

Advanced Biomarkers of Hepatotoxicity in Psychiatry: A Narrative Review and Recommendations for New Psychoactive Substances

Abstract
:1. Introduction
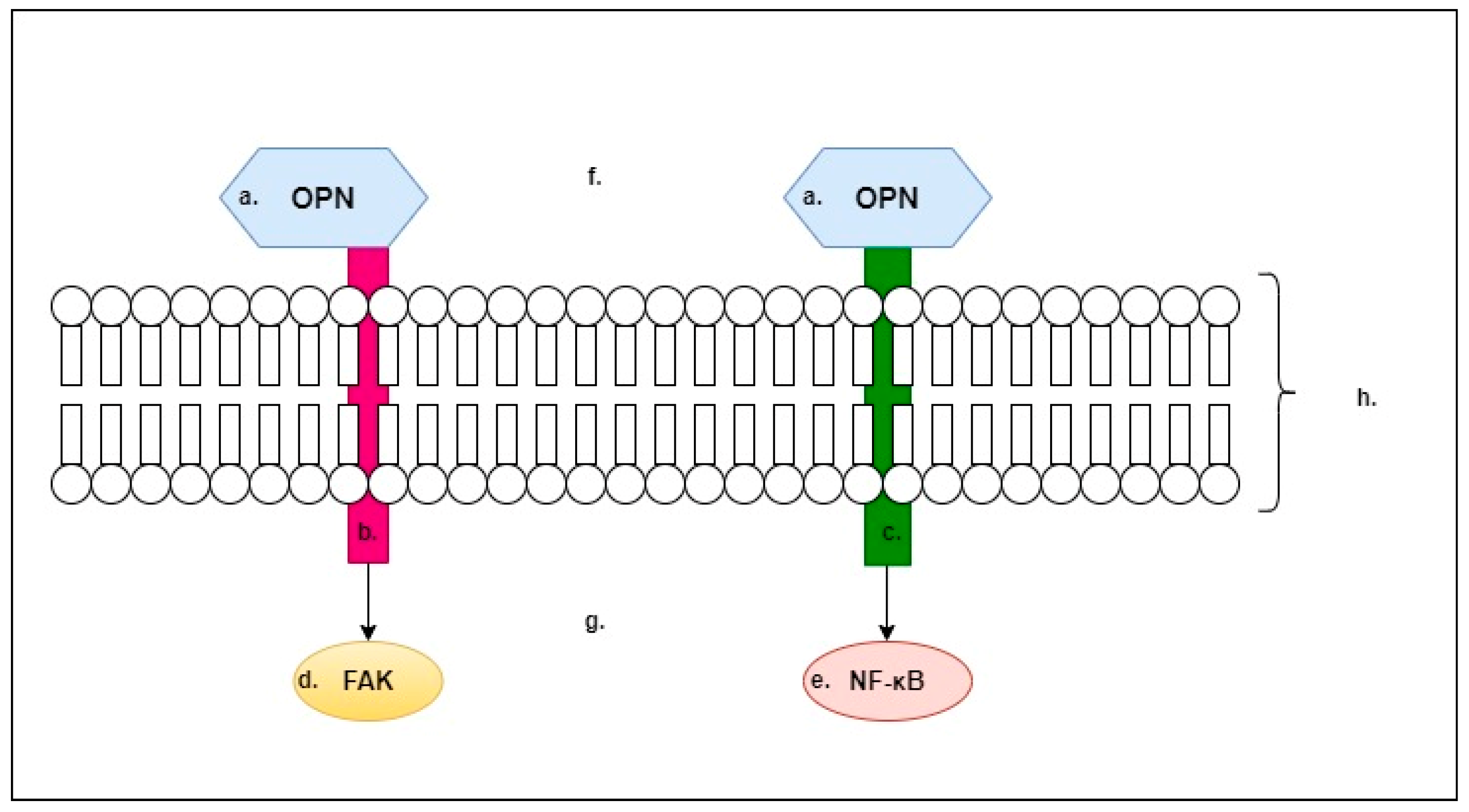
2. Osteopontin (OPN)
2.1. OPN in Psychiatry
2.2. OPN and Use of Psychoactive Substances
3. High-Mobility Group Box 1 Protein (HMGB1; HMG-1; Amphoterin)
3.1. HMGB1 in Psychiatry
3.2. HMGB1 and Use of Psychoactive Substances
4. Glutamate Dehydrogenase (GDH, GLDH)
4.1. GDH in Psychiatry
4.2. GDH and Use of Psychoactive Substances
5. Recommendations for Hepatotoxicity Testing of New Psychoactive Substances
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AIM2 | AIM2 inflammasome |
| ALD | alcoholic liver disease |
| ALT | alanine aminotransferase |
| AST | aspartate aminotransferase |
| BAL | bronchoalveolar lavage |
| BDKRB1 | bradykinin receptor B1 |
| CCL11 | eotaxin |
| CD44 | cluster of differentiation 44 |
| CDI | clostridium difficile infection |
| COPD | chronic obstructive pulmonary disease |
| COX | cyclooxygenase |
| CSDS | chronic social defeat stress |
| CUMS | chronic unpredictable mild stress |
| DILI | drug-induced liver injury |
| DIP | desquamative interstitial pneumonia |
| DKK1 | dickkopf-related protein 1 |
| EGF | epidermal growth factor |
| EGFR | epidermal growth factor receptor |
| FAK | focal adhesion kinase |
| FEP | first psychotic episode |
| Gcy | glycyrrhizin |
| GDH, GLDH | glutamate dehydrogenase |
| GGT | gamma-glutamyltranspeptidase |
| GLP-1 | glucagon-like peptide-1 |
| GLUD1 | gene encodes glutamate dehydrogenase |
| HCV | hepatitis C virus |
| HIV | human immunodeficiency virus |
| HMGB1 | high mobility group box 1 protein; amphoterin |
| IL-1β | interleukin-1β |
| IL6 | interleukin-6 |
| IL33 | interleukin-33 |
| KOR | kappa opioid receptors |
| KP | kynurenine |
| LPS | lipopolysaccharide |
| MCP-1 | monocyte chemotactic protein 1 |
| MCV | mean erythrocyte volume |
| MDD | major depressive disorder |
| MMP1/MMP3/MMP9 | matrix metalloproteinase-1/-3/-9 |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| NAFLD | non-alcoholic fatty liver disease |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NLRP | nucleotide-binding oligomerization domain |
| nPM | nano-sized particulate matter (nPM) |
| NPSs | new psychoactive substances |
| NCSLS | non-small cell lung cancer |
| OPN | osteopontin |
| PBMCs | peripheral blood mononuclear cells |
| PCR | polymerase chain reaction |
| PDE4 | phosphodiesterase-4 |
| PLCH | Langerhans cell histiocytosis |
| RAGE | receptor for advanced glycation end products |
| RANKL | receptor activator of NF-kappaB ligand |
| SPP1 | secreted phosphoprotein 1 |
| TLR2/TLR4 | toll-like receptor 2/4 |
| TMEM168 | transmembrane protein 168 |
| TNF-α | tumour necrosis factor alpha |
References
- Peacock, A.; Bruno, R.; Gisev, N.; Degenhardt, L.; Hall, W.; Sedefov, R.; White, J.; Thomas, K.V.; Farrell, M.; Griffiths, P. New psychoactive substances: Challenges for drug surveillance, control, and public health responses. Lancet 2019, 394, 1668–1684. [Google Scholar] [CrossRef]
- Ordak, M.; Nasierowski, T.; Muszynska, E.; Bujalska-Zadrozny, M. Psychoactive Substances Taken with Mephedrone and HCV Infection. J. Clin. Med. 2021, 10, 3218. [Google Scholar] [CrossRef]
- Ordak, M.; Nasierowski, T.; Pawlik, K.; Muszynska, E.; Bujalska-Zadrozny, M. Effectiveness of the methadone programme in the treatment of patients on a mephedrone binge and dependent on heroin: A retrospective study, 2010–2019. Int. J. Psychiatry Clin. Pract. 2020, 24, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Ordak, M.; Nasierowski, T.; Muszynska, E.; Bujalska-Zadrozny, M. The Psychiatric Characteristics of People on a Mephedrone (“bath salts”) Binge. Subst. Use Misuse 2020, 55, 1610–1617. [Google Scholar] [CrossRef]
- Ordak, M.; Nasierowski, T.; Muszynska, E.; Bujalska-Zadrozny, M. Optimisation of methadone treatment in a group of patients on a mephedrone binge and dependent on many psychoactive substances. Int. J. Psychiatry Clin. Pract. 2020, 24, 38–42. [Google Scholar] [CrossRef]
- Ordak, M.; Zmysłowska, A.; Bielski, M.; Rybak, D.; Tomaszewska, M.; Wyszomierska, K.; Kmiec, A.; Garlicka, N.; Zalewska, M.; Zalewski, M.; et al. Pharmacotherapy of Patients Taking New Psychoactive Substances: A Systematic Review and Analysis of Case Reports. Front. Psychiatry 2021, 12, 669921. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Wu, D.; Jiang, W.; Li, J.; Long, J.; Jia, C.; Zhou, T. Molecular Biomarkers in Drug-Induced Liver Injury: Challenges and Future Perspectives. Front. Pharmacol. 2019, 10, 1667. [Google Scholar] [CrossRef]
- Bruha, R.; Vitek, L.; Smid, V. Osteopontin–A potential biomarker of advanced liver disease. Ann. Hepatol. 2020, 19, 344–352. [Google Scholar] [CrossRef]
- Schomaker, S.; Potter, D.; Warner, R.; Larkindale, J.; King, N.; Porter, A.C.; Owens, J.; Tomlinson, L.; Sauer, J.-M.; Johnson, K.; et al. Serum glutamate dehydrogenase activity enables early detection of liver injury in subjects with underlying muscle impairments. PLoS ONE 2020, 15, e0229753. [Google Scholar] [CrossRef]
- Gaskell, H.; Ge, X.; Nieto, N. High-Mobility Group Box-1 and Liver Disease. Hepatol. Commun. 2018, 2, 1005–1020. [Google Scholar] [CrossRef] [PubMed]
- Oton-Gonzalez, L.; Mazziotta, C.; Iaquinta, M.R.; Mazzoni, E.; Nocini, R.; Trevisiol, L.; D’agostino, A.; Tognon, M.; Rotondo, J.C.; Martini, F. Genetics and Epigenetics of Bone Remodeling and Metabolic Bone Diseases. Int. J. Mol. Sci. 2022, 23, 1500. [Google Scholar] [CrossRef]
- Chen, L.; Huan, X.; Xiao, G.-H.; Yu, W.-H.; Li, T.-F.; Gao, X.-D.; Zhang, Y.-C. Osteopontin and its downstream carcinogenic molecules: Regulatory mechanisms and prognostic value in cancer progression. Neoplasma 2022, 69, 1253–1269. [Google Scholar] [CrossRef]
- Si, J.; Wang, C.; Zhang, D.; Wang, B.; Zhou, Y. Osteopontin in Bone Metabolism and Bone Diseases. Med. Sci. Monit. 2020, 26, e919159. [Google Scholar] [CrossRef] [PubMed]
- Nuñez-Garcia, M.; Gomez-Santos, B.; Buqué, X.; García-Rodriguez, J.L.; Romero, M.R.; Marin, J.J.; Arteta, B.; García-Monzón, C.; Castaño, L.; Syn, W.-K.; et al. Osteopontin regulates the cross-talk between phosphatidylcholine and cholesterol metabolism in mouse liver. J. Lipid Res. 2017, 58, 1903–1915. [Google Scholar] [CrossRef] [PubMed]
- Iida, T.; Wagatsuma, K.; Hirayama, D.; Nakase, H. Is Osteopontin a Friend or Foe of Cell Apoptosis in Inflammatory Gastrointestinal and Liver Diseases? Int. J. Mol. Sci. 2017, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Rosmus, D.-D.; Lange, C.; Ludwig, F.; Ajami, B.; Wieghofer, P. The Role of Osteopontin in Microglia Biology: Current Concepts and Future Perspectives. Biomedicines 2022, 10, 840. [Google Scholar] [CrossRef]
- Sun, Z.; Andersson, R. NF-kappaB Activation and Inhibition: A Review. Shock 2002, 18, 99–106. [Google Scholar] [CrossRef]
- Wen, Y.; Jeong, S.; Xia, Q.; Kong, X. Role of Osteopontin in Liver Diseases. Int. J. Biol. Sci. 2016, 12, 1121–1128. [Google Scholar] [CrossRef]
- Song, Z.; Chen, W.; Athavale, D.; Ge, X.; Desert, R.; Das, S.; Han, H.; Nieto, N. Osteopontin Takes Center Stage in Chronic Liver Disease. Hepatology 2020, 73, 1594–1608. [Google Scholar] [CrossRef]
- Srungaram, P.; Rule, J.A.; Yuan, H.J.; Reimold, A.; Dahl, B.; Sanders, C.; Lee, W.M.; Acute Liver Failure Study Group. Plasma osteopontin in acute liver failure. Cytokine 2015, 73, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Kariya, Y.; Kariya, Y. Osteopontin in Cancer: Mechanisms and Therapeutic Targets. Int. J. Transl. Med. 2022, 2, 419–447. [Google Scholar] [CrossRef]
- Arriazu, E.; Ge, X.; Leung, T.-M.; Magdaleno, F.; Lopategi, A.; Lu, Y.; Kitamura, N.; Urtasun, R.; Theise, N.; Antoine, D.J.; et al. Signalling via the osteopontin and high mobility group box-1 axis drives the fibrogenic response to liver injury. Gut 2017, 66, 1123–1137. [Google Scholar] [CrossRef]
- Seth, D.; Duly, A.; Kuo, P.C.; McCaughan, G.W.; Haber, P.S. Osteopontin is an important mediator of alcoholic liver disease via hepatic stellate cell activation. World J. Gastroenterol. 2014, 20, 13088–13104. [Google Scholar] [CrossRef] [PubMed]
- Morales-Ibanez, O.; Domínguez, M.; Ki, S.H.; Marcos, M.; Chaves, J.F.; Nguyen-Khac, E.; Houchi, H.; Affò, S.; Sancho-Bru, P.; Altamirano, J.; et al. Human and experimental evidence supporting a role for osteopontin in alcoholic hepatitis. Hepatology 2013, 58, 1742–1756. [Google Scholar] [CrossRef]
- Orman, E.S.; Odena, G.; Bataller, R. Alcoholic liver disease: Pathogenesis, management, and novel targets for therapy. J. Gastroenterol. Hepatol. 2013, 28 (Suppl. S1), 77–84. [Google Scholar] [CrossRef]
- Rittling, S.R.; Singh, R. Osteopontin in Immune-mediated Diseases. J. Dent. Res. 2015, 94, 1638–1645. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.F.; Lett, G.S.; Haubein, N.C. Osteopontin is a marker for cancer aggressiveness and patient survival. Br. J. Cancer 2010, 103, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Castello, L.M.; Raineri, D.; Salmi, L.; Clemente, N.; Vaschetto, R.; Quaglia, M.; Garzaro, M.; Gentilli, S.; Navalesi, P.; Cantaluppi, V.; et al. Osteopontin at the Crossroads of Inflammation and Tumor Progression. Mediat. Inflamm. 2017, 2017, 4049098. [Google Scholar] [CrossRef]
