
Splicing Modulators Are Involved in Human Polyglutamine Diversification via Protein Complexes Shuttling between Nucleus and Cytoplasm

Abstract
:1. Introduction
2. Results
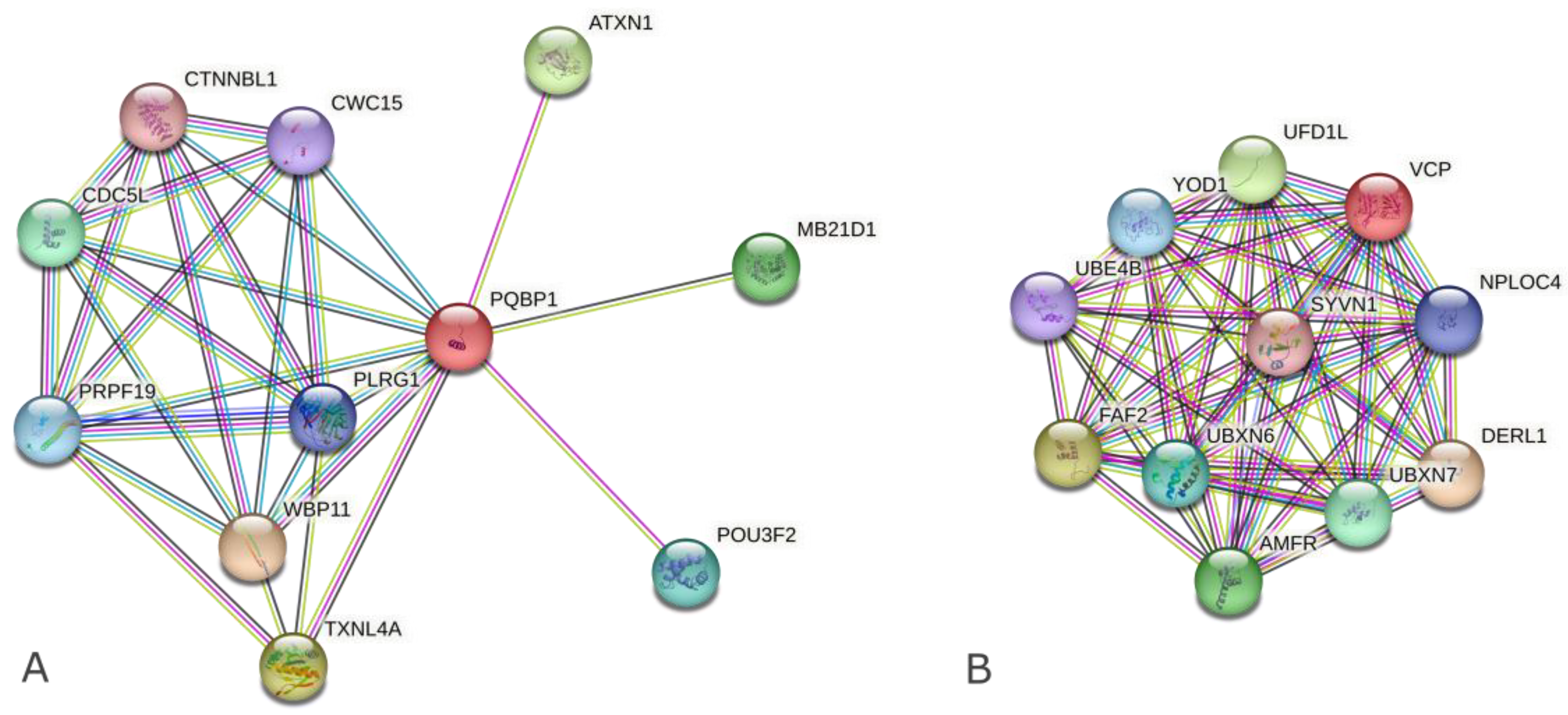
2.1. Search for PPI-Related Information on Complex Formation and Biological Functions Focusing on PolyQ-Binding Ability
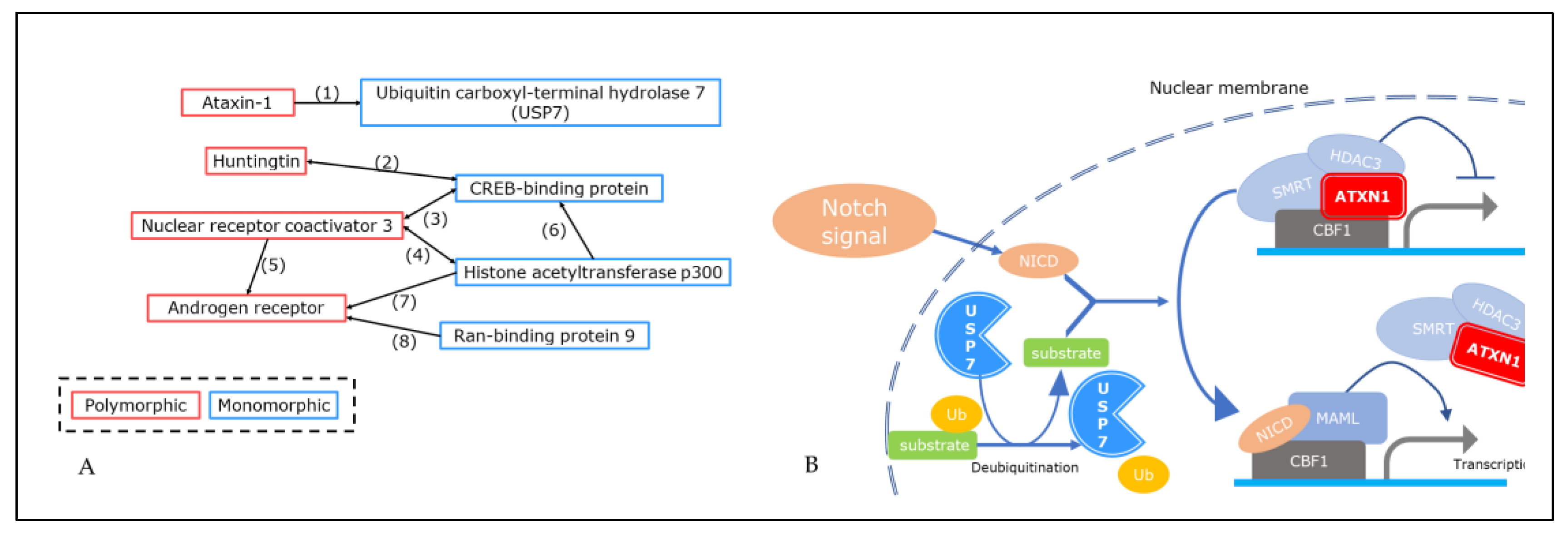
2.2. Search for PPI Information Focusing on Nucleus- and Cytoplasm-Localized Intrinsically Disordered (ID) Hub Proteins
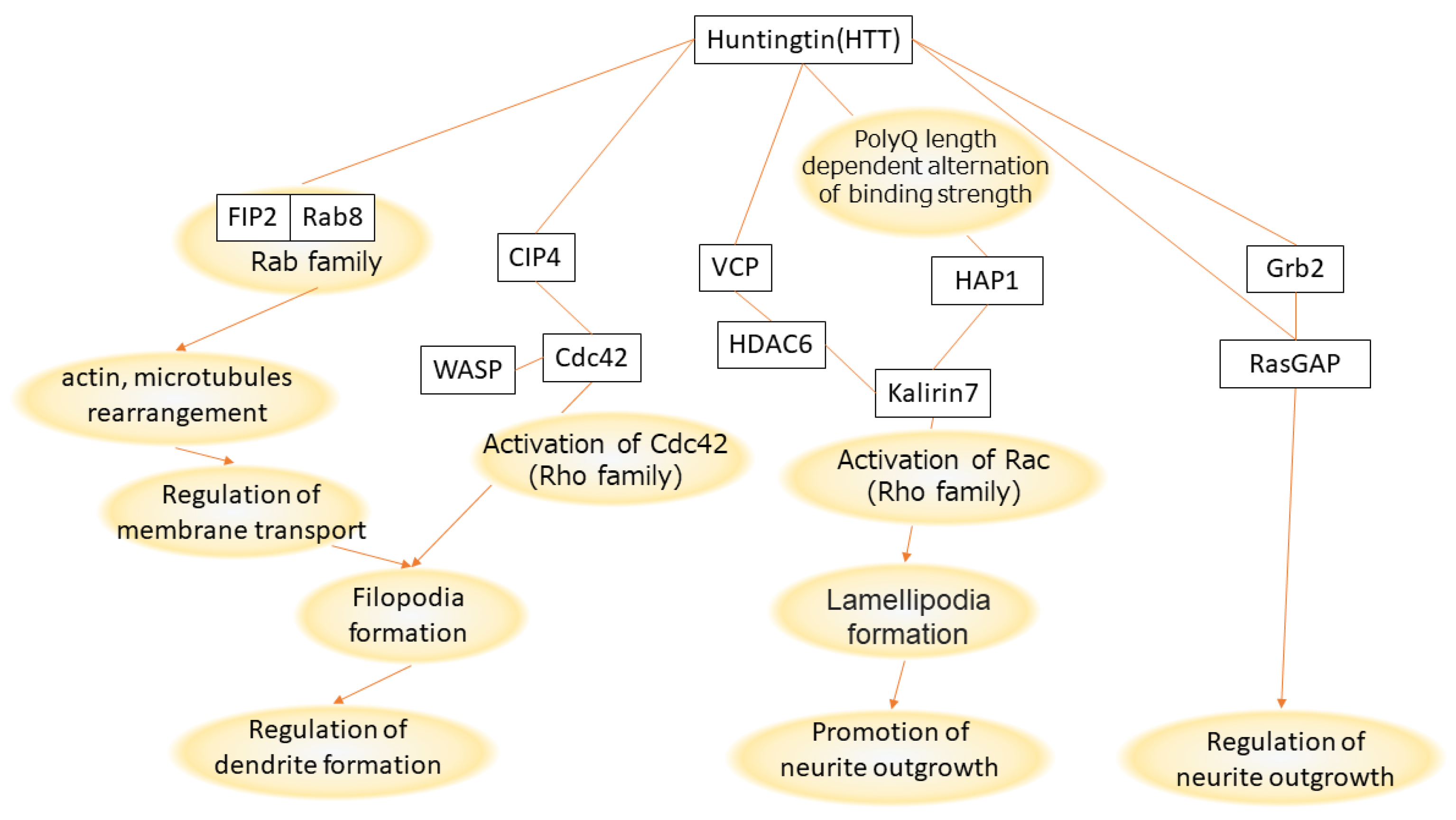
2.3. Review of Signaling Pathways That Explain Effect of Huntingtin (HTT) PolyQ Length on Neurite Length
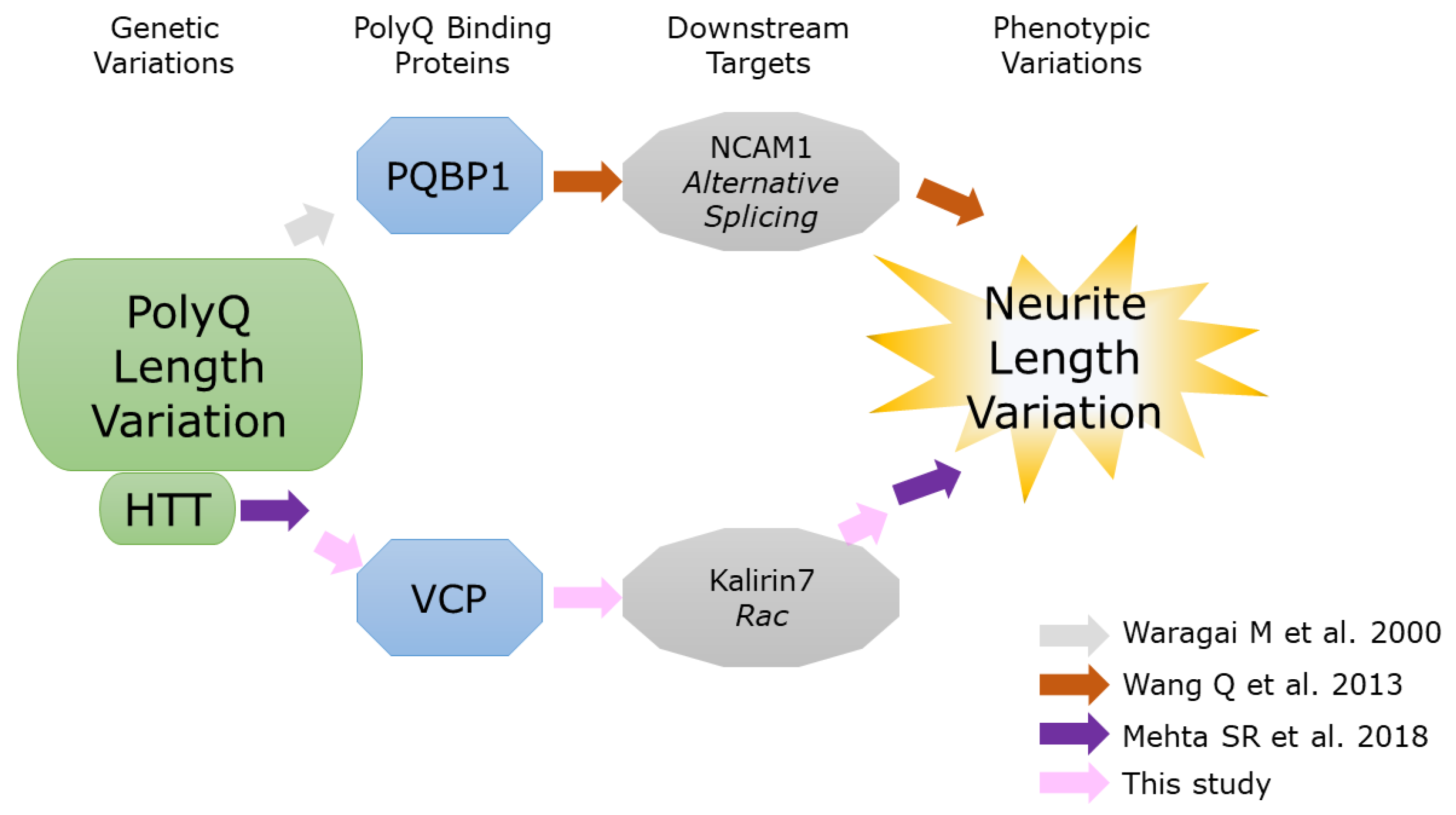
3. Discussion
3.1. PolyQ Repeat Length Variation
3.2. Difference between the Two PolyQ-Binding Proteins
3.3. Suitable Protein Complex
3.4. Traceability of PPI Information
4. Materials and Methods
- (1)
- A search for information on PPIs associated with the complex formation and biological functions focusing on polyQ-binding ability:
- (2)
- A search for information on PPIs focusing on nucleus- and cytoplasm-localized ID hub proteins:
- (3)
- Review of signaling pathways that explain the effect of HTT polyQ length on neurite length:
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shimada, M.K.; Sanbonmatsu, R.; Yamaguchi-Kabata, Y.; Yamasaki, C.; Suzuki, Y.; Chakraborty, R.; Gojobori, T.; Imanishi, T. Selection pressure on human STR loci and its relevance in repeat expansion disease. Mol. Genet. Genom. 2016, 291, 1851–1869. [Google Scholar] [CrossRef]
- Andrés, A.M.; Soldevila, M.; Lao, O.; Volpini, V.; Saitou, N.; Jacobs, H.T.; Hayasaka, I.; Calafell, F.; Bertranpetit, J. Comparative genetics of functional trinucleotide tandem repeats in humans and apes. J. Mol. Evol. 2004, 59, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Ota, M.; Gonja, H.; Koike, R.; Fukuchi, S. Multiple-Localization and Hub Proteins. PLoS ONE 2016, 11, e0156455. [Google Scholar] [CrossRef] [PubMed]
