
Stem/Progenitor Cells and Related Therapy in Bronchopulmonary Dysplasia
 ,
,
Abstract
:1. Introduction
2. Stem/Progenitor Cells in the Lung
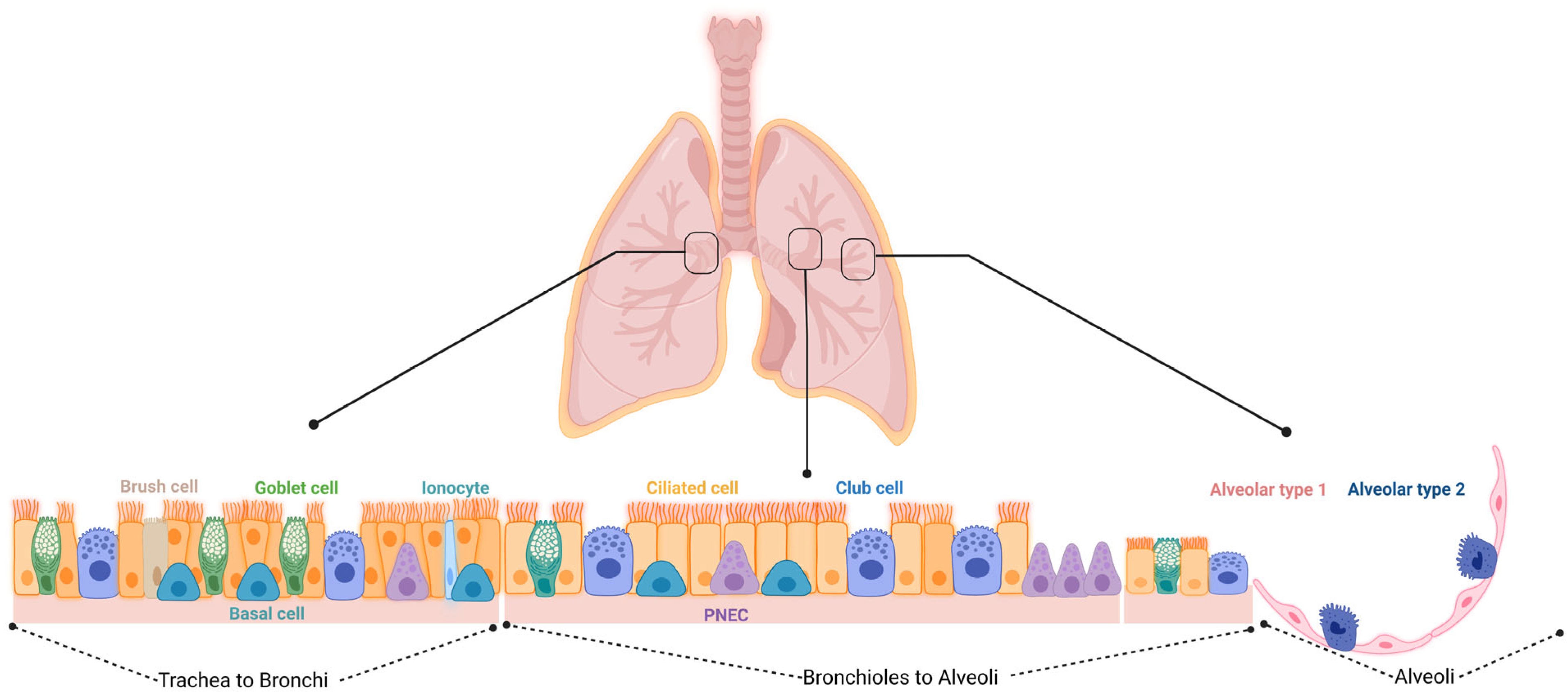
2.1. Epithelial Progenitors
2.1.1. Basal Cells (BCs)
2.1.2. Secretory Cells (SCs): Variant Club Cells (VCCs), Club Cells, and Goblet Cells
2.1.3. Pulmonary Neuroendocrine Cells (PNECs)
2.1.4. Alveolar Progenitor Cells

2.1.5. Non-Alveolar Progenitors
2.1.6. BASCs and DASCs
2.1.7. Lineage-Negative Epithelial Stem/Progenitors (LNEPs)

{kind=link}
{kind=link}
| Cell Type | Location | Markers | Differentiation Potential | Refs. |
|---|---|---|---|---|
| Basal cells (BCs) | Epithelium in the trachea and bronchioles | TP63, KRT5, KRT14, NGFR, PDPN, KRT8, N2ICD, and c-Myb/TP73 | Self-renewal; AT1, AT2 basal luminal precursor cell, neuroendocrine cells, ionocytes CCs, GCs, and ciliated cells | [23,87,88,89,90,91,92,93,94] |
| Variant club cells/ Clara cells | Bronchi and bronchioles (epithelium) | SCGB1A1 and CYP2F2− | Self-renewal: GCs and ciliated cells | [82,95,96] |
| Neuroendocrine cells (PNECs) | Epithelium, at branching points in the bronchi and bronchioles | GRP, ASCL1, CXCR4, NCAD and ROBO | Multiciliated cells and secretory cells | [49,55,97,98,99,100] |
| Broncho-alveolar stem cells (BASCs) | Epithelium and distal airway alveolar sacs | SCGB1A1 and SFTPC | Self-renewal: AT2 Ciliated cells: GCs and AT1 | [101,102,103,104] |
| AT2 | Distal airway alveolar sacs | SFTPC and LysM | Self-Renewal: AT1, SCA1+ mesenchymal progenitors, SFTPC+ AT1 progenitors, | [66,83,105,106,107,108] |
| Distal airway stem cells (DASCs), Alveolar progenitor | Distal airways, bronchioles, and alveolar regions | TP63 and KRT5 | Self-Renewal: AT2 | [84] |
| Self-Renewal: | [86] | |||
| AT2, SCGB1A1+, and CYP2F2+ CCs | [86,96] | |||
| AT1 | HOPX, | AT2 | [84] | |
| Club cells, ciliated cells | Terminal bronchioles and less in the respiratory bronchioles, trachea, and bronchioles | SCGB1A1, and CYP2F2 | BCs | [86] |
| Ciliated cells | [96] | |||
| TubIVa and FOXJ1 | GCs | [109] | ||
| Integrin α6β4+ alveolar progenitor | Terminal bronchioles, broncho-alveolar junctions, and less in the respiratory bronchioles | SCGB1A1−, SPC−, ITGα6, and ITGβ4 | Self-Renewal: AT1 Ciliated cells: AT2 and CCs | [96] |
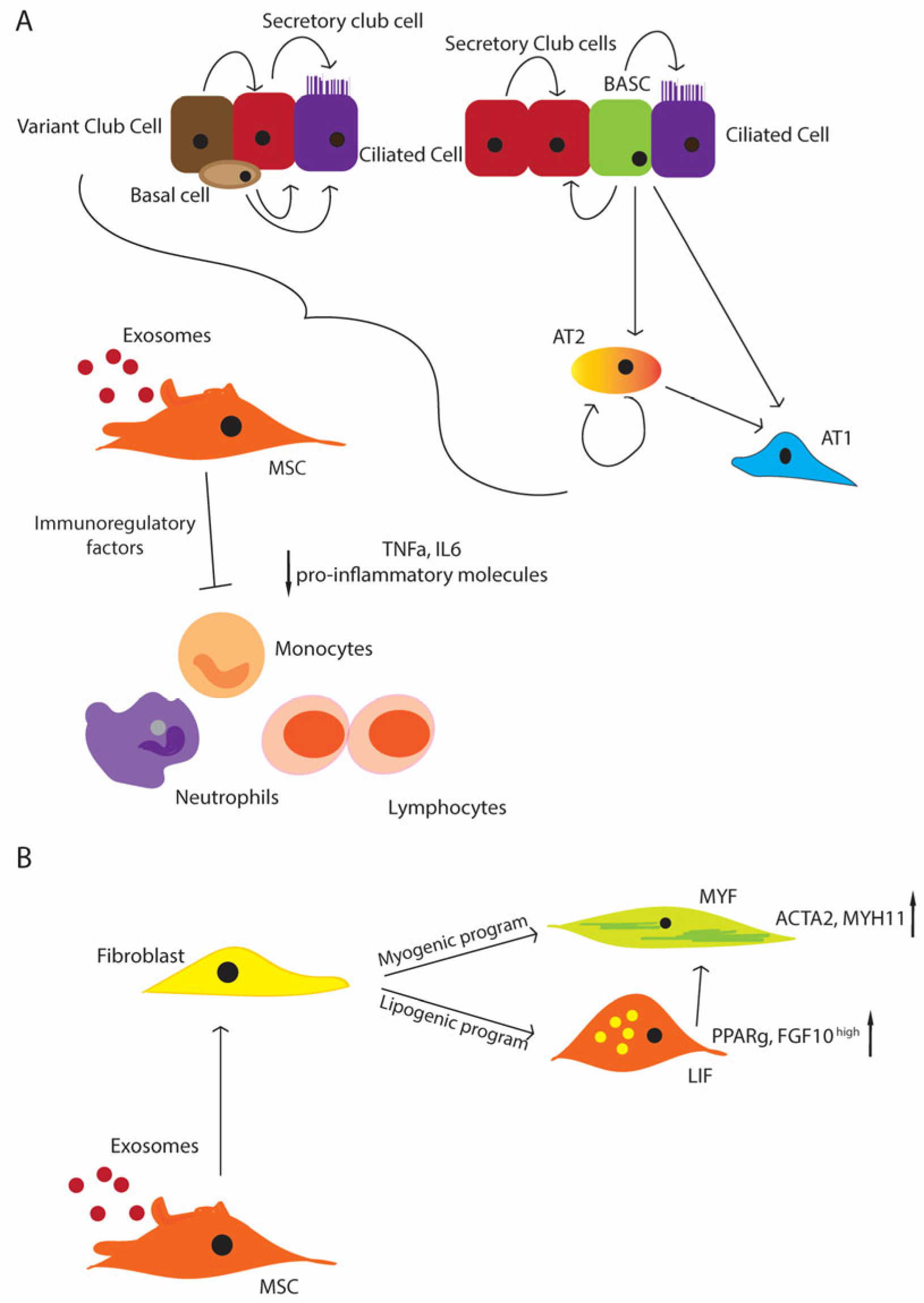
3. Role of EVs—Secretome of MSCs
4. Growth Factors and Inflammation in BPD
5. Fibroblast Growth Factor 10 (FGF10)
6. Vascular Endothelial Growth Factor (VEGF)
7. NF-κB
8. Use of Stem Cells for the Treatment of BPD
9. Animal Studies
10. Clinical Trials
11. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, L.; Oza, S.; Hogan, D.; Perin, J.; Rudan, I.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, Regional, and National Causes of Child Mortality in 2000–13, with Projections to Inform Post-2015 Priorities: An Updated Systematic Analysis. Lancet 2015, 385, 430–440. [Google Scholar] [CrossRef]
- Liu, L.; Johnson, H.L.; Cousens, S.; Perin, J.; Scott, S.; Lawn, J.E.; Rudan, I.; Campbell, H.; Cibulskis, R.; Li, M.; et al. Global, Regional, and National Causes of Child Mortality: An Updated Systematic Analysis for 2010 with Time Trends since 2000. Lancet 2012, 379, 2151–2161. [Google Scholar] [CrossRef] [PubMed]
- Botet, F.; Figueras-Aloy, J.; Miracle-Echegoyen, X.; Rodriguez-Miguelez, J.M.; Salvia-Roiges, M.D.; Carbonell-Estrany, X. Trends in Survival among Extremely-Low-Birth-Weight Infants (Less than 1000 g) without Significant Bronchopulmonary Dysplasia. BMC Pediatr. 2012, 12, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhandari, V.; Elias, J.A. Cytokines in Tolerance to Hyperoxia-Induced Injury in the Developing and Adult Lung. Free Radic. Biol. Med. 2006, 41, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, V.; Choo-Wing, R.; Lee, C.G.; Zhu, Z.; Nedrelow, J.H.; Chupp, G.L.; Zhang, X.; Matthay, M.A.; Ware, L.B.; Homer, R.J.; et al. Hyperoxia Causes Angiopoietin 2–Mediated Acute Lung Injury and Necrotic Cell Death. Nat. Med. 2006, 12, 1286–1293. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.K.; Hindori-Mohangoo, A.D.; Delnord, M.; Durox, M.; Szamotulska, K.; Macfarlane, A.; Alexander, S.; Barros, H.; Gissler, M.; Blondel, B.; et al. Quantifying the Burden of Stillbirths before 28 Weeks of Completed Gestational Age in High-Income Countries: A Population-Based Study of 19 European Countries. Lancet 2018, 392, 1639–1646. [Google Scholar] [CrossRef]
- Thébaud, B.; Goss, K.N.; Laughon, M.; Whitsett, J.A.; Abman, S.H.; Steinhorn, R.H.; Aschner, J.L.; Davis, P.G.; McGrath-Morrow, S.A.; Soll, R.F.; et al. Bronchopulmonary Dysplasia. Nat. Rev. Dis. Prim. 2019, 5, 78. [Google Scholar] [CrossRef]
- Mestan, K.K.; Leibel, S.L.; Sajti, E.; Pham, B.; Hietalati, S.; Laurent, L.; Parast, M. Leveraging the Placenta to Advance Neonatal Care. Front. Pediatr. 2023, 11, 1174174. [Google Scholar] [CrossRef]