- Cabiati, M.; Gaggini, M.; Cesare, M.M.; Caselli, C.; De Simone, P.; Filipponi, F.; Basta, G.; Gastaldelli, A.; Del Ry, S. Osteopontin in hepatocellular carcinoma: A possible biomarker for diagnosis and follow-up. Cytokine 2017, 99, 59–65. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhou, C.; Yu, X.-X.; Wu, C.; Jia, H.-L.; Gao, X.-M.; Yang, J.-M.; Wang, C.-Q.; Luo, Q.; Zhu, Y.; et al. Osteopontin promotes metastasis of intrahepatic cholangiocarcinoma through recruiting MAPK1 and mediating Ser675 phosphorylation of β-Catenin. Cell Death Dis. 2018, 9, 179. [Google Scholar] [CrossRef]
- Shirakawa, K.; Sano, M. Osteopontin in Cardiovascular Diseases. Biomolecules 2021, 11, 1047. [Google Scholar] [CrossRef]
- Vianello, S.; Pantic, B.; Fusto, A.; Bello, L.; Galletta, E.; Borgia, D.; Gavassini, B.F.; Semplicini, C.; Sorarù, G.; Vitiello, L.; et al. SPP1 genotype and glucocorticoid treatment modify osteopontin expression in Duchenne muscular dystrophy cells. Hum. Mol. Genet. 2017, 26, 3342–3351. [Google Scholar] [CrossRef] [PubMed]
- Hylén, U.; McGlinchey, A.; Orešič, M.; Bejerot, S.; Humble, M.B.; Särndahl, E.; Hyötyläinen, T.; Eklund, D. Potential Transdiagnostic Lipid Mediators of Inflammatory Activity in Individuals With Serious Mental Illness. Front. Psychiatry 2021, 12, 778325. [Google Scholar] [CrossRef] [PubMed]
- Mantere, O.; Trontti, K.; García-González, J.; Balcells, I.; Saarnio, S.; Mäntylä, T.; Lindgren, M.; Kieseppä, T.; Raij, T.; Honkanen, J.K.; et al. Immunomodulatory effects of antipsychotic treatment on gene expression in first-episode psychosis. J. Psychiatr. Res. 2018, 109, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Kovács, M.; Tényi, T.; Kugyelka, R.; Prenek, L.; Hau, L.; Magyar, E.; Herold, R.; Balogh, P.; Simon, D. Elevated Osteopontin and Interferon Gamma Serum Levels and Increased Neutrophil-to-Lymphocyte Ratio Are Associated With the Severity of Symptoms in Schizophrenia. Front. Psychiatry 2019, 10, 996. [Google Scholar] [CrossRef] [PubMed]
- Guest, P.C.; Urday, S.; Ma, D.; Stelzhammer, V.; Harris, L.W.; Amess, B.; Pietsch, S.; Oheim, C.; Ozanne, S.E.; Bahn, S. Proteomic analysis of the maternal protein restriction rat model for schizophrenia: Identification of translational changes in hormonal signaling pathways and glutamate neurotransmission. Proteomics 2012, 12, 3580–3589. [Google Scholar] [CrossRef] [PubMed]
- Kutlu, Z.H.Z.; Ozcan, H. Comparison of Serum Osteopontin Levels with Healthy Controls in Schizophrenia Patients. Fırat Tıp Dergisi/Firat Med. J. 2021, 26, 74–78. [Google Scholar]
- Çakici, N.; Bot, M.; Lamers, F.; Janssen, T.; van der Spek, P.J.; de Haan, L.; Bahn, S.; Penninx, B.W.; van Beveren, N.J. Increased serum levels of leptin and insulin in both schizophrenia and major depressive disorder: A cross-disorder proteomics analysis. Eur. Neuropsychopharmacol. 2019, 29, 835–846. [Google Scholar] [CrossRef]
- Zhang, K.; Ma, M.; Dong, C.; Hashimoto, K. Role of Inflammatory Bone Markers in the Antidepressant Actions of (R)-Ketamine in a Chronic Social Defeat Stress Model. Int. J. Neuropsychopharmacol. 2018, 21, 1025–1030. [Google Scholar] [CrossRef]
- Kadriu, B.; Gold, P.W.; Luckenbaugh, D.A.; Lener, M.S.; Ballard, E.D.; Niciu, M.J.; Henter, I.D.; Park, L.T.; De Sousa, R.T.; Yuan, P.; et al. Acute ketamine administration corrects abnormal inflammatory bone markers in major depressive disorder. Mol. Psychiatry 2018, 23, 1626–1631. [Google Scholar] [CrossRef]
- Xiong, Z.; Fujita, Y.; Zhang, K.; Pu, Y.; Chang, L.; Ma, M.; Chen, J.; Hashimoto, K. Beneficial effects of (R)-ketamine, but not its metabolite (2R,6R)-hydroxynorketamine, in the depression-like phenotype, inflammatory bone markers, and bone mineral density in a chronic social defeat stress model. Behav. Brain Res. 2019, 368, 111904. [Google Scholar] [CrossRef] [PubMed]
- Stelzhammer, V.; Guest, P.C.; Rothermundt, M.; Sondermann, C.; Michael, N.; Schwarz, E.; Rahmoune, H.; Bahn, S. Electroconvulsive therapy exerts mainly acute molecular changes in serum of major depressive disorder patients. Eur. Neuropsychopharmacol. 2013, 23, 1199–1207. [Google Scholar] [CrossRef]
- Zhang, J.; Fujita, Y.; Chang, L.; Pu, Y.; Qu, Y.; Wang, S.; Hashimoto, K. Beneficial effects of anti-RANKL antibody in depression-like phenotype, inflammatory bone markers, and bone mineral density in male susceptible mice after chronic social defeat stress. Behav. Brain Res. 2020, 379, 112397. [Google Scholar] [CrossRef] [PubMed]
- Cleland, B.T.; Papanek, P.; Ingraham, B.A.; Harkins, A.; Garnier-Villarreal, M.; Woo, D.; Csuka, M.; Ng, A.V. Determinants of low bone mineral density in people with multiple sclerosis: Role of physical activity. Mult. Scler. Relat. Disord. 2020, 38, 101864. [Google Scholar] [CrossRef] [PubMed]
- Huan, Y.; Wei, J.; Su, T.; Gao, Y. Urine proteome changes in a chronic unpredictable mild stress (CUMS) mouse model of major depressive disorder. J. Pharm. Biomed. Anal. 2021, 199, 114064. [Google Scholar] [CrossRef]
- Cho, H.H.; Park, H.T.; Kim, Y.J.; Bae, Y.C.; Suh, K.T.; Jung, J.S. Induction of osteogenic differentiation of human mesenchymal stem cells by histone deacetylase inhibitors. J. Cell. Biochem. 2005, 96, 533–542. [Google Scholar] [CrossRef]
- Ventorp, F.; Barzilay, R.; Erhardt, S.; Samuelsson, M.; Träskman-Bendz, L.; Janelidze, S.; Weizman, A.; Offen, D.; Brundin, L. The CD44 ligand hyaluronic acid is elevated in the cerebrospinal fluid of suicide attempters and is associated with increased blood–brain barrier permeability. J. Affect. Disord. 2016, 193, 349–354. [Google Scholar] [CrossRef]
- Wanby, P.; Brudin, L.; Von, S.-P.; Carlsson, M. Modestly degraded microarchitecture and high serum levels of osteopontin in Swedish females with anorexia nervosa. Eat. Weight. Disord. 2020, 26, 2165–2172. [Google Scholar] [CrossRef]
- Al-Ayadhi, L.Y.; Mostafa, G.A. Increased serum osteopontin levels in autistic children: Relation to the disease severity. Brain Behav. Immun. 2011, 25, 1393–1398. [Google Scholar] [CrossRef]
- Xu, N.; Li, X.; Zhong, Y. Inflammatory Cytokines: Potential Biomarkers of Immunologic Dysfunction in Autism Spectrum Disorders. Mediat. Inflamm. 2015, 2015, 531518. [Google Scholar] [CrossRef]
- Liu, W.; Li, L.; Xia, X.; Zhou, X.; Du, Y.; Yin, Z.; Wang, J. Integration of Urine Proteomic and Metabolomic Profiling Reveals Novel Insights Into Neuroinflammation in Autism Spectrum Disorder. Front. Psychiatry 2022, 13, 780747. [Google Scholar] [CrossRef] [PubMed]
- Silva, K.; Hope-Lucas, C.; White, T.; Hairston, T.-K.; Rameau, T.; Brown, A. Cortical neurons are a prominent source of the proinflammatory cytokine osteopontin in HIV-associated neurocognitive disorders. J. NeuroVirol. 2015, 21, 174–185. [Google Scholar] [CrossRef] [PubMed]
- Patouraux, S.; Bonnafous, S.; Voican, C.S.; Anty, R.; Saint-Paul, M.-C.; Rosenthal-Allieri, M.-A.; Agostini, H.; Njike, M.; Barri-Ova, N.; Naveau, S.; et al. The Osteopontin Level in Liver, Adipose Tissue and Serum Is Correlated with Fibrosis in Patients with Alcoholic Liver Disease. PLoS ONE 2012, 7, e35612. [Google Scholar] [CrossRef] [PubMed]
- Simão, A.; Madaleno, J.; Silva, N.; Rodrigues, F.M.R.; Caseiro, P.; Costa, J.N.; Carvalho, A. Plasma osteopontin is a biomarker for the severity of alcoholic liver cirrhosis, not for hepatocellular carcinoma screening. BMC Gastroenterol. 2015, 15, 73. [Google Scholar] [CrossRef]
- Hamid, O.I.A.; Sabik, L.M.E.; Abdelfadeel, K.F.; Shaban, S.F. Tramadol aggravates cardiovascular toxicity in a rat model of alcoholism: Involvement of intermediate microfilament proteins and immune-expressed osteopontin. J. Biochem. Mol. Toxicol. 2021, 35, 1–15. [Google Scholar] [CrossRef]
- Das, S.; Song, Z.; Han, H.; Ge, X.; Desert, R.; Athavale, D.; Komakula, S.S.B.; Magdaleno, F.; Chen, W.; Lantvit, D.; et al. Intestinal Osteopontin Protects From Alcohol-induced Liver Injury by Preserving the Gut Microbiome and the Intestinal Barrier Function. Cell. Mol. Gastroenterol. Hepatol. 2022, 14, 813–839. [Google Scholar] [CrossRef]
- Suri, A.; Singh, N.; Bansal, S.K. A Study on the Serum γ-Glutamyltranspeptidase and Plasma Osteopontin in Alcoholic Liver Disease. J. Lab. Physicians 2022, 14, 101–108. [Google Scholar] [CrossRef]
- Jiang, Y.-J.; Chao, C.-C.; Chang, A.-C.; Chen, P.-C.; Cheng, F.-J.; Liu, J.-F.; Liu, P.-I.; Huang, C.-L.; Guo, J.-H.; Huang, W.-C.; et al. Cigarette smoke-promoted increases in osteopontin expression attract mesenchymal stem cell recruitment and facilitate lung cancer metastasis. J. Adv. Res. 2022, 41, 77–87. [Google Scholar] [CrossRef]
- Rychlíková, J.; Vecka, M.; Jáchymová, M.; Macášek, J.; Hrabák, P.; Zeman, M.; Vávrová, L.; Řoupal, J.; Krechler, T.; ák, A. Osteopontin as a discriminating marker for pancreatic cancer and chronic pancreatitis. Cancer Biomark. 2016, 17, 55–65. [Google Scholar] [CrossRef]
- Weber, C.E.; Erşahin, H.; Kuo, P.C.; Mi, Z. Pancreatic Cancer and Osteopontin. Pancreas 2016, 45, e35–e36. [Google Scholar] [CrossRef]
- Cao, J.; Li, J.; Sun, L.; Qin, T.; Xiao, Y.; Chen, K.; Qian, W.; Duan, W.; Lei, J.; Ma, J.; et al. Hypoxia-driven paracrine osteopontin/integrin αvβ3 signaling promotes pancreatic cancer cell epithelial–mesenchymal transition and cancer stem cell-like properties by modulating forkhead box protein M1. Mol. Oncol. 2019, 13, 228–245. [Google Scholar] [CrossRef] [PubMed]