- Tsang, B.; Pritišanac, I.; Scherer, S.W.; Moses, A.M.; Forman-Kay, J.D. Phase Separation as a Missing Mechanism for Interpretation of Disease Mutations. Cell 2020, 183, 1742–1756. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, P.; Chakravarty, D. Intrinsically disordered proteins/regions and insight into their biomolecular interactions. Biophys. Chem. 2022, 283, 106769. [Google Scholar] [CrossRef]
- Horn, J.M.; Zhu, Y.; Ahn, S.Y.; Obermeyer, A.C. Self-assembly of globular proteins with intrinsically disordered protein polyelectrolytes and block copolymers. Soft Matter 2022, 18, 5759–5769. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.E.; Dyson, H.J. Intrinsically disordered proteins in cellular signalling and regulation. Nat. Rev. Mol. Cell Biol. 2015, 16, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Schilling, J.; Broemer, M.; Atanassov, I.; Duernberger, Y.; Vorberg, I.; Dieterich, C.; Dagane, A.; Dittmar, G.; Wanker, E.; van Roon-Mom, W.; et al. Deregulated Splicing Is a Major Mechanism of RNA-Induced Toxicity in Huntington’s Disease. J. Mol. Biol. 2019, 431, 1869–1877. [Google Scholar] [CrossRef]
- Saudou, F.; Humbert, S. The Biology of Huntingtin. Neuron 2016, 89, 910–926. [Google Scholar] [CrossRef]
- Waragai, M.; Lammers, C.-H.; Takeuchi, S.; Imafuku, I.; Udagawa, Y.; Kanazawa, I.; Kawabata, M.; Mouradian, M.M.; Okazawa, H. PQBP-1, a novel polyglutamine tract-binding protein, inhibits transcription activation by Brn-2 and affects cell survival. Hum. Mol. Genet. 1999, 8, 977–987. [Google Scholar] [CrossRef]
