci. Rep. 2018, 8, 14214. [Google Scholar] [CrossRef]
- Kahramanoglou, C.; Seshasayee, A.S.N.; Prieto, A.I.; Ibberson, D.; Schmidt, S.; Zimmermann, J.; Benes, V.; Fraser, G.M.; Luscombe, N.M. Direct and Indirect Effects of H-NS and Fis on Global Gene Expression Control in Escherichia coli. Nucleic Acids Res. 2011, 39, 2073–2091. [Google Scholar] [CrossRef] [PubMed]
- Brunet, Y.R.; Khodr, A.; Logger, L.; Aussel, L.; Mignot, T.; Rimsky, S.; Cascales, E. H-NS Silencing of the Salmonella Pathogenicity Island 6-Encoded Type VI Secretion System Limits Salmonella Enterica Serovar Typhimurium Interbacterial Killing. Infect. Immun. 2015, 83, 2738–2750. [Google Scholar] [CrossRef] [PubMed]
- Ryan, V.T.; Grimwade, J.E.; Nievera, C.J.; Leonard, A.C. IHF and HU Stimulate Assembly of Pre-Replication Complexes at Escherichia coli OriC by Two Different Mechanisms. Mol. Microbiol. 2002, 46, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Datta, C.; Jha, R.K.; Ganguly, S.; Nagaraja, V. NapA (Rv0430), a Novel Nucleoid-Associated Protein That Regulates a Virulence Operon in Mycobacterium Tuberculosis in a Supercoiling-Dependent Manner. J. Mol. Biol. 2019, 431, 1576–1591. [Google Scholar] [CrossRef] [PubMed]
- Ali Azam, T.; Iwata, A.; Nishimura, A.; Ueda, S.; Ishihama, A. Growth Phase-Dependent Variation in Protein Composition of the Escherichia coli Nucleoid. J. Bacteriol. 1999, 181, 6361–6370. [Google Scholar] [CrossRef]
- Chesnokov, Y.; Mozhaev, A.; Kamyshinsky, R.; Gordienko, A.; Dadinova, L. Structural Insights into Iron Ions Accumulation in Dps Nanocage. Int. J. Mol. Sci. 2022, 23, 5313. [Google Scholar] [CrossRef]
- Gordienko, A.M.; Mozhaev, A.A.; Gibizova, V.V.; Dadinova, L.A. An Anomalous Small-Angle X-Ray Scattering Study of the Formation of Iron Clusters in the Inner Cavity of the Ferritin-Like Protein Dps. Moscow Univ. Phys. Bull. 2022, 77, 858–867. [Google Scholar] [CrossRef]
- Shtykova, E.V.; Petoukhov, M.V.; Mozhaev, A.A. Formation of Iron Oxide Nanoparticles in the Internal Cavity of Ferritin-Like Dps Protein: Studies by Anomalous X-Ray Scattering. Biochemistry 2022, 87, 511–523. [Google Scholar] [CrossRef]
- Almiron, M.; Link, A.J.; Furlong, D.; Kolter, R. A Novel DNA-Binding Protein with Regulatory and Protective Roles in Starved Escherichia coli. Genes Dev. 1992, 6, 2646–2654. [Google Scholar] [CrossRef]
- Nair, S.; Finkel, S.E. Dps Protects Cells against Multiple Stresses during Stationary Phase. J. Bacteriol. 2004, 186, 4192–4198. [Google Scholar] [CrossRef]
- Karas, V.O.; Westerlaken, I.; Meyer, A.S. The DNA-Binding Protein from Starved Cells (Dps) Utilizes Dual Functions To Defend Cells against Multiple Stresses. J. Bacteriol. 2015, 197, 3206–3215. [Google Scholar] [CrossRef] [PubMed]
- Stephani, K.; Weichart, D.; Hengge, R. Dynamic Control of Dps Protein Levels by ClpXP and ClpAP Proteases in Escherichia coli. Mol. Microbiol. 2003, 49, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Janissen, R.; Arens, M.M.A.; Vtyurina, N.N.; Rivai, Z.; Sunday, N.D.; Eslami-Mossallam, B.; Gritsenko, A.A.; Laan, L.; de Ridder, D.; Artsimovitch, I.; et al. Global DNA Compaction in Stationary-Phase Bacteria Does Not Affect Transcription. Cell 2018, 174, 1188–1199.e14. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.A.; Filman, D.J.; Finkel, S.E.; Kolter, R.; Hogle, J.M. The Crystal Structure of Dps, a Ferritin Homolog That Binds and Protects DNA. Nat. Struct. Biol. 1998, 5, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Azam, T.A.; Ishihama, A. Twelve Species of the Nucleoid-Associated Protein from Escherichia coli. J. Biol. Chem. 1999, 274, 33105–33113. [Google Scholar] [CrossRef] [PubMed]