- Niikura, Y.; Ishii, T.; Hosoki, K.; Nagase, T.; Yamashita, N. Ovary-dependent emphysema augmentation and osteopontin induction in adult female mice. Biochem. Biophys. Res. Commun. 2015, 461, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Shan, M.; You, R.; Yuan, X.; Frazier, M.V.; Porter, P.; Seryshev, A.; Hong, J.-S.; Song, L.-Z.; Zhang, Y.; Hilsenbeck, S.; et al. Agonistic induction of PPARγ reverses cigarette smoke-induced emphysema. J. Clin. Investig. 2014, 124, 1371–1381. [Google Scholar] [CrossRef]
- Sun, D.; Ding, D.; Li, Q.; Xie, M.; Xu, Y.; Liu, X. The preventive and therapeutic effects of AAV1-KLF4-shRNA in cigarette smoke-induced pulmonary hypertension. J. Cell. Mol. Med. 2021, 25, 1238–1251. [Google Scholar] [CrossRef]
- Ehnert, S.; Aspera-Werz, R.H.; Ihle, C.; Trost, M.; Zirn, B.; Flesch, I.; Schröter, S.; Relja, B.; Nussler, A.K. Smoking Dependent Alterations in Bone Formation and Inflammation Represent Major Risk Factors for Complications Following Total Joint Arthroplasty. J. Clin. Med. 2019, 8, 406. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.-J.; Li, Y.-S.; Zhang, F.-J. Osteopontin, a bridge links osteoarthritis and osteoporosis. Front. Endocrinol. 2022, 13, 1012508. [Google Scholar] [CrossRef]
- Kimura, H.; Konno, S.; Nakamaru, Y.; Makita, H.; Taniguchi, N.; Shimizu, K.; Suzuki, M.; Ono, J.; Ohta, S.; Izuhara, K.; et al. Sinus Computed Tomographic Findings in Adult Smokers and Nonsmokers with Asthma. Analysis of Clinical Indices and Biomarkers. Ann. Am. Thorac. Soc. 2017, 14, 332–341. [Google Scholar] [CrossRef]
- Ge, Q.; Ruan, C.-C.; Ma, Y.; Tang, X.-F.; Wu, Q.-H.; Wang, J.-G.; Zhu, D.-L.; Gao, P.-J. Osteopontin regulates macrophage activation and osteoclast formation in hypertensive patients with vascular calcification. Sci. Rep. 2017, 7, 40253. [Google Scholar] [CrossRef] [PubMed]
- Arnez, M.F.M.; Monteiro, P.M.; Paula-Silva, F.W.G.; Dessotti, G.B.; de Menezes, L.M.; Küchler, E.C.; Alves, S.Y.F.; Matsumoto, M.A.N.; Stuani, M.B.S. Impact of cigarette smoke on osteogenic and osteoclast signaling in middle palatal suture. Braz. Dent. J. 2022, 33, 99–108. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, B.; Zhu, J.; Wang, D.; Wang, Y. Nicotine-mediated autophagy of vascular smooth muscle cell accelerates atherosclerosis via nAChRs/ROS/NF-kappaB signaling pathway. Atherosclerosis 2019, 284, 1–10. [Google Scholar] [CrossRef]
- Konno, S.; Taniguchi, N.; Makita, H.; Nakamaru, Y.; Shimizu, K.; Shijubo, N.; Fuke, S.; Takeyabu, K.; Oguri, M.; Kimura, H.; et al. Distinct Phenotypes of Cigarette Smokers Identified by Cluster Analysis of Patients with Severe Asthma. Ann. Am. Thorac. Soc. 2015, 12, 1771–1780. [Google Scholar] [CrossRef]
- Maneechotesuwan, K.; Kasetsinsombat, K.; Wongkajornsilp, A.; Barnes, P.J. Simvastatin up-regulates adenosine deaminase and suppresses osteopontin expression in COPD patients through an IL-13-dependent mechanism. Respir. Res. 2016, 17, 104. [Google Scholar] [CrossRef]
- Bishop, E.; Theophilus, E.H.; Fearon, I.M. In vitro and clinical studies examining the expression of osteopontin in cigarette smoke-exposed endothelial cells and cigarette smokers. BMC Cardiovasc. Disord. 2012, 12, 75. [Google Scholar] [CrossRef] [PubMed]
- Prasse, A.; Stahl, M.; Schulz, G.; Kayser, G.; Wang, L.; Ask, K.; Yalcintepe, J.; Kirschbaum, A.; Bargagli, E.; Zissel, G.; et al. Essential Role of Osteopontin in Smoking-Related Interstitial Lung Diseases. Am. J. Pathol. 2009, 174, 1683–1691. [Google Scholar] [CrossRef] [PubMed]
- Negri, B.M.; Pimentel, S.P.; Casati, M.Z.; Cirano, F.R.; Casarin, R.C.; Ribeiro, F.V. Impact of a chronic smoking habit on the osteo-immunoinflammatory mediators in the peri-implant fluid of clinically healthy dental implants. Arch. Oral Biol. 2016, 70, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Fu, K.; Miyamoto, Y.; Otake, K.; Sumi, K.; Saika, E.; Matsumura, S.; Sato, N.; Ueno, Y.; Seo, S.; Uno, K.; et al. Involvement of the accumbal osteopontin-interacting transmembrane protein 168 in methamphetamine-induced place preference and hyperlocomotion in mice. Sci. Rep. 2017, 7, 13084. [Google Scholar] [CrossRef]
- Nitta, A. Novel molecules-related drug dependence in mice. Nihon Yakurigaku Zasshi 2020, 155, 140–144. [Google Scholar] [CrossRef]
- Ge, Y.; Huang, M.; Yao, Y.-M. The Effect and Regulatory Mechanism of High Mobility Group Box-1 Protein on Immune Cells in Inflammatory Diseases. Cells 2021, 10, 1044. [Google Scholar] [CrossRef]
- Starkova, T.Y.; Polyanichko, A.M.; Artamonova, T.O.; Tsimokha, A.S.; Tomilin, A.N.; Chikhirzhina, E.V. Structural Characteristics of High-Mobility Group Proteins HMGB1 and HMGB2 and Their Interaction with DNA. Int. J. Mol. Sci. 2023, 24, 3577. [Google Scholar] [CrossRef]
- Wang, J.; Li, R.; Peng, Z.; Hu, B.; Rao, X.; Li, J. HMGB1 participates in LPS-induced acute lung injury by activating the AIM2 inflammasome in macrophages and inducing polarization of M1 macrophages via TLR2, TLR4, and RAGE/NF-κB signaling pathways Corrigendum in /10.3892/ijmm.2020.4530. Int. J. Mol. Med. 2020, 45, 61–80. [Google Scholar] [CrossRef]
- Schmidt, A.M.; Yan, S.D.; Yan, S.F.; Stern, D.M. The multiligand receptor RAGE as a progression factor amplifying immune and inflammatory responses. J. Clin. Investig. 2001, 108, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Son, M. The Immune Tolerance Role of the HMGB1-RAGE Axis. Cells 2021, 10, 564. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Wu, M.; Lu, Y.; Wu, X.; Yu, B.; Chen, R.; Lu, J.; Tong, H. HMGB1-activatied NLRP3 inflammasome induces thrombocytopenia in heatstroke rat. PeerJ 2022, 10, e13799. [Google Scholar] [CrossRef]
- Chen, R.; Kang, R.; Tang, D. The mechanism of HMGB1 secretion and release. Exp. Mol. Med. 2022, 54, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Sameer, A.S.; Nissar, S. Toll-Like Receptors (TLRs): Structure, Functions, Signaling, and Role of Their Polymorphisms in Colorectal Cancer Susceptibility. BioMed Res. Int. 2021, 2021, 1157023. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bloom, O.; Zhang, M.; Vishnubhakat, J.M.; Ombrellino, M.; Che, J.; Frazier, A.; Yang, H.; Ivanova, S.; Borovikova, L.; et al. HMG-1 as a Late Mediator of Endotoxin Lethality in Mice. Science 1999, 285, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Scaffidi, P.; Misteli, T.; Bianchi, M.E. Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 2002, 418, 191–195. [Google Scholar] [CrossRef]
- Xu, J.; Tao, P.; Lu, D.; Jiang, Y.; Xia, Q. Role of high-mobility group box 1 in cancer. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2022, 47, 505–511. [Google Scholar]
- Athavale, D.; Song, Z.; Desert, R.; Han, H.; Das, S.; Ge, X.; Komakula, S.S.B.; Chen, W.; Gao, S.; Lantvit, D.; et al. Ablation of high-mobility group box-1 in the liver reduces hepatocellular carcinoma but causes hyperbilirubinemia in Hippo signaling-deficient mice. Hepatol. Commun. 2022, 6, 2155–2169. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.-Q.; Jia, C.-Q.; Liu, C.-T.; Lu, X.-F.; Zhong, N.; Zhang, Z.-L.; Fan, W.; Li, Y.-Q. Serum high mobility group box chromosomal protein 1 is associated with clinicopathologic features in patients with hepatocellular carcinoma. Dig. Liver Dis. 2008, 40, 446–452. [Google Scholar] [CrossRef]
- Khambu, B.; Yan, S.; Huda, N.; Yin, X.-M. Role of High-Mobility Group Box-1 in Liver Pathogenesis. Int. J. Mol. Sci. 2019, 20, 5314. [Google Scholar] [CrossRef]
- Ge, X.; Arriazu, E.; Magdaleno, F.; Antoine, D.J.; Cruz, R.D.; Theise, N.; Nieto, N. High Mobility Group Box-1 Drives Fibrosis Progression Signaling via the Receptor for Advanced Glycation End Products in Mice. Hepatology 2018, 68, 2380–2404. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.H.; Park, J.H.; Jee, M.H.; Keum, S.J.; Cho, M.-S.; Yoon, S.K.; Jang, S.K. Hepatitis C Virus Infection Is Blocked by HMGB1 Released from Virus-Infected Cells. J. Virol. 2011, 85, 9359–9368. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Ming, B.; Dong, L. The Role of HMGB1 in Rheumatic Diseases. Front. Immunol. 2022, 13, 815257. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Cui, X.; Liu, Q. Emerging role of HMGB1 in lung diseases: Friend or foe. J. Cell. Mol. Med. 2016, 21, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Peng, X.; Li, H.; Chen, F.; Chen, Y.; Zhang, Y.; Le, K. The performance of the alarmin HMGB1 in pediatric diseases: From lab to clinic. Immun. Inflamm. Dis. 2021, 9, 8–30. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Chen, R.; Zhang, Q.; Hou, W.; Wu, S.; Cao, L.; Huang, J.; Yu, Y.; Fan, X.-G.; Yan, Z.; et al. HMGB1 in health and disease. Mol. Asp. Med. 2014, 40, 1–116. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Cheng, N.; Zhang, C. Schizophrenia and Alarmins. Medicina 2022, 58, 694. [Google Scholar] [CrossRef]
- Al-Dujaili, A.H.; Mousa, R.F.; Al-Hakeim, H.K.; Maes, M. High Mobility Group Protein 1 and Dickkopf-Related Protein 1 in Schizophrenia and Treatment-Resistant Schizophrenia: Associations With Interleukin-6, Symptom Domains, and Neurocognitive Impairments. Schizophr. Bull. 2021, 47, 530–541. [Google Scholar] [CrossRef]
- Kozłowska, E.; Brzezińska-Błaszczyk, E.; Agier, J.; Wysokiński, A.; Żelechowska, P. Alarmins (IL-33, sST2, HMGB1, and S100B) as potential biomarkers for schizophrenia. J. Psychiatr. Res. 2021, 138, 380–387. [Google Scholar] [CrossRef]