- Okazawa, H.; Rich, T.; Chang, A.; Lin, X.; Waragai, M.; Kajikawa, M.; Enokido, Y.; Komuro, A.; Kato, S.; Shibata, M.; et al. Interaction between mutant ataxin-1 and PQBP-1 affects transcription and cell death. Neuron 2002, 34, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Waragai, M.; Junn, E.; Kajikawa, M.; Takeuchi, S.; Kanazawa, I.; Shibata, M.; Mouradian, M.M.; Okazawa, H. PQBP-1/Npw38, a Nuclear Protein Binding to the Polyglutamine Tract, Interacts with U5-15kD/dim1p via the Carboxyl-Terminal Domain. Biochem. Biophys. Res. Commun. 2000, 273, 592–595. [Google Scholar] [CrossRef]
- Mizuguchi, M.; Obita, T.; Serita, T.; Kojima, R.; Nabeshima, Y.; Okazawa, H. Mutations in the PQBP1 gene prevent its interaction with the spliceosomal protein U5-15kD. Nat. Commun. 2014, 5, 3822. [Google Scholar] [CrossRef] [PubMed]
- Alanis-Lobato, G.; Andrade-Navarro, M.A.; Schaefer, M.H. HIPPIE v2.0: Enhancing meaningfulness and reliability of protein–protein interaction networks. Nucleic Acids Res. 2016, 45, D408–D414. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, D607–D613. [Google Scholar] [CrossRef]
- Dennis, G.; Sherman, B.; Hosack, D.; Yang, J.; Gao, W.; Lane, H.; Lempicki, R. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003, 4, R60. [Google Scholar] [CrossRef]
- Hong, S.; Kim, S.-J.; Ka, S.; Choi, I.; Kang, S. USP7, a Ubiquitin-Specific Protease, Interacts with Ataxin-1, the SCA1 Gene Product. Mol. Cell. Neurosci. 2002, 20, 298–306. [Google Scholar] [CrossRef]
- Oates, M.E.; Romero, P.; Ishida, T.; Ghalwash, M.; Mizianty, M.J.; Xue, B.; Dosztányi, Z.; Uversky, V.N.; Obradovic, Z.; Kurgan, L.; et al. D2P2: Database of disordered protein predictions. Nucleic Acids Res. 2012, 41, D508–D516. [Google Scholar] [CrossRef]
- Demarest, S.J.; Martinez-Yamout, M.; Chung, J.; Chen, H.; Xu, W.; Dyson, H.J.; Evans, R.M.; Wright, P.E. Mutual synergistic folding in recruitment of CBP/p300 by p160 nuclear receptor coactivators. Nature 2002, 415, 549–553. [Google Scholar] [CrossRef]