- Ceci, P. DNA Condensation and Self-Aggregation of Escherichia coli Dps Are Coupled Phenomena Related to the Properties of the N-Terminus. Nucleic Acids Res. 2004, 32, 5935–5944. [Google Scholar] [CrossRef] [PubMed]
- Kamyshinsky, R.; Chesnokov, Y.; Dadinova, L.; Mozhaev, A.; Orlov, I.; Petoukhov, M.; Orekhov, A.; Shtykova, E.; Vasiliev, A.; Kamyshinsky, R.; et al. Polymorphic Protective Dps-DNA Co-Crystals by Cryo Electron Tomography and Small Angle X-ray Scattering. Biomolecules 2019, 10, 39. [Google Scholar] [CrossRef]
- Dadinova, L.A.; Chesnokov, Y.M.; Kamyshinsky, R.A.; Orlov, I.A.; Petoukhov, M.V.; Mozhaev, A.A.; Soshinskaya, E.Y.; Lazarev, V.N.; Manuvera, V.A.; Orekhov, A.S.; et al. Protective Dps–DNA Co-crystallization in Stressed Cells: An in Vitro Structural Study by Small-angle X-ray Scattering and Cryo-electron Tomography. FEBS Lett. 2019, 593, 1360–1371. [Google Scholar] [CrossRef]
- Dubrovin, E.V.; Dadinova, L.A.; Petoukhov, M.V.; Soshinskaya, E.Y.; Mozhaev, A.A.; Klinov, D.V.; Schäffer, T.E.; Shtykova, E.V.; Batishchev, O.V. Spatial Organization of Dps and DNA–Dps Complexes. J. Mol. Biol. 2021, 433, 166930. [Google Scholar] [CrossRef] [PubMed]
- Reich, Z.; Wachtel, E.; Minsky, A. Liquid-Crystalline Mesophases of Plasmid DNA in Bacteria. Science 1994, 264, 1460–1463. [Google Scholar] [CrossRef]
- Wolf, S.G.; Frenkiel, D.; Arad, T.; Finkel, S.E.; Kolter, R.; Minsky, A. DNA Protection by Stress-Induced Biocrystallization. Nature 1999, 400, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Frenkiel-Krispin, D.; Ben-Avraham, I.; Englander, J.; Shimoni, E.; Wolf, S.G.; Minsky, A. Nucleoid Restructuring in Stationary-State Bacteria. Mol. Microbiol. 2004, 51, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Hud, N.V.; Downing, K.H. Cryoelectron Microscopy of λ Phage DNA Condensates in Vitreous Ice: The Fine Structure of DNA Toroids. Proc. Natl. Acad. Sci. USA 2001, 98, 14925–14930. [Google Scholar] [CrossRef] [PubMed]
- Dadinova, L.; Kamyshinsky, R.; Chesnokov, Y.; Mozhaev, A.; Matveev, V.; Gruzinov, A.; Vasiliev, A.; Shtykova, E. Structural Rearrangement of Dps-DNA Complex Caused by Divalent Mg and Fe Cations. Int. J. Mol. Sci. 2021, 22, 6056. [Google Scholar] [CrossRef] [PubMed]
- Soshinskaya, E.Y.; Dadinova, L.A.; Mozhaev, A.A.; Shtykova, E.V. Effect of Buffer Composition on Conformational Flexibility of N-Terminal Fragments of Dps and the Nature of Interactions with DNA. Small-Angle X-Ray Scattering Study. Crystallogr. Rep. 2020, 65, 891–899. [Google Scholar] [CrossRef]
- Guerra, J.P.L.; Blanchet, C.E.; Vieira, B.J.C.; Almeida, A.V.; Waerenborgh, J.C.; Jones, N.C.; Hoffmann, S.V.; Tavares, P.; Pereira, A.S. The Conformation of the N-Terminal Tails of Deinococcus Grandis Dps Is Modulated by the Ionic Strength. Int. J. Mol. Sci. 2022, 23, 4871. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lim, C.J.; Dröge, P.; Yan, J. Regulation of Bacterial DNA Packaging in Early Stationary Phase by Competitive DNA Binding of Dps and IHF. Sci. Rep. 2016, 5, 18146. [Google Scholar] [CrossRef]
- Frenkiel-Krispin, D. Regulated Phase Transitions of Bacterial Chromatin: A Non-Enzymatic Pathway for Generic DNA Protection. EMBO J. 2001, 20, 1184–1191. [Google Scholar] [CrossRef]