- Yilmaz, N.; Yelboga, Z.; Yilmaz, Y.; Demirpence, O. High mobility group box-1 levels in schizophrenia: Potential biomarker of remission phase. J. Med. Biochem. 2021, 40, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Mousa, R.F.; Al-Hakeim, H.K.; Alhaideri, A.; Maes, M. Chronic fatigue syndrome and fibromyalgia-like symptoms are an integral component of the phenome of schizophrenia: Neuro-immune and opioid system correlates. Metab. Brain Dis. 2021, 36, 169–183. [Google Scholar] [CrossRef] [PubMed]
- Al-Hakeim, H.K.; Mousa, R.F.; Al-Dujaili, A.H.; Maes, M. In schizophrenia, non-remitters and partial remitters to treatment with antipsychotics are qualitatively distinct classes with respect to neurocognitive deficits and neuro-immune biomarkers: Results of soft independent modeling of class analogy. Metab. Brain Dis. 2021, 36, 939–955. [Google Scholar] [CrossRef] [PubMed]
- Almulla, A.F.; Al-Rawi, K.F.; Maes, M.; Al-Hakeim, H.K. In schizophrenia, immune-inflammatory pathways are strongly associated with depressive and anxiety symptoms, which are part of a latent trait which comprises neurocognitive impairments and schizophrenia symptoms. J. Affect. Disord. 2021, 287, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Li, X.; Hie, G.; Yuan, X.; Lü, L.; Song, X. Analysis of the changes of serum high mobility group protein B1 and cytokines in first-episode schizophrenia patients. Zhonghua Yi Xue Za Zhi 2015, 95, 3818–3822. [Google Scholar]
- Chen, S.; Gou, M.; Chen, W.; Xiu, M.; Fan, H.; Tan, Y.; Tian, L. Alterations in innate immune defense distinguish first-episode schizophrenia patients from healthy controls. Front. Psychiatry 2022, 13, 1024299. [Google Scholar] [CrossRef] [PubMed]
- Almulla, A.A.-R.K.; Maes, M.; Kadhem Al-Hakeim, H. In Schizophrenia, Depression and Anxiety Symptoms Are Driven by Immune-Inflammatory Pathways. Preprints.org 2020, 2020120783. [Google Scholar] [CrossRef]
- Al-Hakeim, H.K.M.R.F.; Al-Dujaili, A.H.; Maes, M. Pathway-Phenotypes of Non-responders and Partial Responders to Treatment With Antipsychotics in Schizophrenia: A Machine Learning Study. Preprints.org 2020, 2020040231. [Google Scholar] [CrossRef]
- Ershova, E.S.; Shmarina, G.V.; Porokhovnik, L.N.; Zakharova, N.V.; Kostyuk, G.P.; Umriukhin, P.E.; Kutsev, S.I.; Sergeeva, V.A.; Veiko, N.N.; Kostyuk, S.V. In Vitro Analysis of Biological Activity of Circulating Cell-Free DNA Isolated from Blood Plasma of Schizophrenic Patients and Healthy Controls. Genes 2022, 13, 551. [Google Scholar] [CrossRef]
- Ganapathiraju, M.K.; Thahir, M.; Handen, A.; Sarkar, S.N.; Sweet, R.A.; Nimgaonkar, V.L.; Loscher, C.E.; Bauer, E.M.; Chaparala, S. Schizophrenia interactome with 504 novel protein–protein interactions. Schizophrenia 2016, 2, 16012. [Google Scholar] [CrossRef]
- Zhang, H.; Ding, L.; Shen, T.; Peng, D. HMGB1 involved in stress-induced depression and its neuroinflammatory priming role: A systematic review. Gen. Psychiatry 2019, 32, e100084. [Google Scholar] [CrossRef]
- Franklin, T.C.; Xu, C.; Duman, R.S. Depression and sterile inflammation: Essential role of danger associated molecular patterns. Brain Behav. Immun. 2018, 72, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhao, Z.; Lu, L.; Liu, J.; Wu, X.; Sun, J.; Wei, Y.; Dong, J. The role of HMGB1 in neuroinflammation and tissue repair: A potential therapeutic target for depression? Tradit. Med. Mod. Med. 2018, 1, 85–93. [Google Scholar] [CrossRef]
- Das, U.N. Vagus Nerve Stimulation, Depression, and Inflammation. Neuropsychopharmacology 2007, 32, 2053–2054. [Google Scholar] [CrossRef] [PubMed]
- Lian, Y.-J.; Gong, H.; Wu, T.-Y.; Su, W.-J.; Zhang, Y.; Yang, Y.-Y.; Peng, W.; Zhang, T.; Zhou, J.-R.; Jiang, C.-L.; et al. Ds-HMGB1 and fr-HMGB induce depressive behavior through neuroinflammation in contrast to nonoxid-HMGB1. Brain Behav. Immun. 2017, 59, 322–332. [Google Scholar] [CrossRef]
- Yan, W.; Xie, L.; Bi, Y.; Zeng, T.; Zhao, D.; Lai, Y.; Gao, T.; Sun, X.; Shi, Y.; Dong, Z.; et al. Combined rs-fMRI study on brain functional imaging and mechanism of RAGE-DAMPs of depression: Evidence from MDD patients to chronic stress-induced depression models in cynomolgus monkeys and mice. Clin. Transl. Med. 2021, 11, e541. [Google Scholar] [CrossRef]
- Chen, L.; Jiang, H.; Bao, T.; Wang, Y.; Meng, H.; Sun, Y.; Liu, P.; Quan, S.; Li, W.; Qi, S.; et al. Acupuncture Ameliorates Depressive Behaviors by Modulating the Expression of Hippocampal Iba-1 and HMGB1 in Rats Exposed to Chronic Restraint Stress. Front. Psychiatry 2022, 13, 903004. [Google Scholar] [CrossRef]
- Rana, T.; Behl, T.; Mehta, V.; Uddin, S.; Bungau, S. Molecular insights into the therapeutic promise of targeting HMGB1 in depression. Pharmacol. Rep. 2021, 73, 31–42. [Google Scholar] [CrossRef]
- Caso, J.R.; MacDowell, K.S.; González-Pinto, A.; García, S.; de Diego-Adeliño, J.; Carceller-Sindreu, M.; Sarramea, F.; Caballero-Villarraso, J.; Gracia-García, P.; De la Cámara, C.; et al. Gut microbiota, innate immune pathways, and inflammatory control mechanisms in patients with major depressive disorder. Transl. Psychiatry 2021, 11, 645. [Google Scholar] [CrossRef]
- Hisaoka-Nakashima, K.; Azuma, H.; Ishikawa, F.; Nakamura, Y.; Wang, D.; Liu, K.; Wake, H.; Nishibori, M.; Nakata, Y.; Morioka, N. Corticosterone Induces HMGB1 Release in Primary Cultured Rat Cortical Astrocytes: Involvement of Pannexin-1 and P2X7 Receptor-Dependent Mechanisms. Cells 2020, 9, 1068. [Google Scholar] [CrossRef]
- Xu, X.; Lu, Y.-N.; Cheng, J.-H.; Lan, H.-W.; Lu, J.-M.; Jin, G.-N.; Xu, G.-H.; Jin, C.-H.; Ma, J.; Piao, H.-N.; et al. Ginsenoside Rh2 reduces depression in offspring of mice with maternal toxoplasma infection during pregnancy by inhibiting microglial activation via the HMGB1/TLR4/NF-kappaB signaling pathway. J. Ginseng Res. 2021, 46, 62–70. [Google Scholar] [CrossRef]
- Xie, J.; Bi, B.; Qin, Y.; Dong, W.; Zhong, J.; Li, M.; Cheng, Y.; Xu, J.; Wang, H. Inhibition of phosphodiesterase-4 suppresses HMGB1/RAGE signaling pathway and NLRP3 inflammasome activation in mice exposed to chronic unpredictable mild stress. Brain Behav. Immun. 2021, 92, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Lian, Y.-J.; Su, W.-J.; Peng, W.; Dong, X.; Liu, L.-L.; Gong, H.; Zhang, T.; Jiang, C.-L.; Wang, Y.-X. HMGB1 mediates depressive behavior induced by chronic stress through activating the kynurenine pathway. Brain Behav. Immun. 2018, 72, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Feng, T.; Tripathi, A.; Pillai, A. Inflammatory Pathways in Psychiatric Disorders: The Case of Schizophrenia and Depression. Curr. Behav. Neurosci. Rep. 2020, 7, 128–138. [Google Scholar] [CrossRef]
- Guo, Z.; Zhao, F.; Wang, Y.; Geng, M.; Zhang, Y.; Ma, Q.; Xu, X. Sevoflurane Exerts an Anti-depressive Action by Blocking the HMGB1/TLR4 Pathway in Unpredictable Chronic Mild Stress Rats. J. Mol. Neurosci. 2019, 69, 546–556. [Google Scholar] [CrossRef]
- Miao, L.; Huang, F.; Jiang, W.; Sun, Y.; Chen, Y.; Zhang, M. Curcumin Ameliorates Chronic Mild Stress-Induced Depressive-Like Behavior via HMGB1/TLR4/NF-κB Signaling Pathway. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Xu, X.; Zeng, X.-Y.; Cui, Y.-X.; Li, Y.-B.; Cheng, J.-H.; Zhao, X.-D.; Xu, G.-H.; Ma, J.; Piao, H.-N.; Jin, X.; et al. Antidepressive Effect of Arctiin by Attenuating Neuroinflammation via HMGB1/TLR4- and TNF-α/TNFR1-Mediated NF-κB Activation. ACS Chem. Neurosci. 2020, 11, 2214–2230. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Piao, H.; Aosai, F.; Zeng, X.; Cheng, J.; Cui, Y.; Li, J.; Ma, J.; Jin, X.; Piao, L. Arctigenin protects against depression by inhibiting microglial activation and neuroinflammation via HMGB1/TLR4/NF-κB and TNF-α/TNFR1/NF-κB pathways. Br. J. Pharmacol. 2020, 177, 5224–5245. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhao, L.; Wang, Y.; Guo, Q.; An, Q.; Geng, J.; Zhang, C.; Guo, Z. Ketamine Regulates the Autophagy Flux and Polarization of Microglia through the HMGB1-RAGE Axis and Exerts Antidepressant Effects in Mice. J. Neuropathol. Exp. Neurol. 2022, 81, 931–942. [Google Scholar] [CrossRef]
- Costa-Ferro, Z.S.M.; Prado-Lima, P.A.S.D.; Onsten, G.A.; Oliveira, G.N.; Brito, G.C.; Ghilardi, I.M.; dos Santos, P.G.; Bertinatto, R.J.; da Silva, D.V.; Salamoni, S.D.; et al. Bone marrow mononuclear cell transplant prevents rat depression and modulates inflammatory and neurogenic molecules. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 113, 110455. [Google Scholar] [CrossRef]
- Deng, Z.; Liu, J.; He, S.; Gao, W. The Pyroptosis-Related Signature Predicts Diagnosis and Indicates Immune Characteristic in Major Depressive Disorder. Front. Pharmacol. 2022, 13, 848939. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Huang, X.; Pan, X.; Zhang, T.; Hou, C.; Su, W.-J.; Liu, L.-L.; Li, J.-M.; Wang, Y.-X. Minocycline prevents the depressive-like behavior through inhibiting the release of HMGB1 from microglia and neurons. Brain Behav. Immun. 2020, 88, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Hisaoka-Nakashima, K.; Tomimura, Y.; Yoshii, T.; Ohata, K.; Takada, N.; Zhang, F.F.; Nakamura, Y.; Liu, K.; Wake, H.; Nishibori, M.; et al. High-mobility group box 1-mediated microglial activation induces anxiodepressive-like behaviors in mice with neuropathic pain. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 92, 347–362. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhu, W.; Cai, X.; Zhang, W.; Yu, Z.; Li, X.; Zhang, J.; Cai, M.; Xiang, J.; Cai, D. Minocycline alleviates NLRP3 inflammasome-dependent pyroptosis in monosodium glutamate-induced depressive rats. Biochem. Biophys. Res. Commun. 2020, 526, 553–559. [Google Scholar] [CrossRef]