- Lin, C.H.; Hare, B.J.; Wagner, G.; Harrison, S.C.; Maniatis, T.; Fraenkel, E. A small domain of CBP/p300 binds diverse proteins: Solution structure and functional studies. Mol. Cell 2001, 8, 581–590. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, M.; Liu, H.; Guo, H.; Wang, Y.; Cheng, H.; Chen, L. Role of nuclear receptor coactivator 3 (Ncoa3) in pluripotency maintenance. J. Biol. Chem. 2012, 287, 38295–38304. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.R.; Tom, C.M.; Wang, Y.; Bresee, C.; Rushton, D.; Mathkar, P.P.; Tang, J.; Mattis, V.B. Human Huntington’s Disease iPSC-Derived Cortical Neurons Display Altered Transcriptomics, Morphology, and Maturation. Cell Rep. 2018, 25, 1081–1096.e1086. [Google Scholar] [CrossRef] [PubMed]
- Mao, A.H.; Lyle, N.; Pappu, R.V. Describing sequence-ensemble relationships for intrinsically disordered proteins. Biochem. J. 2013, 449, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Mohan, A.; Oldfield, C.J.; Radivojac, P.; Vacic, V.; Cortese, M.S.; Dunker, A.K.; Uversky, V.N. Analysis of Molecular Recognition Features (MoRFs). J. Mol. Biol. 2006, 362, 1043–1059. [Google Scholar] [CrossRef] [PubMed]
- van der Lee, R.; Buljan, M.; Lang, B.; Weatheritt, R.J.; Daughdrill, G.W.; Dunker, A.K.; Fuxreiter, M.; Gough, J.; Gsponer, J.; Jones, D.T.; et al. Classification of Intrinsically Disordered Regions and Proteins. Chem. Rev. 2014, 114, 6589–6631. [Google Scholar] [CrossRef] [PubMed]
- Cornett, J.; Cao, F.; Wang, C.-E.; Ross, C.A.; Bates, G.P.; Li, S.-H.; Li, X.-J. Polyglutamine expansion of huntingtin impairs its nuclear export. Nat. Genet. 2005, 37, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Mühlau, M.; Winkelmann, J.; Rujescu, D.; Giegling, I.; Koutsouleris, N.; Gaser, C.; Arsic, M.; Weindl, A.; Reiser, M.; Meisenzahl, E.M. Variation within the Huntington’s disease gene influences normal brain structure. PLoS ONE 2012, 7, e29809. [Google Scholar] [CrossRef]