- Oluwole, I.; Tan, J.B.C.; DeSouza, S.; Hutchinson, M.; Leigh, R.M.; Cha, M.; Rodriguez, A.; Hou, G.; Rao, S.S.; Narang, A.; et al. The Association between Bronchopulmonary Dysplasia Grade and Risks of Adverse Neurodevelopmental Outcomes among Preterm Infants Born at Less than 30 Weeks of Gestation. J. Matern. Fetal Neonatal Med. 2023, 36, 2167074. [Google Scholar] [CrossRef]
- Yeh, T.F.; Lin, Y.J.; Lin, H.C.; Huang, C.C.; Hsieh, W.S.; Lin, C.H.; Tsai, C.H. Outcomes at School Age after Postnatal Dexamethasone Therapy for Lung Disease of Prematurity. N. Engl. J. Med. 2004, 350, 1304–1313. [Google Scholar] [CrossRef]
- Jensen, E.A.; Wiener, L.E.; Rysavy, M.A.; Dysart, K.C.; Gantz, M.G.; Eichenwald, E.C.; Greenberg, R.G.; Harmon, H.M.; Laughon, M.M.; Watterberg, K.L.; et al. Assessment of Corticosteroid Therapy and Death or Disability According to Pretreatment Risk of Death or Bronchopulmonary Dysplasia in Extremely Preterm Infants. JAMA Netw. Open 2023, 6, e2312277. [Google Scholar] [CrossRef] [PubMed]
- Hillman, N.H.; Jobe, A.H. Preterm Lung and Brain Responses to Mechanical Ventilation and Corticosteroids. J. Perinatol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Homan, T.D.; Nayak, R.P. Short-and Long-Term Complications of Bronchopulmonary Dysplasia. Respir. Care 2021, 66, 1618–1629. [Google Scholar] [CrossRef]
- Shahzad, T.; Radajewski, S.; Chao, C.M.; Bellusci, S.; Ehrhardt, H. Pathogenesis of Bronchopulmonary Dysplasia: When Inflammation Meets Organ Development. Mol. Cell. Pediatr. 2016, 3, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behnke, J.; Kremer, S.; Shahzad, T.; Chao, C.M.; Bottcher-Friebertshauser, E.; Morty, R.E.; Bellusci, S.; Ehrhardt, H. MSC Based Therapies-New Perspectives for the Injured Lung. J. Clin. Med. 2020, 9, 682. [Google Scholar] [CrossRef] [Green Version]
- Goetz, M.J.; Kremer, S.; Behnke, J.; Staude, B.; Shahzad, T.; Holzfurtner, L.; Chao, C.M.; Morty, R.E.; Bellusci, S.; Ehrhardt, H. MSC Based Therapies to Prevent or Treat BPD-A Narrative Review on Advances and Ongoing Challenges. Int. J. Mol. Sci. 2021, 22, 1138. [Google Scholar] [CrossRef]
- Jiang, W.; Xu, J. Immune Modulation by Mesenchymal Stem Cells. Cell Prolif. 2020, 53, e12712. [Google Scholar] [CrossRef] [Green Version]
- Popova, A.P.; Bozyk, P.D.; Bentley, J.K.; Linn, M.J.; Goldsmith, A.M.; Schumacher, R.E.; Weiner, G.M.; Filbrun, A.G.; Hershenson, M.B. Isolation of Tracheal Aspirate Mesenchymal Stromal Cells Predicts Bronchopulmonary Dysplasia. Pediatrics 2010, 126, e1127–e1133. [Google Scholar] [CrossRef] [Green Version]
- Lignelli, E.; Palumbo, F.; Myti, D.; Morty, R.E. Recent Advances in Our Understanding of the Mechanisms of Lung Alveolarization and Bronchopulmonary Dysplasia. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2019, 317, L832–L887. [Google Scholar] [CrossRef]
- Alysandratos, K.-D.; Herriges, M.J.; Kotton, D.N. Epithelial Stem and Progenitor Cells in Lung Repair and Regeneration. Annu. Rev. Physiol. 2021, 83, 529–550. [Google Scholar] [CrossRef]
- Rock, J.R.; Randell, S.H.; Hogan, B.L.M. Airway Basal Stem Cells: A Perspective on Their Roles in Epithelial Homeostasis and Remodeling. Dis. Models Mech. 2010, 3, 545–556. [Google Scholar] [CrossRef] [Green Version]
- Hong, K.U.; Reynolds, S.D.; Watkins, S.; Fuchs, E.; Stripp, B.R. Basal Cells Are a Multipotent Progenitor Capable of Renewing the Bronchial Epithelium. Am. J. Pathol. 2004, 164, 577–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, K.U.; Reynolds, S.D.; Watkins, S.; Fuchs, E.; Stripp, B.R. In Vivo Differentiation Potential of Tracheal Basal Cells: Evidence for Multipotent and Unipotent Subpopulations. Am. J. Physiol. Lung Cell. Mol. Physiol. 2004, 286, L643–L649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rock, J.R.; Onaitis, M.W.; Rawlins, E.L.; Lu, Y.; Clark, C.P.; Xue, Y.; Randell, S.H.; Hogan, B.L. Basal Cells as Stem Cells of the Mouse Trachea and Human Airway Epithelium. Proc. Natl. Acad. Sci. USA 2009, 106, 12771–12775. [Google Scholar] [CrossRef] [PubMed]
- Montoro, D.T.; Haber, A.L.; Biton, M.; Vinarsky, V.; Lin, B.; Birket, S.E.; Yuan, F.; Chen, S.; Leung, H.M.; Villoria, J.; et al. A Revised Airway Epithelial Hierarchy Includes CFTR-Expressing Ionocytes. Nature 2018, 560, 319–324. [Google Scholar] [CrossRef]
- Nakajima, M.; Kawanami, O.; Jin, E.; Ghazizadeh, M.; Honda, M.; Asano, G.; Horiba, K.; Ferrans, V.J. Immunohistochemical and Ultrastructural Studies of Basal Cells, Clara Cells and Bronchiolar Cuboidal Cells in Normal Human Airways. Pathol. Int. 1998, 48, 944–953. [Google Scholar] [CrossRef]
- Boers, J.E.; Ambergen, A.W.; Thunnissen, F.B. Number and Proliferation of Clara Cells in Normal Human Airway Epithelium. Am. J. Respir. Crit. Care Med. 1999, 159, 1585–1591. [Google Scholar] [CrossRef] [Green Version]