- Velásquez, E.; Martins-De-Souza, D.; Velásquez, I.; Carneiro, G.R.A.; Schmitt, A.; Falkai, P.; Domont, G.B.; Nogueira, F.C.S. Quantitative Subcellular Proteomics of the Orbitofrontal Cortex of Schizophrenia Patients. J. Proteome Res. 2019, 18, 4240–4253. [Google Scholar] [CrossRef]
- Wu, T.-Y.; Liu, L.; Zhang, W.; Zhang, Y.; Liu, Y.-Z.; Shen, X.-L.; Gong, H.; Yang, Y.-Y.; Bi, X.-Y.; Jiang, C.-L.; et al. High-mobility group box-1 was released actively and involved in LPS induced depressive-like behavior. J. Psychiatr. Res. 2015, 64, 99–106. [Google Scholar] [CrossRef]
- Franklin, T.C.; Wohleb, E.S.; Zhang, Y.; Fogaça, M.; Hare, B.; Duman, R.S. Persistent Increase in Microglial RAGE Contributes to Chronic Stress–Induced Priming of Depressive-like Behavior. Biol. Psychiatry 2018, 83, 50–60. [Google Scholar] [CrossRef]
- Jiang, Q.; Li, C.; Zeng, W.; Xu, H.; Li, J.; Zhang, T.; Deng, G.; Wang, Y. Inhibition of Connexin 36 attenuates HMGB1-mediated depressive-like behaviors induced by chronic unpredictable mild stress. Brain Behav. 2022, 12, e2470. [Google Scholar] [CrossRef]
- Ghosh, D.; Singh, A.; Kumar, A.; Sinha, N. High mobility group box 1 (HMGB1) inhibition attenuates lipopolysaccharide-induced cognitive dysfunction and sickness-like behavior in mice. Immunol. Res. 2022, 70, 633–643. [Google Scholar] [CrossRef]
- Wang, B.; Lian, Y.-J.; Dong, X.; Peng, W.; Liu, L.-L.; Su, W.-J.; Gong, H.; Zhang, T.; Jiang, C.-L.; Li, J.-S.; et al. Glycyrrhizic acid ameliorates the kynurenine pathway in association with its antidepressant effect. Behav. Brain Res. 2018, 353, 250–257. [Google Scholar] [CrossRef]
- Paudel, Y.N.; Angelopoulou, E.; Semple, B.; Piperi, C.; Othman, I.; Shaikh, M.F. Potential Neuroprotective Effect of the HMGB1 Inhibitor Glycyrrhizin in Neurological Disorders. ACS Chem. Neurosci. 2020, 11, 485–500. [Google Scholar] [CrossRef] [PubMed]
- Bian, H.; Xiao, L.; Liang, L.; Xie, Y.; Wang, H.; Slevin, M.; Tu, W.-J.; Wang, G. Polydatin Prevents Neuroinflammation and Relieves Depression via Regulating Sirt1/HMGB1/NF-κB Signaling in Mice. Neurotox. Res. 2022, 40, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Lian, Y.; Su, W.; Liu, L.; Li, J.; Jiang, C.; Wang, Y. Fr-HMGB1 and ds-HMGB1 activate the kynurenine pathway via different mechanisms in association with depressive-like behavior. Mol. Med. Rep. 2019, 20, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Peters, E.M.J.; Neusetzer, M.; Akinci, S.; Murat, A.; Treuherz, S.; Rose, M.; Leweke, F.; Leichsenring, F.; Conrad, M.L.; Kruse, J. Multimodal Psychotherapeutic Inpatient Therapy of Depression Is Successful in Patients With High Cytokine Production. Front. Psychiatry 2020, 11, 571636. [Google Scholar] [CrossRef]
- Marie-Claire, C.; Courtin, C.; Curis, E.; Bouaziz-Amar, E.; Laplanche, J.-L.; Jacob, A.; Etain, B.; Blanchard, A.; Bellivier, F. Increased plasma levels of high mobility group box 1 protein in patients with bipolar disorder: A pilot study. J. Neuroimmunol. 2019, 334, 576993. [Google Scholar] [CrossRef]
- Maes, M.; Carvalho, A.F. The Compensatory Immune-Regulatory Reflex System (CIRS) in Depression and Bipolar Disorder. Mol. Neurobiol. 2018, 55, 8885–8903. [Google Scholar] [CrossRef]
- Fries, G.R.; Walss-Bass, C.; Bauer, M.E.; Teixeira, A.L. Revisiting inflammation in bipolar disorder. Pharmacol. Biochem. Behav. 2019, 177, 12–19. [Google Scholar] [CrossRef]
- Yasuhara, D.; Hashiguchi, T.; Kawahara, K.; Nakahara, T.; Harada, T.; Taguchi, H.; Yamada, S.; Maruyama, I.; Inui, A. High mobility group box 1 and refeeding-resistance in anorexia nervosa. Mol. Psychiatry 2007, 12, 976–977. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, Z.; Lu, L.; Liu, J.; Sun, J.; Dong, J. Icariin and icaritin ameliorated hippocampus neuroinflammation via mediating HMGB1 expression in social defeat model in mice. Int. Immunopharmacol. 2019, 75, 105799. [Google Scholar] [CrossRef]
- Zolfaghari, F.S.; Pirri, F.; Gauvin, E.; Peeri, M.; Amiri, S. Exercise and fluoxetine treatment during adolescence protect against early life stress-induced behavioral abnormalities in adult rats. Pharmacol. Biochem. Behav. 2021, 205, 173190. [Google Scholar] [CrossRef]
- Di Salvo, E.; Casciaro, M.; Quartuccio, S.; Genovese, L.; Gangemi, S. Do Alarmins Have a Potential Role in Autism Spectrum Disorders Pathogenesis and Progression? Biomolecules 2018, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.; Zheng, Y.; Xu, F.; Han, X.; Zhao, H. HMGB1 in nervous system diseases: A common biomarker and potential therapeutic target. Front. Neurol. 2022, 13, 1029891. [Google Scholar] [CrossRef] [PubMed]
- Chirico, V.; Lacquaniti, A.; Salpietro, V.; Munafò, C.; Calabrò, M.P.; Buemi, M.; Arrigo, T.; Salpietro, C. High-mobility group box 1 (HMGB1) in childhood: From bench to bedside. Eur. J. Pediatr. 2014, 173, 1123–1136. [Google Scholar] [CrossRef]
- Makris, G.; Chouliaras, G.; Apostolakou, F.; Papageorgiou, C.; Chrousos, G.P.; Papassotiriou, I.; Pervanidou, P. Increased Serum Concentrations of High Mobility Group Box 1 (HMGB1) Protein in Children with Autism Spectrum Disorder. Children 2021, 8, 478. [Google Scholar] [CrossRef]
- Carissimi, C.; Laudadio, I.; Palone, F.; Fulci, V.; Cesi, V.; Cardona, F.; Alfonsi, C.; Cucchiara, S.; Isoldi, S.; Stronati, L. Functional analysis of gut microbiota and immunoinflammation in children with autism spectrum disorders. Dig. Liver Dis. 2019, 51, 1366–1374. [Google Scholar] [CrossRef]
- Russo, A.J. Decreased Epidermal Growth Factor (EGF) Associated with HMGB1 and Increased Hyperactivity in Children with Autism. Biomark. Insights 2013, 8, 35–41. [Google Scholar] [CrossRef]
- Emanuele, E.; Boso, M.; Brondino, N.; Pietra, S.; Barale, F.; di Nemi, S.U.; Politi, P. Increased serum levels of high mobility group box 1 protein in patients with autistic disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 681–683. [Google Scholar] [CrossRef]
- Meyer, U.; Feldon, J.; Dammann, O. Schizophrenia and Autism: Both Shared and Disorder-Specific Pathogenesis Via Perinatal Inflammation? Pediatr. Res. 2011, 69, 26R–33R. [Google Scholar] [CrossRef] [PubMed]
- Russo1, A.J.; Mensah, A.; Bowman, J. Increased Hmgb1 Associated With Low Zinc and Symptom Severity in Children with Autism. Adv. Neurol. Neurosci. 2020, 3, 45–48. [Google Scholar]
- Babinská, K.; Bucová, M.; Ďurmanová, V.; Lakatošová, S.; Jánošíková, D.; Bakos, J.; Hlavatá, A.; Ostatníková, D. Increased Plasma Levels of the High Mobility Group Box 1 Protein (HMGB1) Are Associated With a Higher Score of Gastrointestinal Dysfunction in Individuals With Autism. Physiol. Res. 2014, 63, S613–S618. [Google Scholar] [CrossRef]
- Dipasquale, V.; Cutrupi, M.C.; Colavita, L.; Manti, S.; Cuppari, C.; Salpietro, C. Neuroinflammation in Autism Spectrum Disorders: The Role of High Mobility Group Box 1 Protein. Int. J. Mol. Cell. Med. 2017, 6, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.J. Increased Epidermal Growth Factor Receptor (EGFR) Associated with Hepatocyte Growth Factor (HGF) and Symptom Severity in Children with Autism Spectrum Disorders (ASDs). J. Central Nerv. Syst. Dis. 2014, 6, 79–83. [Google Scholar] [CrossRef]
- Haghani, A.; Johnson, R.G.; Woodward, N.C.; Feinberg, J.I.; Lewis, K.; Ladd-Acosta, C.; Safi, N.; Jaffe, A.E.; Sioutas, C.; Allayee, H.; et al. Adult mouse hippocampal transcriptome changes associated with long-term behavioral and metabolic effects of gestational air pollution toxicity. Transl. Psychiatry 2020, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Sun, L.; Wang, T.; Liu, C.; Zhang, H.; Zhang, C.; Yu, L. Gestational exposure to PM2.5 leads to cognitive dysfunction in mice offspring via promoting HMGB1-NLRP3 axis mediated hippocampal inflammation. Ecotoxicol. Environ. Saf. 2021, 223, 112617. [Google Scholar] [CrossRef]
- Rauvala, H.; Rouhiainen, A. RAGE as a receptor of HMGB1 (Amphoterin): Roles in health and disease. Curr. Mol. Med. 2007, 7, 725–734. [Google Scholar] [CrossRef]
- Crews, F.T.; Vetreno, R.P. Mechanisms of neuroimmune gene induction in alcoholism. Psychopharmacology 2016, 233, 1543–1557. [Google Scholar] [CrossRef] [PubMed]
- Orio, L.; Antón, M.; Rodríguez-Rojo, I.C.; Correas, Á.; García-Bueno, B.; Corral, M.; de Fonseca, F.R.; García-Moreno, L.M.; Maestú, F.; Cadaveira, F. Young alcohol binge drinkers have elevated blood endotoxin, peripheral inflammation and low cortisol levels: Neuropsychological correlations in women. Addict. Biol. 2017, 23, 1130–1144. [Google Scholar] [CrossRef]
- Vannier, A.G.; Wardwell, B.; Fomin, V.; PeBenito, A.; Wolczynski, N.; Piaker, S.; Kedrin, D.; Chung, R.T.; Schaefer, E.; Goodman, R.; et al. Serum HMGB1 associates with liver disease and predicts readmission and mortality in patients with alcohol use disorder. Alcohol 2021, 95, 37–43. [Google Scholar] [CrossRef]