- Yue, S.; Serra, H.G.; Zoghbi, H.Y.; Orr, H.T. The spinocerebellar ataxia type 1 protein, ataxin-1, has RNA-binding activity that is inversely affected by the length of its polyglutamine tract. Hum. Mol. Genet. 2001, 10, 25–30. [Google Scholar] [CrossRef]
- Tirabassi, G.; Cignarelli, A.; Perrini, S.; delli Muti, N.; Furlani, G.; Gallo, M.; Pallotti, F.; Paoli, D.; Giorgino, F.; Lombardo, F.; et al. Influence of CAG Repeat Polymorphism on the Targets of Testosterone Action. Int. J. Endocrinol. 2015, 2015, 298107. [Google Scholar] [CrossRef]
- Pan, B.; Li, R.; Chen, Y.; Tang, Q.; Wu, W.; Chen, L.; Lu, C.; Pan, F.; Ding, H.; Xia, Y.; et al. Genetic Association Between Androgen Receptor Gene CAG Repeat Length Polymorphism and Male Infertility: A Meta-Analysis. Medicine 2016, 95, e2878. [Google Scholar] [CrossRef]
- Olsen, N.J.; Benko, A.L.; Kovacs, W.J. Variation in the androgen receptor gene exon 1 CAG repeat correlates with manifestations of autoimmunity in women with lupus. Endocr. Connect. 2014, 3, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Okazawa, H. PQBP1, an intrinsically disordered/denatured protein at the crossroad of intellectual disability and neurodegenerative diseases. Neurochem. Int. 2018, 119, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Moore, M.J.; Adelmant, G.; Marto, J.A.; Silver, P.A. PQBP1, a factor linked to intellectual disability, affects alternative splicing associated with neurite outgrowth. Genes Dev. 2013, 27, 615–626. [Google Scholar] [CrossRef]
- Liu, X.; Dou, L.-X.; Han, J.; Zhang, Z.C. The Renpenning syndrome—Associated protein PQBP1 facilitates the nuclear import of splicing factor TXNL4A through the karyopherin beta2 receptor. J. Biol. Chem. 2020, 295, 4093–4100. [Google Scholar] [CrossRef] [PubMed]