- Tata, P.R.; Mou, H.; Pardo-Saganta, A.; Zhao, R.; Prabhu, M.; Law, B.M.; Vinarsky, V.; Cho, J.L.; Breton, S.; Sahay, A.; et al. Dedifferentiation of Committed Epithelial Cells into Stem Cells in Vivo. Nature 2013, 503, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Rock, J.R.; Gao, X.; Xue, Y.; Randell, S.H.; Kong, Y.Y.; Hogan, B.L. Notch-Dependent Differentiation of Adult Airway Basal Stem Cells. Cell Stem Cell 2011, 8, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Watson, J.K.; Rulands, S.; Wilkinson, A.C.; Wuidart, A.; Ousset, M.; Van Keymeulen, A.; Göttgens, B.; Blanpain, C.; Simons, B.D.; Rawlins, E.L. Clonal Dynamics Reveal Two Distinct Populations of Basal Cells in Slow-Turnover Airway Epithelium. Cell Rep. 2015, 12, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, V.H.; Nadarajan, P.; Graham, T.A.; Pipinikas, C.P.; Brown, J.M.; Falzon, M.; Nye, E.; Poulsom, R.; Lawrence, D.; Wright, N.A.; et al. Stochastic Homeostasis in Human Airway Epithelium Is Achieved by Neutral Competition of Basal Cell Progenitors. eLife 2013, 2, e00966. [Google Scholar] [CrossRef]
- Plasschaert, L.W.; Zilionis, R.; Choo-Wing, R.; Savova, V.; Knehr, J.; Roma, G.; Klein, A.M.; Jaffe, A.B. A Single-Cell Atlas of the Airway Epithelium Reveals the CFTR-Rich Pulmonary Ionocyte. Nature 2018, 560, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Carraro, G.; Mulay, A.; Yao, C.; Mizuno, T.; Konda, B.; Petrov, M.; Lafkas, D.; Arron, J.R.; Hogaboam, C.M.; Chen, P.; et al. Single-Cell Reconstruction of Human Basal Cell Diversity in Normal and Idiopathic Pulmonary Fibrosis Lungs. Am. J. Respir. Crit. Care Med. 2020, 202, 1540–1550. [Google Scholar] [CrossRef] [PubMed]
- Zaragosi, L.E.; Deprez, M.; Barbry, P. Using Single-Cell RNA Sequencing to Unravel Cell Lineage Relationships in the Respiratory Tract. Biochem. Soc. Trans. 2020, 48, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Ruiz Garcia, S.; Deprez, M.; Lebrigand, K.; Cavard, A.; Paquet, A.; Arguel, M.J.; Magnone, V.; Truchi, M.; Caballero, I.; Leroy, S.; et al. Novel Dynamics of Human Mucociliary Differentiation Revealed by Single-Cell RNA Sequencing of Nasal Epithelial Cultures. Development 2019, 146, 177428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaeger, B.; Schupp, J.C.; Plappert, L.; Terwolbeck, O.; Artysh, N.; Kayser, G.; Engelhard, P.; Adams, T.S.; Zweigerdt, R.; Kempf, H.; et al. Airway Basal Cells Show a Dedifferentiated KRT17highPhenotype and Promote Fibrosis in Idiopathic Pulmonary Fibrosis. Nat. Commun. 2022, 13, 5637. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Riccio, P.; Schotsaert, M.; Mori, M.; Lu, J.; Lee, D.K.; Garcia-Sastre, A.; Xu, J.; Cardoso, W. V Spatial-Temporal Lineage Restrictions of Embryonic P63(+) Progenitors Establish Distinct Stem Cell Pools in Adult Airways. Dev. Cell 2018, 44, 752–761.e4. [Google Scholar] [CrossRef] [Green Version]
- Shui, J.E.; Wang, W.; Liu, H.; Stepanova, A.; Liao, G.; Qian, J.; Ai, X.; Ten, V.; Lu, J.; Cardoso, W.V. Prematurity Alters the Progenitor Cell Program of the Upper Respiratory Tract of Neonates. Sci. Rep. 2021, 11, 10799. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Liu, Z. Clara Cell 10-KD Protein in Inflammatory Upper Airway Diseases. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 25–30. [Google Scholar] [CrossRef]
- Lee, R.M.; O’Brodovich, H. Airway Epithelial Damage in Premature Infants with Respiratory Failure. Am. Rev. Respir. Dis. 1988, 137, 450–457. [Google Scholar] [CrossRef]
- Zheng, D.; Limmon, G.V.; Yin, L.; Leung, N.H.; Yu, H.; Chow, V.T.; Chen, J. Regeneration of Alveolar Type I and II Cells from Scgb1a1-Expressing Cells Following Severe Pulmonary Damage Induced by Bleomycin and Influenza. PLoS ONE 2012, 7, e48451. [Google Scholar] [CrossRef] [Green Version]
- Whitsett, J.A. Airway Epithelial Differentiation and Mucociliary Clearance. Ann. Am. Thorac. Soc. 2018, 15, S143–S148. [Google Scholar] [CrossRef] [PubMed]
- Lumsden, A.B.; McLean, A.; Lamb, D. Goblet and Clara Cells of Human Distal Airways: Evidence for Smoking Induced Changes in Their Numbers. Thorax 1984, 39, 844–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira Braga, F.A.; Kar, G.; Berg, M.; Carpaij, O.A.; Polanski, K.; Simon, L.M.; Brouwer, S.; Gomes, T.; Hesse, L.; Jiang, J.; et al. A Cellular Census of Human Lungs Identifies Novel Cell States in Health and in Asthma. Nat. Med. 2019, 25, 1153–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Water, R.; Willems, L.N.; Van Muijen, G.N.; Franken, C.; Fransen, J.A.; Dijkman, J.H.; Kramps, J.A. Ultrastructural Localization of Bronchial Antileukoprotease in Central and Peripheral Human Airways by a Gold-Labeling Technique Using Monoclonal Antibodies. Am. Rev. Respir. Dis. 1986, 133, 882–890. [Google Scholar] [PubMed]
- Barth, P.J.; Wolf, M.; Ramaswamy, A. Distribution and Number of Clara Cells in the Normal and Disturbed Development of the Human Fetal Lung. Pediatr. Pathol. 1994, 14, 637–651. [Google Scholar] [CrossRef] [PubMed]