- Hammel, M.; Amlanjyoti, D.; Reyes, F.E.; Chen, J.-H.; Parpana, R.; Tang, H.Y.H.; Larabell, C.A.; Tainer, J.A.; Adhya, S. HU Multimerization Shift Controls Nucleoid Compaction. Sci. Adv. 2016, 2, e1600650. [Google Scholar] [CrossRef]
- Kashchiev, D. Nucleation, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2000; ISBN 9780750646826. [Google Scholar]
- Van Driessche, A.E.S.S.; Van Gerven, N.; Bomans, P.H.H.H.; Joosten, R.R.M.M.; Friedrich, H.; Gil-Carton, D.; Sommerdijk, N.A.J.M.J.M.; Sleutel, M. Molecular Nucleation Mechanisms and Control Strategies for Crystal Polymorph Selection. Nature 2018, 556, 89–94. [Google Scholar] [CrossRef]
- Hud, N.V.; Vilfan, I.D. Toroidal DNA Condensates: Unraveling the Fine Structure and the Role of Nucleation in Determining Size. Annu. Rev. Biophys. Biomol. Struct. 2005, 34, 295–318. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Tibbelin, G.; Kajino, T.; Asami, O.; Ladenstein, R. The Multi-Layered Structure of Dps with a Novel Di-Nuclear Ferroxidase Center. J. Mol. Biol. 2003, 329, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Joshi, A.; Arora, K.; Mukhopadhyay, S.; Guptasarma, P. Nucleoid-Associated Proteins Undergo Liquid-Liquid Phase Separation with DNA into Multiphasic Condensates Resembling Bacterial Nucleoids. bioRxiv 2022. [Google Scholar] [CrossRef]
- He, F.E. E. coli Genomic DNA Extraction. Bio-Protocol 2011, 1, e97. [Google Scholar] [CrossRef]
- Vingataramin, L.; Frost, E.H. A Single Protocol for Extraction of GDNA from Bacteria and Yeast. Biotechniques 2015, 58, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Kremer, J.R.; Mastronarde, D.N.; McIntosh, J.R. Computer Visualization of Three-Dimensional Image Data Using IMOD. J. Struct. Biol. 1996, 116, 71–76. [Google Scholar] [CrossRef]
- Liu, Y.-T.; Zhang, H.; Wang, H.; Tao, C.-L.; Bi, G.-Q.; Zhou, Z.H. Isotropic Reconstruction for Electron Tomography with Deep Learning. Nat. Commun. 2022, 13, 6482. [Google Scholar] [CrossRef]
- Tang, G.; Peng, L.; Baldwin, P.R.; Mann, D.S.; Jiang, W.; Rees, I.; Ludtke, S.J. EMAN2: An Extensible Image Processing Suite for Electron Microscopy. J. Struct. Biol. 2007, 157, 38–46. [Google Scholar] [CrossRef]
- Galaz-Montoya, J.G.; Flanagan, J.; Schmid, M.F.; Ludtke, S.J. Single Particle Tomography in EMAN2. J. Struct. Biol. 2015, 190, 279–290. [Google Scholar] [CrossRef]
- Scheres, S.H.W. RELION: Implementation of a Bayesian Approach to Cryo-EM Structure Determination. J. Struct. Biol. 2012, 180, 519–530. [Google Scholar] [CrossRef]
- Kimanius, D.; Forsberg, B.O.; Scheres, S.H.W.; Lindahl, E. Accelerated Cryo-EM Structure Determination with Parallelisation Using GPUs in RELION-2. eLife 2016, 5, e18722. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chesnokov, Y.; Kamyshinsky, R.; Mozhaev, A.; Shtykova, E.; Vasiliev, A.; Orlov, I.; Dadinova, L. Morphological Diversity of Dps Complex with Genomic DNA. Int. J. Mol. Sci. 2023, 24, 8534. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24108534
Chesnokov Y, Kamyshinsky R, Mozhaev A, Shtykova E, Vasiliev A, Orlov I, Dadinova L. Morphological Diversity of Dps Complex with Genomic DNA. International Journal of Molecular Sciences. 2023; 24(10):8534. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24108534
Chicago/Turabian StyleChesnokov, Yuri, Roman Kamyshinsky, Andrey Mozhaev, Eleonora Shtykova, Alexander Vasiliev, Ivan Orlov, and Liubov Dadinova. 2023. "Morphological Diversity of Dps Complex with Genomic DNA" International Journal of Molecular Sciences 24, no. 10: 8534. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24108534