- Barshir, R.; Shwartz, O.; Smoly, I.Y.; Yeger-Lotem, E. Comparative Analysis of Human Tissue Interactomes Reveals Factors Leading to Tissue-Specific Manifestation of Hereditary Diseases. PLoS Comput. Biol. 2014, 10, e1003632. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, M.H.; Fontaine, J.-F.; Vinayagam, A.; Porras, P.; Wanker, E.E.; Andrade-Navarro, M.A. HIPPIE: Integrating Protein Interaction Networks with Experiment Based Quality Scores. PLoS ONE 2012, 7, e31826. [Google Scholar] [CrossRef]
- Fresno, C.; Fernández, E.A. RDAVIDWebService: A versatile R interface to DAVID. Bioinformatics 2013, 29, 2810–2811. [Google Scholar] [CrossRef]
- Consortium, T.U. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2022, 51, D523–D531. [Google Scholar] [CrossRef]
- Yamasaki, C.; Murakami, K.; Takeda, J.; Sato, Y.; Noda, A.; Sakate, R.; Habara, T.; Nakaoka, H.; Todokoro, F.; Matsuya, A.; et al. H-InvDB in 2009: Extended database and data mining resources for human genes and transcripts. Nucleic Acids Res. 2010, 38, D626–D632. [Google Scholar] [CrossRef]
- Goel, R.; Harsha, H.C.; Pandey, A.; Prasad, T.S. Human Protein Reference Database and Human Proteinpedia as resources for phosphoproteome analysis. Mol. Biosyst. 2012, 8, 453–463. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | ||||||||||
| PolyQ-Disease-Causing Proteins (Carriers) | ||||||||||
| HTT | AR | ATXN1 | ATXN3 | ATXN7 | ||||||
| Binding proteins (binders) | PQBP1 | 2 | 4 | 1 | 0 | 0 | ||||
| VCP | 52 | 44 | 25 | 34 | 11 | |||||
| (b) | ||||||||||
| PolyQ-Disease-Causing Proteins (Carriers) | ||||||||||
| HTT | AR | ATN1 | ATXN1 | ATXN2 | ATXN3 | ATXN7 | CACNA1A | TBP | ||
| Binding proteins (binders) | PQBP1 | 2 | 3 | 3 | 9 | 8 | 0 | 2 | 1 | 4 |
| VCP | 10 | 6 | 4 | 5 | 7 | 29 | 5 | 2 | 12 | |
| Features | |||
|---|---|---|---|
| Specific | Shared | ||
| Binding proteins (binders) | PQBP1 | Regulation of transcription | Nucleoplasm (CC) Cytoplasm (CC) Protein binding (MF) |
| VCP | Ubiquitin-proteasome for ER stress | ||
| (a) | ||||
| Specific GO (2) | Protein Name | # PPIs (3) | Molecular Function | Biological Pathway |