- Thomas, W.; Seidenspinner, S.; Kawczyńska-Leda, N.; Chmielnicka-Kopaczyk, M.; Marx, A.; Wirbelauer, J.; Szymankiewicz, M.; Speer, C.P. Clara Cell Secretory Protein in Tracheobronchial Aspirates and Umbilical Cord Serum of Extremely Premature Infants with Systemic Inflammation. Neonatology 2010, 97, 228–234. [Google Scholar] [CrossRef]
- Ramsay, P.L.; DeMayo, F.J.; Hegemier, S.E.; Wearden, M.E.; Smith, C.V.; Welty, S.E. Clara Cell Secretory Protein Oxidation and Expression in Premature Infants Who Develop Bronchopulmonary Dysplasia. Am. J. Respir. Crit. Care Med. 2001, 164, 155–161. [Google Scholar] [CrossRef]
- Schilders, K.A.; Eenjes, E.; van Riet, S.; Poot, A.A.; Stamatialis, D.; Truckenmuller, R.; Hiemstra, P.S.; Rottier, R.J. Regeneration of the Lung: Lung Stem Cells and the Development of Lung Mimicking Devices. Respir. Res. 2016, 17, 44. [Google Scholar] [CrossRef] [Green Version]
- Evans, M.J.; Plopper, C.G. The Role of Basal Cells in Adhesion of Columnar Epithelium to Airway Basement Membrane. Am. Rev. Respir. Dis. 1988, 138, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Hoyt, R.F., Jr.; Feldman, H.; Sorokin, S.P. Neuroepithelial Bodies (NEB) and Solitary Endocrine Cells in the Hamster Lung. Exp. Lung Res. 1982, 3, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.S.; Krasnow, M.A. Formation of a Neurosensory Organ by Epithelial Cell Slithering. Cell 2015, 163, 394–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weichselbaum, M.; Sparrow, M.P.; Hamilton, E.J.; Thompson, P.J.; Knight, D.A. A Confocal Microscopic Study of Solitary Pulmonary Neuroendocrine Cells in Human Airway Epithelium. Respir. Res. 2005, 6, 115. [Google Scholar] [CrossRef] [Green Version]
- Cutz, E.; Pan, J.; Yeger, H.; Domnik, N.J.; Fisher, J.T. Recent Advances and Contraversies on the Role of Pulmonary Neuroepithelial Bodies as Airway Sensors. Semin. Cell Dev. Biol. 2013, 24, 40–50. [Google Scholar] [CrossRef]
- Lee, R.J.; Kofonow, J.M.; Rosen, P.L.; Siebert, A.P.; Chen, B.; Doghramji, L.; Xiong, G.; Adappa, N.D.; Palmer, J.N.; Kennedy, D.W.; et al. Bitter and Sweet Taste Receptors Regulate Human Upper Respiratory Innate Immunity. J. Clin. Investig. 2014, 124, 1393–1405. [Google Scholar] [CrossRef]
- Branchfield, K.; Nantie, L.; Verheyden, J.M.; Sui, P.; Wienhold, M.D.; Sun, X. Pulmonary Neuroendocrine Cells Function as Airway Sensors to Control Lung Immune Response. Science 2016, 351, 707–710. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.E.; Lock, J.E.; Elde, R.P.; Thompson, T.R. Pulmonary Neuroendocrine Cells in Hyaline Membrane Disease and Bronchopulmonary Dysplasia. Pediatr. Res. 1982, 16, 446–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, D.E.; Kulik, T.J.; Lock, J.E.; Elde, R.P.; Thompson, T.R. Bombesin-, Calcitonin-, and Serotonin-Immunoreactive Pulmonary Neuroendocrine Cells in Acute and Chronic Neonatal Lung Disease. Pediatr. Pulmonol. 1985, 1, S13–S20. [Google Scholar]
- Cullen, A.; Van Marter, L.J.; Allred, E.N.; Moore, M.; Parad, R.B.; Sunday, M.E. Urine Bombesin-like Peptide Elevation Precedes Clinical Evidence of Bronchopulmonary Dysplasia. Am. J. Respir. Crit. Care Med. 2002, 165, 1093–1097. [Google Scholar] [CrossRef]
- Cutz, E.; Yeger, H.; Pan, J. Pulmonary Neuroendocrine Cell System in Pediatric Lung Disease-Recent Advances. Pediatr. Dev. Pathol. 2007, 10, 419–435. [Google Scholar] [CrossRef]
- Fujinaga, H.; Baker, C.D.; Ryan, S.L.; Markham, N.E.; Seedorf, G.J.; Balasubramaniam, V.; Abman, S.H. Hyperoxia Disrupts Vascular Endothelial Growth Factor-Nitric Oxide Signaling and Decreases Growth of Endothelial Colony-Forming Cells from Preterm Infants. Am. J. Physiol. Lung Cell. Mol. Physiol. 2009, 297, L1160–L1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, M.J.; Cabral, L.J.; Stephens, R.J.; Freeman, G. Transformation of Alveolar Type 2 Cells to Type 1 Cells Following Exposure to NO2. Exp. Mol. Pathol. 1975, 22, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Fehrenbach, H. Alveolar Epithelial Type II Cell: Defender of the Alveolus Revisited. Respir. Res. 2001, 2, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Beers, M.F.; Moodley, Y. When Is an Alveolar Type 2 Cell an Alveolar Type 2 Cell? A Conundrum for Lung Stem Cell Biology and Regenerative Medicine. Am. J. Respir. Cell Mol. Biol. 2017, 57, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Barkauskas, C.E.; Cronce, M.J.; Rackley, C.R.; Bowie, E.J.; Keene, D.R.; Stripp, B.R.; Randell, S.H.; Noble, P.W.; Hogan, B.L. Type 2 Alveolar Cells Are Stem Cells in Adult Lung. J. Clin. Investig. 2013, 123, 3025–3036. [Google Scholar] [CrossRef] [PubMed]
- Nabhan, A.N.; Brownfield, D.G.; Harbury, P.B.; Krasnow, M.A.; Desai, T.J. Single-Cell Wnt Signaling Niches Maintain Stemness of Alveolar Type 2 Cells. Science 2018, 359, 1118–1123. [Google Scholar] [CrossRef] [Green Version]