- Shang, Y.; Jiang, M.; Chen, N.; Jiang, X.-L.; Zhan, Z.-Y.; Zhang, Z.-H.; Zuo, R.-M.; Wang, H.; Lan, X.-Q.; Ren, J.; et al. Inhibition of HMGB1/TLR4 Signaling Pathway by Digitoflavone: A Potential Therapeutic Role in Alcohol-Associated Liver Disease. J. Agric. Food Chem. 2022, 70, 2968–2983. [Google Scholar] [CrossRef]
- Gangemi, S.; Casciaro, M.; Trapani, G.; Quartuccio, S.; Navarra, M.; Pioggia, G.; Imbalzano, E. Association between HMGB1 and COPD: A Systematic Review. Mediat. Inflamm. 2015, 2015, 164913. [Google Scholar] [CrossRef]
- Ma, Y.; Long, Y.; Chen, Y. Roles of Inflammasome in Cigarette Smoke-Related Diseases and Physiopathological Disorders: Mechanisms and Therapeutic Opportunities. Front. Immunol. 2021, 12, 7220049. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Li, J.; Song, Q.; Cheng, W.; Chen, P. The role of HMGB1/RAGE/TLR4 signaling pathways in cigarette smoke-induced inflammation in chronic obstructive pulmonary disease. Immun. Inflamm. Dis. 2022, 10, e711. [Google Scholar] [CrossRef]
- Chen, L.; Sun, X.; Zhong, X. Role of RAGE and its ligand HMGB1 in the development of COPD. Postgrad. Med. 2022, 134, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Le, Y.; Wang, Y.; Zhou, L.; Xiong, J.; Tian, J.; Yang, X.; Gai, X.; Sun, Y. Cigarette smoke-induced HMGB1 translocation and release contribute to migration and NF-κB activation through inducing autophagy in lung macrophages. J. Cell. Mol. Med. 2020, 24, 1319–1331. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-L.; Chien, M.-H.; Chou, Y.-E.; Chang, J.-H.; Liu, T.-C.; Tsao, T.C.-Y.; Chou, M.-C.; Yang, S.-F. Association of EGFR mutations and HMGB1 genetic polymorphisms in lung adenocarcinoma patients. J. Cancer 2019, 10, 2907–2914. [Google Scholar] [CrossRef]
- El Tabaa, M.M.; Anis, A.; Elgharabawy, R.M.; El-Borai, N.B. GLP-1 mediates the neuroprotective action of crocin against cigarette smoking-induced cognitive disorders via suppressing HMGB1-RAGE/TLR4-NF-κB pathway. Int. Immunopharmacol. 2022, 110, 108995. [Google Scholar] [CrossRef]
- Mohammadi, L.; Han, D.D.; Xu, F.; Huang, A.; Derakhshandeh, R.; Rao, P.; Whitlatch, A.; Cheng, J.; Keith, R.J.; Hamburg, N.M.; et al. Chronic E-Cigarette Use Impairs Endothelial Function on the Physiological and Cellular Levels. Arter. Thromb. Vasc. Biol. 2022, 42, 1333–1350. [Google Scholar] [CrossRef]
- Lee, H.S.; Park, H.-W. Role of mTOR in the Development of Asthma in Mice With Cigarette Smoke-Induced Cellular Senescence. J. Gerontol. A Biol. Sci. Med. Sci. 2022, 77, 433–442. [Google Scholar] [CrossRef]
- Gao, S.; Zhang, H.; He, J.; Zheng, H.; Zhang, P.; Xu, J.; Shen, Z.; Zhao, H.; Wang, F.; Hu, Z.; et al. Neuronal HMGB1 in nucleus accumbens regulates cocaine reward memory. Addict. Biol. 2020, 25, e12739. [Google Scholar] [CrossRef]
- Frank, M.G.; Adhikary, S.; Sobesky, J.L.; Weber, M.D.; Watkins, L.R.; Maier, S.F. The danger-associated molecular pattern HMGB1 mediates the neuroinflammatory effects of methamphetamine. Brain Behav. Immun. 2016, 51, 99–108. [Google Scholar] [CrossRef]
- Masai, K.; Kuroda, K.; Isooka, N.; Kikuoka, R.; Murakami, S.; Kamimai, S.; Wang, D.; Liu, K.; Miyazaki, I.; Nishibori, M.; et al. Neuroprotective Effects of Anti-high Mobility Group Box-1 Monoclonal Antibody Against Methamphetamine-Induced Dopaminergic Neurotoxicity. Neurotox. Res. 2021, 39, 1511–1523. [Google Scholar] [CrossRef] [PubMed]
- Alabed, S.; Zhou, H.; Sariyer, I.K.; Chang, S.L. Meta-Analysis of Methamphetamine Modulation on Amyloid Precursor Protein through HMGB1 in Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 4781. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, T.; Zhang, X.; Chao, J.; Hu, G.; Yao, H. Role of high-mobility group box 1 in methamphetamine-induced activation and migration of astrocytes. J. Neuroinflammation 2015, 12, 156. [Google Scholar] [CrossRef] [PubMed]
- Plaitakis, A.; Kalef-Ezra, E.; Kotzamani, D.; Zaganas, I.; Spanaki, C. The Glutamate Dehydrogenase Pathway and Its Roles in Cell and Tissue Biology in Health and Disease. Biology 2017, 6, 11. [Google Scholar] [CrossRef]
- Kravos, M. Glutamate Dehydrogenase Applicability in Clinical Practice. Med. Res. Arch. 2021, 9. [Google Scholar] [CrossRef]
- Karaca, M.; Martin-Levilain, J.; Grimaldi, M.; Li, L.; Dizin, E.; Emre, Y.; Maechler, P. Liver Glutamate Dehydrogenase Controls Whole-Body Energy Partitioning Through Amino Acid–Derived Gluconeogenesis and Ammonia Homeostasis. Diabetes 2018, 67, 1949–1961. [Google Scholar] [CrossRef]
- Voss, C.M.; Arildsen, L.; Nissen, J.D.; Waagepetersen, H.S.; Schousboe, A.; Maechler, P.; Ott, P.; Vilstrup, H.; Walls, A.B. Glutamate Dehydrogenase Is Important for Ammonia Fixation and Amino Acid Homeostasis in Brain During Hyperammonemia. Front. Neurosci. 2021, 15, 646291. [Google Scholar] [CrossRef]
- Frieden, C. Glutamate Dehydrogenase. J. Biol. Chem. 1965, 240, 2028–2035. [Google Scholar] [CrossRef]
- Desai, M.; Knight, K.; Gray, J.M.; Nguyen, V.; Boone, J.; Sorrentino, D. Low glutamate dehydrogenase levels are associated with colonization in Clostridium difficile PCR-only positive patients with inflammatory bowel disease. Eur. J. Gastroenterol. Hepatol. 2020, 32, 1099–1105. [Google Scholar] [CrossRef]
- Viazis, N.; Pontas, C.; Karmiris, K.; Dimas, I.; Fragaki, M.; Paspatis, G.; Drygiannakis, I.; Koutroubakis, I.E.; Moschovis, D.; Tzouvala, M.; et al. Prevalence of Clostridium difficile infection among hospitalized inflammatory bowel disease patients in Greece. Eur. J. Gastroenterol. Hepatol. 2019, 31, 773–776. [Google Scholar] [CrossRef]
- McGill, M.R.; Jaeschke, H. Biomarkers of mitotoxicity after acute liver injury: Further insights into the interpretation of glutamate dehydrogenase. J. Clin. Transl. Res. 2021, 7, 61–65. [Google Scholar]
- Zhao, Y.; Wang, J.; Zhang, J.; Sun, Z.; Niu, R.; Manthari, R.K.; Ommati, M.M.; Wang, S.; Wang, J. Fluoride exposure induces mitochondrial damage and mitophagy via activation of the IL-17A pathway in hepatocytes. Sci. Total. Environ. 2022, 804, 150184. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Martínez, O.; Méndez, I.; Turrubiate, I.; Valente-Godínez, H.; Pérez-Mendoza, M.; García-Tejada, P.; Díaz-Muñoz, M. Restricted feeding modulates the daily variations of liver glutamate dehydrogenase activity, expression, and histological location. Exp. Biol. Med. 2017, 242, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Teschke, R.; Eickhoff, A.; Brown, A.C.; Neuman, M.G.; Schulze, J. Diagnostic Biomarkers in Liver Injury by Drugs, Herbs, and Alcohol: Tricky Dilemma after EMA Correctly and Officially Retracted Letter of Support. Int. J. Mol. Sci. 2019, 21, 212. [Google Scholar] [CrossRef]
- Umbaugh, D.S.; Jaeschke, H. Biomarkers of drug-induced liver injury: A mechanistic perspective through acetaminophen hepatotoxicity. Expert Rev. Gastroenterol. Hepatol. 2021, 15, 363–375. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Zhu, J.; Li, Y.; Luo, W.; Xiang, H.; Zhang, Q.; Peng, W. Serum biomarkers of isoniazid-induced liver injury: Aminotransferases are insufficient, and OPN, L-FABP and HMGB1 can be promising novel biomarkers. J. Appl. Toxicol. 2022, 42, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Church, R.J.; Schomaker, S.J.; Eaddy, J.S.; Boucher, G.G.; Kreeger, J.M.; Aubrecht, J.; Watkins, P.B. Glutamate dehydrogenase as a biomarker for mitotoxicity; insights from furosemide hepatotoxicity in the mouse. PLoS ONE 2020, 15, e0240562. [Google Scholar] [CrossRef]
- Michalak, S.; Rybacka-Mossakowska, J.; Ambrosius, W.; Gazdulska, J.; Gołda-Gocka, I.; Kozubski, W.; Ramlau, R. The Markers of Glutamate Metabolism in Peripheral Blood Mononuclear Cells and Neurological Complications in Lung Cancer Patients. Dis. Markers 2016, 2016, 2895972. [Google Scholar] [CrossRef]
- Spinelli, J.B.; Yoon, H.; Ringel, A.E.; Jeanfavre, S.; Clish, C.B.; Haigis, M.C. Metabolic recycling of ammonia via glutamate dehydrogenase supports breast cancer biomass. Science 2017, 358, 941–946. [Google Scholar] [CrossRef]
- Jin, L.; Li, D.; Alesi, G.N.; Fan, J.; Kang, H.-B.; Lu, Z.; Boggon, T.J.; Jin, P.; Yi, H.; Wright, E.R.; et al. Glutamate Dehydrogenase 1 Signals through Antioxidant Glutathione Peroxidase 1 to Regulate Redox Homeostasis and Tumor Growth. Cancer Cell 2015, 27, 257–270. [Google Scholar] [CrossRef]
- Savushkina, O.K.; Tereshkina, E.B.; Prokhorova, T.A.; Boksha, I.S.; Burminskii, D.S.; Vorobyeva, E.A.; Morozova, M.A.; Burbaeva, G.S. Platelet glutamate dehydrogenase activity and efficacy of antipsychotic therapy in patients with schizophrenia. J. Med. Biochem. 2020, 39, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Burbaeva, G.S.; Boksha, I.S.; Tereshkina, E.B.; Savushkina, O.K.; Turishcheva, M.S.; Starodubtseva, L.I.; Brusov, O.S.; Morozova, M.A. Effect of olanzapine treatment on platelet glutamine synthetase-like protein and glutamate dehydrogenase immunoreactivity in schizophrenia. World J. Biol. Psychiatry 2006, 7, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Burbaeva, G.S.; Boksha, I.S.; Starodubtseva, I.L.; Savushkina, O.; Tereshkina, E.B.; Turishcheva, M.S.; Prokhorova, T.; Vorob’Eva, E.A.; Morozova, M. [Glutamate dysmetabolism in patients with schizophrenia]. Vestn. Ross. Akad. Med. Nauk. 2007, 3, 19–24. [Google Scholar]
- Plaitakis, A.; Zaganas, I.; Spanaki, C. Deregulation of glutamate dehydrogenase in human neurologic disorders. J. Neurosci. Res. 2013, 91, 1007–1017. [Google Scholar] [CrossRef]