| Present | Ataxin-1 (ATXN1) | 159 | RNA binding | Regulation of nucleic acid metabolism |
| Androgen receptor (AR) | 150 | Ligand-activated receptor | Signal transduction | |
| Huntingtin (HTT) | 59 | DNA binding | Regulation of nucleic acid metabolism | |
| Absent | Nuclear receptor coactivator 3 (NCOA3) | 47 | Transcriptional regulation | Intracellular signaling |
| (b) | ||||
| Specific GO (2) | Protein Name | # PPIs (3) | Molecular Function | Biological Pathway |
| n/a | Histone acetyltransferase p300 (EP300) | 209 | Transcriptional regulation | Regulation of nucleic acid metabolism |
| CREB-binding protein (CREBBP) | 198 | Transcriptional regulation | Regulation of nucleic acid metabolism | |
| Ubiquitin carboxyl-terminal hydrolase 7 (USP7) | 40 | Deubiquitinating enzyme | Protein metabolism | |
| Ran-binding protein 9 (RANBP9) | 28 | Enzyme binding | Protein-containing complex assembly | |
| Heterogeneous nuclear ribonucleoprotein D0 (HNRNPD) | 19 | mRNA binding | Regulation of nucleic acid metabolism | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimada, M.K. Splicing Modulators Are Involved in Human Polyglutamine Diversification via Protein Complexes Shuttling between Nucleus and Cytoplasm. Int. J. Mol. Sci. 2023, 24, 9622. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24119622
Shimada MK. Splicing Modulators Are Involved in Human Polyglutamine Diversification via Protein Complexes Shuttling between Nucleus and Cytoplasm. International Journal of Molecular Sciences. 2023; 24(11):9622. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24119622
Chicago/Turabian StyleShimada, Makoto K. 2023. "Splicing Modulators Are Involved in Human Polyglutamine Diversification via Protein Complexes Shuttling between Nucleus and Cytoplasm" International Journal of Molecular Sciences 24, no. 11: 9622. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24119622