- Desai, T.J.; Brownfield, D.G.; Krasnow, M.A. Alveolar Progenitor and Stem Cells in Lung Development, Renewal and Cancer. Nature 2014, 507, 190–194. [Google Scholar] [CrossRef] [Green Version]
- Uhal, B.D. Cell Cycle Kinetics in the Alveolar Epithelium. Am. J. Physiol. 1997, 272, L1031–L1045. [Google Scholar] [CrossRef]
- Raman, T.; O’Connor, T.P.; Hackett, N.R.; Wang, W.; Harvey, B.G.; Attiyeh, M.A.; Dang, D.T.; Teater, M.; Crystal, R.G. Quality Control in Microarray Assessment of Gene Expression in Human Airway Epithelium. BMC Genom. 2009, 10, 493. [Google Scholar] [CrossRef] [Green Version]
- Maniscalco, W.M.; Watkins, R.H.; O’Reilly, M.A.; Shea, C.P. Increased Epithelial Cell Proliferation in Very Premature Baboons with Chronic Lung Disease. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2002, 283, L991–L1001. [Google Scholar] [CrossRef] [Green Version]
- Abman, S.H.; Collaco, J.M.; Shepherd, E.G.; Keszler, M.; Cuevas-Guaman, M.; Welty, S.E.; Truog, W.E.; McGrath-Morrow, S.A.; Moore, P.E.; Rhein, L.M.; et al. Interdisciplinary Care of Children with Severe Bronchopulmonary Dysplasia. J. Pediatr. 2017, 181, 12–28.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caskey, S.; Gough, A.; Rowan, S.; Gillespie, S.; Clarke, J.; Riley, M.; Megarry, J.; Nicholls, P.; Patterson, C.; Halliday, H.L.; et al. Structural and Functional Lung Impairment in Adult Survivors of Bronchopulmonary Dysplasia. Ann. Am. Thorac. Soc. 2016, 13, 1262–1270. [Google Scholar] [CrossRef] [Green Version]
- Thebaud, B.; Abman, S.H. Bronchopulmonary Dysplasia: Where Have All the Vessels Gone? Roles of Angiogenic Growth Factors in Chronic Lung Disease. Am. J. Respir. Crit. Care Med. 2007, 175, 978–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angusamy, S.; Mansour, T.; Abdulmageed, M.; Han, R.; Schutte, B.C.; LaPres, J.; Harkema, J.R.; Omar, S.A. Altered Thymocyte and T Cell Development in Neonatal Mice with Hyperoxia-Induced Lung Injury. J. Perinat. Med. 2018, 46, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Liu, S. Mouse Pneumonectomy Model of Compensatory Lung Growth. J. Vis. Exp. 2014, 94, e52294. [Google Scholar] [CrossRef] [Green Version]
- Voswinckel, R.; Motejl, V.; Fehrenbach, A.; Wegmann, M.; Mehling, T.; Fehrenbach, H.; Seeger, W. Characterisation of Post-Pneumonectomy Lung Growth in Adult Mice. Eur. Respir. J. 2004, 24, 524–532. [Google Scholar] [CrossRef] [Green Version]
- Lechner, A.J.; Driver, I.H.; Lee, J.; Conroy, C.M.; Nagle, A.; Locksley, R.M.; Rock, J.R. Recruited Monocytes and Type 2 Immunity Promote Lung Regeneration Following Pneumonectomy. Cell Stem Cell 2017, 21, 120–134.e7. [Google Scholar] [CrossRef] [Green Version]
- Ahmadvand, N.; Khosravi, F.; Lingampally, A.; Wasnick, R.; Vazquez-Armendariz, I.; Carraro, G.; Heiner, M.; Rivetti, S.; Lv, Y.; Wilhelm, J.; et al. Identification of a Novel Subset of Alveolar Type 2 Cells Enriched in PD-L1 and Expanded Following Pneumonectomy. Eur. Respir. J. 2021, 58, 2004168. [Google Scholar] [CrossRef]
- Kalymbetova, T.V.; Selvakumar, B.; Rodríguez-Castillo, J.A.; Gunjak, M.; Malainou, C.; Heindl, M.R.; Moiseenko, A.; Chao, C.M.; Vadász, I.; Mayer, K.; et al. Resident Alveolar Macrophages Are Master Regulators of Arrested Alveolarization in Experimental Bronchopulmonary Dysplasia. J. Pathol. 2018, 245, 153–159. [Google Scholar] [CrossRef]
- Shafa, M.; Ionescu, L.I.; Vadivel, A.; Collins, J.J.P.; Xu, L.; Zhong, S.; Kang, M.; de Caen, G.; Daneshmand, M.; Shi, J.; et al. Human Induced Pluripotent Stem Cell–Derived Lung Progenitor and Alveolar Epithelial Cells Attenuate Hyperoxia-Induced Lung Injury. Cytotherapy 2018, 20, 108–125. [Google Scholar] [CrossRef]
- Choi, J.; Park, J.-E.; Tsagkogeorga, G.; Yanagita, M.; Koo, B.-K.; Han, N.; Lee, J.-H. Inflammatory Signals Induce AT2 Cell-Derived Damage-Associated Transient Progenitors That Mediate Alveolar Regeneration. Cell Stem Cell 2020, 27, 366–382.e7. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Bhang, D.H.; Beede, A.; Huang, T.L.; Stripp, B.R.; Bloch, K.D.; Wagers, A.J.; Tseng, Y.H.; Ryeom, S.; Kim, C.F. Lung Stem Cell Differentiation in Mice Directed by Endothelial Cells via a BMP4-NFATc1-Thrombospondin-1 Axis. Cell 2014, 156, 440–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawlins, E.L.; Okubo, T.; Xue, Y.; Brass, D.M.; Auten, R.L.; Hasegawa, H.; Wang, F.; Hogan, B.L. The Role of Scgb1a1+ Clara Cells in the Long-Term Maintenance and Repair of Lung Airway, but Not Alveolar, Epithelium. Cell Stem Cell 2009, 4, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Chapman, H.A.; Li, X.; Alexander, J.P.; Brumwell, A.; Lorizio, W.; Tan, K.; Sonnenberg, A.; Wei, Y.; Vu, T.H. Integrin Alpha6beta4 Identifies an Adult Distal Lung Epithelial Population with Regenerative Potential in Mice. J. Clin. Investig. 2011, 121, 2855–2862. [Google Scholar] [CrossRef] [Green Version]