- Lander, S.S.; Chornyy, S.; Safory, H.; Gross, A.; Wolosker, H.; Gaisler-Salomon, I.; Sima, L.S.; Sergiy, C.; Hazem, S.; Amit, G.; et al. Glutamate dehydrogenase deficiency disrupts glutamate homeostasis in hippocampus and prefrontal cortex and impairs recognition memory. Genes Brain Behav. 2020, 19, e12636. [Google Scholar] [CrossRef]
- Duarte, J.M.N.; Xin, L. Magnetic Resonance Spectroscopy in Schizophrenia: Evidence for Glutamatergic Dysfunction and Impaired Energy Metabolism. Neurochem. Res. 2019, 44, 102–116. [Google Scholar] [CrossRef]
- Burbaeva, G.S.; Boksha, I.S.; Tereshkina, E.B.; Savushkina, O.K.; Starodubtseva, L.I.; Turishcheva, M.S.; Mukaetova-Ladinska, E. Systemic Neurochemical Alterations in Schizophrenic Brain: Glutamate Metabolism in Focus. Neurochem. Res. 2007, 32, 1434–1444. [Google Scholar] [CrossRef]
- Burbaeva, G.S.; Boksha, I.S.; Turishcheva, M.S.; Vorobyeva, E.A.; Savushkina, O.K.; Tereshkina, E.B. Glutamine synthetase and glutamate dehydrogenase in the prefrontal cortex of patients with schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2003, 27, 675–680. [Google Scholar] [CrossRef]
- Tereshkina, E.B.; Prokhorova, T.A.; Boksha, I.S.; Savushkina, O.K.; Vorobyeva, E.A.; Burbaeva, G.S. Comparative study of glutamate dehydrogenase in the brain of patients with schizophrenia and mentally healthy people. Zh. Nevrol. Psikhiatr. Im. S. S. Korsakova 2017, 117, 101–107. [Google Scholar] [CrossRef]
- Olga Savushkina, I.B.; Tereshkina, E.; Prokhorova, T.; Vorobyeva, E.; Burbaeva, G. Glutamate metabolizing enzymes in frontal, cingulate and cerebellar cortex: Anomalities revealed in schizophrenia. Psychiatry 2018, 77, 16–25. [Google Scholar] [CrossRef]
- Burbaeva, G.; Turishcheva, M.; Savushkina, O.; Tereshkina, E.; Boksha, I. Increase of glutamate dehydrogenase levels in frontal and cingulate cortex in schizophrenia. Schizophr. Res. 2000, 41, 255. [Google Scholar] [CrossRef]
- Burbaeva, G.; Boksha, I.S.; Turishcheva, M.S.; Savushkina, O.K.; Tereshkina, E.B.; Vorob’eva, E.A. Impaired cerebral glutamate metabolism in mental diseases (Alzheimer’s disease, schizophrenia. Vestn. Ross. Akad. Med. Nauk. 2001, 7, 34–37. [Google Scholar]
- Irina, S.; Boksha, O.K.S.; Tereshkina, E.B.; Prokhorova, T.A.; Elizabeta, B. Mukaetova-Ladinska Enzymes of Glutamate System. Biochem. Approaches Glutamatergic Neurotransmission 2017, 139, 469–506. [Google Scholar]
- Venkataramaiah, C.; Payani, S.; Priya, B.L.; Pradeepkiran, J.A. Therapeutic potentiality of a new flavonoid against ketamine induced glutamatergic dysregulation in schizophrenia: In vivo and in silico approach. Biomed. Pharmacother. 2021, 138, 111453. [Google Scholar] [CrossRef]
- Lander, S.S.; Khan, U.; Lewandowski, N.; Chakraborty, D.; Provenzano, F.A.; Mingote, S.; Chornyy, S.; Frigerio, F.; Maechler, P.; Kaphzan, H.; et al. Glutamate Dehydrogenase–Deficient Mice Display Schizophrenia-Like Behavioral Abnormalities and CA1-Specific Hippocampal Dysfunction. Schizophr. Bull. 2018, 45, 127–137. [Google Scholar] [CrossRef]
- Gluck, M.R.; Thomas, R.G.; Davis, K.L.; Haroutunian, V. Implications for Altered Glutamate and GABA Metabolism in the Dorsolateral Prefrontal Cortex of Aged Schizophrenic Patients. Am. J. Psychiatry 2002, 159, 1165–1173. [Google Scholar] [CrossRef]
- Pochueva, V.; Savushkina, O.; Boksha, I.; Tereshkina, E.; Prokhorova, T.; Sheshenin, V.; Vorobyeva, E.; Burbaeva, G. Late onset schizophrenia and delusional disorder: Activity of platelet energy, glutamate, and glutathione metabolizing enzymes. Eur. Psychiatry 2022, 65, S778–S779. [Google Scholar] [CrossRef]
- Jang, T.-S.; Byoung-Hoon, O.; Jin-Sook, C. Association of Tardive Dyskinesia with Cognitive Deficit in Schizophrenia. Korean J. Biol. Psychiatry 1999, 6, 89–95. [Google Scholar]
- Shimizu, E.; Shirasawa, H.; Kodama, K.; Kuroyanagi, H.; Shirasawa, T.; Sato, T.; Simizu, B. Glutamate dehydrogenase mRNA is immediately induced after phencyclidine treatment in the rat brain. Schizophr. Res. 1997, 25, 251–258. [Google Scholar] [CrossRef]
- Prokhorova, T.A.; Boksha, I.; Savushkina, O.; Tereshkina, E.; Vorobyeva, E.A.; Pomytkin, A.; Kaleda, V.; Burbaeva, G. Glutamate dehydrogenase activity in platelets of patients with endogenous psychosis. Zh. Nevrol. Psikhiatr. Im. S. S. Korsakova 2016, 116, 44–48. [Google Scholar] [CrossRef]
- Boksha, I.S.; Omel’chenko, M.A.; Savushkina, O.K.; Prokhorova, T.A.; Tereshkina, E.B.; Vorobyeva, E.A.; Burbaeva, G.S. Links of platelet glutamate and glutathione metabolism with attenuated positive and negative symptoms in depressed patients at clinical high risk for psychosis. Eur. Arch. Psychiatry Clin. Neurosci 2022, 273, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Savushkina, O.K.; Tereshkina, E.B.; Prokhorova, T.A.; Boksha, I.S.; Safarova, T.P.; Yakovleva, O.B.; Kornilov, V.V.; Shipilova, E.S.; Vorobyeva, E.A.; Burbaeva, G.S. Evaluation of Platelet Glutamate Dehydrogenase Activity in Late-Life Depressions. Psychiatry 2021, 19, 34–41. [Google Scholar] [CrossRef]
- Savushkina, O.; Boksha, I.; Omel’chenko, M.; Tereshkina, E.; Prokhorova, T.; Vorobieva, E.; Burbaeva, G. Activity of enzymes of glutamate, energy and glutathione metabolism in the first juvenile depression with attenuated symptoms of schizophrenia. Zh. Nevrol. Psikhiatr. Im. S. S. Korsakova 2022, 122, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Safarova, T.; Yakovleva, O.; Savushkina, O.; Prokhorova, T. New approaches to the typology and prediction of the effectiveness of therapy for late-life depression. Zh. Nevrol. Psikhiatr. Im. S. S. Korsakova 2022, 122, 36–42. [Google Scholar] [CrossRef]
- El-Ansary, A. Data of multiple regressions analysis between selected biomarkers related to glutamate excitotoxicity and oxidative stress in Saudi autistic patients. Data Brief 2016, 7, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Abu Shmais, G.A.; Al-Ayadhi, L.Y.; Al-Dbass, A.M.; El-Ansary, A.K. Mechanism of nitrogen metabolism-related parameters and enzyme activities in the pathophysiology of autism. J. Neurodev. Disord. 2012, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Shimmura, C.; Suzuki, K.; Iwata, Y.; Tsuchiya, K.J.; Ohno, K.; Matsuzaki, H.; Iwata, K.; Kameno, Y.; Takahashi, T.; Wakuda, T.; et al. Enzymes in the glutamate-glutamine cycle in the anterior cingulate cortex in postmortem brain of subjects with autism. Mol. Autism 2013, 4, 6. [Google Scholar] [CrossRef]
- Gaertner, I.; Altendorf, K.; Batra, A.; Gaertner, H.J. Relevance of Liver Enzyme Elevations With Four Different Neuroleptics: A Retrospective Review of 7,263 Treatment Courses. J. Clin. Psychopharmacol. 2001, 21, 215–222. [Google Scholar] [CrossRef]
- Nam, A.-R.; In-Sik, K.; Seung-Ju, Y. Inhibitory Effects of Human Glutamate Dehydrogenase Isozymes by Antipsychotic Drugs for Schizophrenia. J. Korea Acad. -Ind. Coop. Soc. 2016, 17, 1975–4701. [Google Scholar]
- Jyoti Batra, S.K.; Seth, P.K. Effect of haloperidol administration on GABA and glutamate dehydrogenase activity in Albino Rat Brain. Pharma Innov. J. 2016, 5, 14–16. [Google Scholar]
- Nakamoto, C.; Kawamura, M.; Nakatsukasa, E.; Natsume, R.; Takao, K.; Watanabe, M.; Abe, M.; Takeuchi, T.; Sakimura, K. GluD1 knockout mice with a pure C57BL/6N background show impaired fear memory, social interaction, and enhanced depressive-like behavior. PLoS ONE 2020, 15, e0229288. [Google Scholar] [CrossRef] [PubMed]
- Kravos, M.; Malešič, I. Glutamate Dehydrogenase as a Marker of Alcohol Dependence. Alcohol Alcohol. 2009, 45, 39–44. [Google Scholar] [CrossRef]
- Chemnitz, G.; Schmidt, E.; Schmidt, F.W.; Lobers, J. Diagnostic and prognostic significance of massively increased serum glutamate dehydrogenase activity. Dtsch. Med. Wochenschr. 1984, 109, 1789–1793. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Zannis, V.; Plaitakis, A. Characterization of glutamate dehydrogenase isoproteins purified from the cerebellum of normal subjects and patients with degenerative neurological disorders, and from human neoplastic cell lines. J. Biol. Chem. 1989, 264, 20730–20735. [Google Scholar] [CrossRef] [PubMed]
- Conigrave, K.M.; Davies, P.; Haber, P.; Whitfield, J.B. Traditional markers of excessive alcohol use. Addiction 2003, 98 (Suppl. S2), 31–43. [Google Scholar] [CrossRef]
- Kravos, M.; Malešič, I. Kinetics and isoforms of serum glutamate dehydrogenase in alcoholics. Alcohol Alcohol. 2008, 43, 281–286. [Google Scholar] [CrossRef]
- Worner, T.M.; Lieber, C.S. Plasma Glutamate Dehydrogenase: Clinical Application in Patients With Alcoholic Liver Disease. Alcohol. Clin. Exp. Res. 1980, 4, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Filis, P.; Nagrath, N.; Fraser, M.; Hay, D.C.; Iredale, J.P.; O’Shaughnessy, P.; Fowler, P.A. Maternal Smoking Dysregulates Protein Expression in Second Trimester Human Fetal Livers in a Sex-Specific Manner. J. Clin. Endocrinol. Metab. 2015, 100, E861–E870. [Google Scholar] [CrossRef]
- Zheng, F.; Zhan, C.-G. Modeling of Pharmacokinetics of Cocaine in Human Reveals the Feasibility for Development of Enzyme Therapies for Drugs of Abuse. PLoS Comput. Biol. 2012, 8, e1002610. [Google Scholar] [CrossRef]
- Shi, Y.; Li, Y.; Zhang, J.; Xiao, Y.; Yan, P.; Zhu, Y. GAD1 but not GAD2 polymorphisms are associated with heroin addiction phenotypes. Neurosci. Lett. 2020, 717, 134704. [Google Scholar] [CrossRef]