- Jain, R.; Barkauskas, C.E.; Takeda, N.; Bowie, E.J.; Aghajanian, H.; Wang, Q.; Padmanabhan, A.; Manderfield, L.J.; Gupta, M.; Li, D.; et al. Plasticity of Hopx(+) Type I Alveolar Cells to Regenerate Type II Cells in the Lung. Nat. Commun. 2015, 6, 6727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, W.; Zhang, T.; Wu, D.Z.; Guan, S.P.; Liew, A.A.; Yamamoto, Y.; Wang, X.; Lim, S.J.; Vincent, M.; Lessard, M.; et al. P63(+)Krt5(+) Distal Airway Stem Cells Are Essential for Lung Regeneration. Nature 2015, 517, 616–620. [Google Scholar] [CrossRef]
- Vaughan, A.E.; Brumwell, A.N.; Xi, Y.; Gotts, J.E.; Brownfield, D.G.; Treutlein, B.; Tan, K.; Tan, V.; Liu, F.C.; Looney, M.R.; et al. Lineage-Negative Progenitors Mobilize to Regenerate Lung Epithelium after Major Injury. Nature 2015, 517, 621–625. [Google Scholar] [CrossRef] [Green Version]
- Paul, M.K.; Bisht, B.; Darmawan, D.O.; Chiou, R.; Ha, V.L.; Wallace, W.D.; Chon, A.T.; Hegab, A.E.; Grogan, T.; Elashoff, D.A.; et al. Dynamic Changes in Intracellular ROS Levels Regulate Airway Basal Stem Cell Homeostasis through Nrf2-Dependent Notch Signaling. Cell Stem Cell 2014, 15, 199–214. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Fallon, T.R.; Saladi, S.V.; Pardo-Saganta, A.; Villoria, J.; Mou, H.; Vinarsky, V.; Gonzalez-Celeiro, M.; Nunna, N.; Hariri, L.P.; et al. Yap Tunes Airway Epithelial Size and Architecture by Regulating the Identity, Maintenance, and Self-Renewal of Stem Cells. Dev. Cell 2014, 30, 151–165. [Google Scholar] [CrossRef] [Green Version]
- Mori, M.; Mahoney, J.E.; Stupnikov, M.R.; Paez-Cortez, J.R.; Szymaniak, A.D.; Varelas, X.; Herrick, D.B.; Schwob, J.; Zhang, H.; Cardoso, W. V Notch3-Jagged Signaling Controls the Pool of Undifferentiated Airway Progenitors. Development 2015, 142, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Bali, A.S.; Randell, S.H.; Hogan, B.L. GRHL2 Coordinates Regeneration of a Polarized Mucociliary Epithelium from Basal Stem Cells. J. Cell Biol. 2015, 211, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, M.; Nishinakamura, R.; Saga, Y.; Kopan, R. Different Assemblies of Notch Receptors Coordinate the Distribution of the Major Bronchial Clara, Ciliated and Neuroendocrine Cells. Development 2012, 139, 4365–4373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, L.; Aster, J.C.; Sklar, J.; Sunday, M.E. Notch-1 Regulates Pulmonary Neuroendocrine Cell Differentiation in Cell Lines and in Transgenic Mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L500–L509. [Google Scholar] [CrossRef]
- Ito, T.; Udaka, N.; Yazawa, T.; Okudela, K.; Hayashi, H.; Sudo, T.; Guillemot, F.; Kageyama, R.; Kitamura, H. Basic Helix-Loop-Helix Transcription Factors Regulate the Neuroendocrine Differentiation of Fetal Mouse Pulmonary Epithelium. Development 2000, 127, 3913–3921. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.A.; Hu, Y.; Yamamoto, Y.; Hoe, N.B.; Wei, T.S.; Mu, D.; Sun, Y.; Joo, L.S.; Dagher, R.; Zielonka, E.M.; et al. Distal Airway Stem Cells Yield Alveoli In Vitro and during Lung Regeneration Following H1N1 Influenza Infection. Cell 2011, 147, 525–538. [Google Scholar] [CrossRef] [Green Version]
- Guha, A.; Vasconcelos, M.; Cai, Y.; Yoneda, M.; Hinds, A.; Qian, J.; Li, G.; Dickel, L.; Johnson, J.E.; Kimura, S.; et al. Neuroepithelial Body Microenvironment Is a Niche for a Distinct Subset of Clara-like Precursors in the Developing Airways. Proc. Natl. Acad. Sci. USA 2012, 109, 12592–12597. [Google Scholar] [CrossRef]
- Linnoila, R.I. Functional Facets of the Pulmonary Neuroendocrine System. Lab. Investig. 2006, 86, 425–444. [Google Scholar] [CrossRef] [Green Version]
- Minoo, P.; Su, G.; Drum, H.; Bringas, P.; Kimura, S. Defects in Tracheoesophageal and Lung Morphogenesis in Nkx2.1(−/−) Mouse Embryos. Dev. Biol. 1999, 209, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, M.; Sumiyama, K.; Morimoto, M. Directed Migration of Pulmonary Neuroendocrine Cells toward Airway Branches Organizes the Stereotypic Location of Neuroepithelial Bodies. Cell Rep. 2015, 13, 2679–2686. [Google Scholar] [CrossRef] [Green Version]
- Yao, E.; Lin, C.; Wu, Q.; Zhang, K.; Song, H.; Chuang, P.T. Notch Signaling Controls Transdifferentiation of Pulmonary Neuroendocrine Cells in Response to Lung Injury. Stem Cells 2018, 36, 377–391. [Google Scholar] [CrossRef] [Green Version]
- Borges, M.; Linnoila, R.I.; Van De Velde, H.J.K.; Chen, H.; Nelkin, B.D.; Mabry, M.; Baylin, S.B.; Ball, D.W. An Achaete-Scute Homologue Essential for Neuroendocrine Differentiation in the Lung. Nature 1997, 386, 852–855. [Google Scholar] [CrossRef] [PubMed]
- Bender Kim, C.F.; Jackson, E.L.; Woolfenden, A.E.; Lawrence, S.; Babar, I.; Vogel, S.; Crowley, D.; Bronson, R.T.; Jacks, T. Identification of Bronchioalveolar Stem Cells in Normal Lung and Lung Cancer. Cell 2005, 121, 823–835. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Goss, A.M.; Cohen, E.D.; Kadzik, R.; Lepore, J.J.; Muthukumaraswamy, K.; Yang, J.; DeMayo, F.J.; Whitsett, J.A.; Parmacek, M.S.; et al. A Gata6-Wnt Pathway Required for Epithelial Stem Cell Development and Airway Regeneration. Nat. Genet. 2008, 40, 862–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parekh, K.R.; Nawroth, J.; Pai, A.; Busch, S.M.; Senger, C.N.; Ryan, A.L. Stem Cells and Lung Regeneration. Am. J. Physiol. Cell Physiol. 2020, 319, C675–C693. [Google Scholar] [CrossRef] [PubMed]
- Lynch, T.J.; Liu, X.; Wei, J.; Engelhardt, J.F. Stem Cell Niches in the Lung. In Lung Stem Cells in the Epithelium and Vasculature; Firth, A., Yuan, J.X.J., Eds.; Springer International Publishing: Cham, Switzeland, 2015; pp. 35–58. ISBN 978-3-319-16232-4. [Google Scholar]
- Kapere Ochieng, J.; Schilders, K.; Kool, H.; Buscop-van Kempen, M.; Boerema-De Munck, A.; Grosveld, F.; Wijnen, R.; Tibboel, D.; Rottier, R.J. Differentiated Type II Pneumocytes Can Be Reprogrammed by Ectopic Sox2 Expression. PLoS ONE 2014, 9, e107248. [Google Scholar] [CrossRef]
- Liu, Y.; Kumar, V.S.; Zhang, W.; Rehman, J.; Malik, A.B. Activation of Type II Cells into Regenerative Stem Cell Antigen-1(+) Cells during Alveolar Repair. Am. J. Respir. Cell Mol. Biol. 2015, 53, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Summer, R.; Fitzsimmons, K.; Dwyer, D.; Murphy, J.; Fine, A. Isolation of an Adult Mouse Lung Mesenchymal Progenitor Cell Population. Am. J. Respir. Cell Mol. Biol. 2007, 37, 152–159. [Google Scholar] [CrossRef] [Green Version]
- Raiser, D.M.; Kim, C.F. Commentary: Sca-1 and Cells of the Lung: A Matter of Different Sorts. Stem Cells 2009, 27, 606–611. [Google Scholar] [CrossRef] [Green Version]
- Willis, G.R.; Fernandez-Gonzalez, A.; Anastas, J.; Vitali, S.H.; Liu, X.; Ericsson, M.; Kwong, A.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal Stromal Cell Exosomes Ameliorate Experimental Bronchopulmonary Dysplasia and Restore Lung Function through Macrophage Immunomodulation. Am. J. Respir. Crit. Care Med. 2018, 197, 104–116. [Google Scholar] [CrossRef]
- Willis, G.R.; Reis, M.; Gheinani, A.H.; Fernandez-Gonzalez, A.; Taglauer, E.S.; Yeung, V.; Liu, X.; Ericsson, M.; Haas, E.; Mitsialis, S.A.; et al. Extracellular Vesicles Protect the Neonatal Lung from Hyperoxic Injury through the Epigenetic and Transcriptomic Reprogramming of Myeloid Cells. Am. J. Respir. Crit. Care Med. 2021, 204, 1418–1432. [Google Scholar] [CrossRef]
- Rani, S.; Ryan, A.E.; Griffin, M.D.; Ritter, T. Mesenchymal Stem Cell-Derived Extracellular Vesicles: Toward Cell-Free Therapeutic Applications. Mol. Ther. 2015, 23, 812–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worthington, E.N.; Hagood, J.S. Therapeutic Use of Extracellular Vesicles for Acute and Chronic Lung Disease. Int. J. Mol. Sci. 2020, 21, 2318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowen, A.; Shahjin, F.; Chand, S.; Odegaard, K.E.; Yelamanchili, S. V Mesenchymal Stem Cell-Derived Extracellular Vesicles: Challenges in Clinical Applications. Front. Cell Dev. Biol. 2020, 8, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Dong, Z.; Cao, Y.; Wang, H.; Liu, S.; Liao, L.; Jin, Y.; Yuan, L.; Li, B. MSC-Derived Exosome Promotes M2 Polarization and Enhances Cutaneous Wound Healing. Stem Cells Int. 2019, 2019, 7132708. [Google Scholar] [CrossRef] [Green Version]
- Aguiar Koga, B.A.; Fernandes, L.A.; Fratini, P.; Sogayar, M.C.; Carreira, A.C.O. Role of MSC-Derived Small Extracellular Vesicles in Tissue Repair and Regeneration. Front. Cell Dev. Biol. 2022, 10, 1047094. [Google Scholar] [CrossRef] [PubMed]
- Karami Fath, M.; Moayedi Banan, Z.; Barati, R.; Mohammadrezakhani, O.; Ghaderi, A.; Hatami, A.; Ghiabi, S.; Zeidi, N.; Asgari, K.; Payandeh, Z.; et al. Recent Advancements to Engineer Mesenchymal Stem Cells and Their Extracellular Vesicles for Targeting and Destroying Tumors. Prog. Biophys. Mol. Biol. 2023, 178, 1–16. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, L.; Zhang, M.; Chen, Z. Mesenchymal Stem Cell-Derived Small Extracellular Vesicles: A Novel Approach for Kidney Disease Treatment. Int. J. Nanomed. 2022, 17, 3603–3618. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Z.; Li, J.; Zhong, H.; Yuan, R.; Deng, Z.; Wu, X. Mechanism of Adipose-Derived Mesenchymal Stem Cell-Derived Extracellular Vesicles Carrying MiR-21-5p in Hyperoxia-Induced Lung Injury. Stem Cell Rev. Rep. 2022, 18, 1007–1024. [Google Scholar] [CrossRef]
- Xi, Y.; Ju, R.; Wang, Y. Mesenchymal Stem Cell-Derived Extracellular Vesicles for the Treatment of Bronchopulmonary Dysplasia. Front. Pediatr. 2022, 10, 852034. [Google Scholar] [CrossRef]
- Ai, D.; Shen, J.; Sun, J.; Zhu, Z.; Gao, R.; Du, Y.; Yuan, L.; Chen, C.; Zhou, J. Mesenchymal Stem Cell-Derived Extracellular Vesicles Suppress Hyperoxia-Induced Transdifferentiation of Rat Alveolar Type 2 Epithelial Cells. Stem Cells Dev. 2022, 31, 53–66. [Google Scholar] [CrossRef]