- Vicente-Rodríguez, M.; Gramage, E.; Herradón, G.; Pérez-García, C. Phosphoproteomic analysis of the striatum from pleiotrophin knockout and midkine knockout mice treated with cocaine reveals regulation of oxidative stress-related proteins potentially underlying cocaine-induced neurotoxicity and neurodegeneration. Toxicology 2013, 314, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Fasakin, O.W.; Oboh, G.; Ademosun, A.O.; Lawal, A.O. The modulatory effects of alkaloid extracts of Cannabis sativa, Datura stramonium, Nicotiana tabacum and male Carica papaya on neurotransmitter, neurotrophic and neuroinflammatory systems linked to anxiety and depression. Inflammopharmacology 2022, 30, 2447–2476. [Google Scholar] [CrossRef] [PubMed]
- Ghoneim, M.T.; Mikhail, M.M.; Mahfouz, M.; Makar, A.B. Effect of hashish smoke on some blood and serum parameters in rabbits. Pharmazie 1980, 35, 226–228. [Google Scholar] [PubMed]
- Ordak, M.; Nasierowski, T.; Muszynska, E. The growing problem of mephedrone use in Warsaw, Poland, 2010–2018. Lancet Psychiatry 2018, 5, 787. [Google Scholar] [CrossRef] [PubMed]
- Ordak, M.; Nasierowski, T.; Muszynska, E. The problem of poly-pharmacotherapy in patients on a mephedrone binge. Pharmacol. Res. 2019, 143, 204. [Google Scholar] [CrossRef] [PubMed]
- Telles-Correia, D.; Barbosa, A.; Cortez-Pinto, H.; Campos, C.; Rocha, N.B.F.; Machado, S. Psychotropic drugs and liver disease: A critical review of pharmacokinetics and liver toxicity. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 26–38. [Google Scholar] [CrossRef]
- Martínez-Vélez, N.A.; Tiburcio, M.; Rey, G.N.; Velázquez, J.A.V.; Arroyo-Belmonte, M.; Sánchez-Hernández, G.Y.; Fernández-Torres, M. Psychoactive Substance Use and Its Relationship to Stress, Emotional State, Depressive Symptomatology, and Perceived Threat During the COVID-19 Pandemic in Mexico. Front. Public Health 2021, 9, 709410. [Google Scholar] [CrossRef]
- Van Hout, M.C.; Benschop, A.; Bujalski, M.; Dąbrowska, K.; Demetrovics, Z.; Felvinczi, K.; Hearne, E.; Henriques, S.; Kaló, Z.; Kamphausen, G.; et al. Health and Social Problems Associated with Recent Novel Psychoactive Substance (NPS) Use Amongst Marginalised, Nightlife and Online Users in Six European Countries. Int. J. Ment. Health Addict. 2018, 16, 480–495. [Google Scholar] [CrossRef]
- Anderson, C.; Morrell, C.; Marchevsky, D. A novel psychoactive substance poses a new challenge in the management of paranoid schizophrenia. BMJ Case Rep. 2015, 2015, bcr201520957. [Google Scholar] [CrossRef]
- Seraj, S.M.; Campbell, E.J.; Argyropoulos, S.K.; Wegermann, K.; Chung, R.T.; Richter, J.M. Hospital readmissions in decompensated cirrhotics: Factors pointing toward a prevention strategy. World J. Gastroenterol. 2017, 23, 6868–6876. [Google Scholar] [CrossRef]
- Holmes, R.; Patel, A.; Desai, A.P. Psychiatric Disorders and Their Treatment: Impact of Outcomes in Patients With Chronic Liver Disease. Clin. Liver Dis. 2022, 20, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Böckmann, V.; Lay, B.; Seifritz, E.; Kawohl, W.; Roser, P.; Habermeyer, B. Patient-Level Predictors of Psychiatric Readmission in Substance Use Disorders. Front. Psychiatry 2019, 10, 828. [Google Scholar] [CrossRef] [PubMed]
- Šprah, L.; Dernovšek, M.Z.; Wahlbeck, K.; Haaramo, P. Psychiatric readmissions and their association with physical comorbidity: A systematic literature review. BMC Psychiatry 2017, 17, 2. [Google Scholar] [CrossRef] [PubMed]
- Kadra, G.; Stewart, R.; Shetty, H.; MacCabe, J.H.; Chang, C.-K.; Kesserwani, J.; Taylor, D.; Hayes, R.D. Antipsychotic polypharmacy prescribing and risk of hospital readmission. Psychopharmacology 2018, 235, 281–289. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Mental Illness | Biomarker | Participants in Study | Results | Reference |
|---|---|---|---|---|
| Schizophrenia | OPN | Study group: 22 patients with schizophrenia Control group: - | Schizophrenic patients under long-term antipsychotics treatment had lower OPN levels than patients under short-term treatment (p = 0.021). | [35] |
| Schizophrenia | HMGB1 | Study group: - 30 schizophrenic patients in the acute exacerbation phase - 29 schizophrenic patients in the remission phase15 healthy controls | HMGB1 levels were higher in study group than in healthy controls, independent of the phase of the disease (acute exacerbation vs. control—p = 0.05; remission vs. control—p = 0.002). | [101] |
| Schizophrenia | HMGB1 | Study group: 115 patients with schizophrenia and 43 healthy controls | HMGB1 levels were higher compared with healthy controls (p < 0.001). | [102] |
| Schizophrenia | GDH | Study group: brain tissues taken at autopsy of 8 schizophrenic patients Control group: brain tissues taken at autopsy of 9 healthy people | Significant differences in GDH levels were observed in the prefrontal cortex (area 10) in schizophrenic patients compared with healthy controls (p < 0.01). | [208] |
| Major depressive Disorder | OPN | Study group: 1172 patients with MDD and 426 healthy controls | OPN levels in patients with MDD were significantly lower compared with healthy controls (p = 0.012). | [38] |
| Major depressive Disorder | OPN | Study group: 28 patients with MDD and 16 healthy controls. | OPN levels in patients with MDD were significantly lower compared with healthy controls. Ketamine significantly increased OPN levels on the 1st day (p < 0.001) and on the 3rd day (p < 0.001) after administration. | [40] |
| Major depressive disorder | GDH | Study group: 78 elderly patients with MDD (including 42 patients before treatment and 36 patients after treatment) and 29 healthy controls | Patients diagnosed with MDD had significant decrease in the activity of GDH compared with healthy controls (p < 0.0008). | [222] |
| Bipolar disorder (BD) | HMGB1 | Study group: 17 patients diagnosed with BD and 16 healthy controls | HMGB1 levels were significantly higher in study group than in healthy controls (p < 0.0001). | [145] |
| Eating disorder | OPN | Study group: 20 patients diagnosed with anorexia nervosa and 78 healthy controls | OPN levels were significantly higher in patients with anorexia compared with healthy controls (p = 0.009). | [48] |
| Eating disorder | HMGB1 | Study group: 11 patients diagnosed with anorexia nervosa (during the observation period and during the refeeding-resistant period) and 11 healthy controls | The average HMGB1 levels in healthy controls and participants during the observation period were significantly lower compared with group during the refeeding-resistant period (p < 0.005). | [148] |
| Autism | OPN | Study group: 42 autistic children Control group: 42 healthy children | The autistic children had significantly higher OPN levels compared with healthy controls (p = 0.02). | [49] |
| Autism | HMGB1 | Study group: 42 autistic children Control group: 38 healthy children | The autistic children had significantly higher HMGB1 levels compared with healthy controls (p = 0.039). | [154] |
| Autism | HMGB1 | Study group: 22 adult patients with autistic disorders and 28 healthy controls | The autistic patients had significantly higher HMGB1 levels than the healthy controls (p < 0.001). | [157] |
| Autism | GDH | Study group: 20 patients with autistic disorders and 20 healthy controls | The GDH activity was lower compared with healthy controls (p = 0.001). | [225] |
| Alcohol dependence | OPN | The retrospective group: 109 participants The prospective group: 95 participants (Included 44 patients with significant liver fibrosis and 65 patients with mild fibrosis.) | The patients with significant liver fibrosis had higher OPN levels compared with the patients with mild fibrosis (p < 0.001). | [53] |
| Alcohol dependence | HMGB1 | Study group: 80 patients with alcohol use disorder and alcohol-associated liver disease | The patients with alcohol use disorder and alcohol-associated liver disease had higher HMGB1 levels compared with patients with alcohol use disorder without alcohol-associated liver disease (p = 0.0002). | [168] |
| Alcohol dependence | GDH | Study group: 238 alcoholic patients (and 141 healthy controls) | GDH levels were significantly higher in alcoholic patients compared with healthy controls (p < 0.0005). | [232] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golub, A.; Ordak, M.; Nasierowski, T.; Bujalska-Zadrozny, M. Advanced Biomarkers of Hepatotoxicity in Psychiatry: A Narrative Review and Recommendations for New Psychoactive Substances. Int. J. Mol. Sci. 2023, 24, 9413. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24119413
Golub A, Ordak M, Nasierowski T, Bujalska-Zadrozny M. Advanced Biomarkers of Hepatotoxicity in Psychiatry: A Narrative Review and Recommendations for New Psychoactive Substances. International Journal of Molecular Sciences. 2023; 24(11):9413. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24119413
Chicago/Turabian StyleGolub, Aniela, Michal Ordak, Tadeusz Nasierowski, and Magdalena Bujalska-Zadrozny. 2023. "Advanced Biomarkers of Hepatotoxicity in Psychiatry: A Narrative Review and Recommendations for New Psychoactive Substances" International Journal of Molecular Sciences 24, no. 11: 9413. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24119413