- El Agha, E.; Bellusci, S. Walking along the Fibroblast Growth Factor 10 Route: A Key Pathway to Understand the Control and Regulation of Epithelial and Mesenchymal Cell-Lineage Formation during Lung Development and Repair after Injury. Scientifica 2014, 2014, 538379. [Google Scholar] [CrossRef]
- Al Alam, D.; El Agha, E.; Sakurai, R.; Kheirollahi, V.; Moiseenko, A.; Danopoulos, S.; Shrestha, A.; Schmoldt, C.; Quantius, J.; Herold, S.; et al. Evidence for the Involvement of Fibroblast Growth Factor 10 in Lipofibroblast Formation during Embryonic Lung Development. Development 2015, 142, 4139–4150. [Google Scholar] [CrossRef] [Green Version]
- McGowan, S.E. The Lipofibroblast: More than a Lipid-Storage Depot. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 316, L869–L871. [Google Scholar] [CrossRef] [PubMed]
- Rehan, V.K.; Torday, J.S. The Lung Alveolar Lipofibroblast: An Evolutionary Strategy against Neonatal Hyperoxic Lung Injury. Antioxid. Redox Signal. 2014, 21, 1893–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupte, V.V.; Ramasamy, S.K.; Reddy, R.; Lee, J.; Weinreb, P.H.; Violette, S.M.; Guenther, A.; Warburton, D.; Driscoll, B.; Minoo, P.; et al. Overexpression of Fibroblast Growth Factor-10 during Both Inflammatory and Fibrotic Phases Attenuates Bleomycin-Induced Pulmonary Fibrosis in Mice. Am. J. Respir. Crit. Care Med. 2009, 180, 424–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, T.; Volckaert, T.; Chanda, D.; Thannickal, V.J.; De Langhe, S.P. Fgf10 Signaling in Lung Development, Homeostasis, Disease, and Repair After Injury. Front. Genet. 2018, 9, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, C.M.; Moiseenko, A.; Kosanovic, D.; Rivetti, S.; El Agha, E.; Wilhelm, J.; Kampschulte, M.; Yahya, F.; Ehrhardt, H.; Zimmer, K.P.; et al. Impact of Fgf10 Deficiency on Pulmonary Vasculature Formation in a Mouse Model of Bronchopulmonary Dysplasia. Hum. Mol. Genet. 2019, 28, 1429–1444. [Google Scholar] [CrossRef]
- Tong, L.; Zhou, J.; Rong, L.; Seeley, E.J.; Pan, J.; Zhu, X.; Liu, J.; Wang, Q.; Tang, X.; Qu, J.; et al. Fibroblast Growth Factor-10 (FGF-10) Mobilizes Lung-Resident Mesenchymal Stem Cells and Protects against Acute Lung Injury. Sci. Rep. 2016, 6, 21642. [Google Scholar] [CrossRef] [Green Version]
- Taghizadeh, S.; Chao, C.-M.; Guenther, S.; Glaser, L.; Gersmann, L.; Michel, G.; Kraut, S.; Goth, K.; Koepke, J.; Heiner, M.; et al. FGF10 Triggers De Novo Alveologenesis in a Bronchopulmonary Dysplasia Model: Impact on Resident Mesenchymal Niche Cells. Stem Cells 2022, 40, 605–617. [Google Scholar] [CrossRef]
- Yu, P.; Wilhelm, K.; Dubrac, A.; Tung, J.K.; Alves, T.C.; Fang, J.S.; Xie, Y.; Zhu, J.; Chen, Z.; De Smet, F.; et al. FGF-Dependent Metabolic Control of Vascular Development. Nature 2017, 545, 224–228. [Google Scholar] [CrossRef] [Green Version]
- Khaki, M.; Salmanian, A.H.; Abtahi, H.; Ganji, A.; Mosayebi, G. Mesenchymal Stem Cells Differentiate to Endothelial Cells Using Recombinant Vascular Endothelial Growth Factor—A. Rep. Biochem. Mol. Biol. 2018, 6, 144–150. [Google Scholar]
- Ahn, S.Y.; Park, W.S.; Kim, Y.E.; Sung, D.K.; Sung, S.I.; Ahn, J.Y.; Chang, Y.S. Vascular Endothelial Growth Factor Mediates the Therapeutic Efficacy of Mesenchymal Stem Cell-Derived Extracellular Vesicles against Neonatal Hyperoxic Lung Injury. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalikkot Thekkeveedu, R.; Guaman, M.C.; Shivanna, B. Bronchopulmonary Dysplasia: A Review of Pathogenesis and Pathophysiology. Respir. Med. 2017, 132, 170–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munshi, U.K.; Niu, J.O.; Siddiq, M.M.; Parton, L.A. Elevation of Interleukin-8 and Interleukin-6 Precedes the Influx of Neutrophils in Tracheal Aspirates from Preterm Infants Who Develop Bronchopulmonary Dysplasia. Pediatr. Pulmonol. 1997, 24, 331–336. [Google Scholar] [CrossRef]
- Wright, C.J.; Kirpalani, H. Targeting Inflammation to Prevent Bronchopulmonary Dysplasia: Can New Insights Be Translated into Therapies? Pediatrics 2011, 128, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Moodie, F.M.; Marwick, J.A.; Anderson, C.S.; Szulakowski, P.; Biswas, S.K.; Bauter, M.R.; Kilty, I.; Rahman, I. Oxidative Stress and Cigarette Smoke Alter Chromatin Remodeling but Differentially Regulate NF-KappaB Activation and Proinflammatory Cytokine Release in Alveolar Epithelial Cells. FASEB J. 2004, 18, 1897–1899. [Google Scholar] [CrossRef]
- Everhart, M.B.; Han, W.; Sherrill, T.P.; Arutiunov, M.; Polosukhin, V.V.; Burke, J.R.; Sadikot, R.T.; Christman, J.W.; Yull, F.E.; Blackwell, T.S. Duration and Intensity of NF-KappaB Activity Determine the Severity of Endotoxin-Induced Acute Lung Injury. J. Immunol. 2006, 176, 4995–5005. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T. The Nuclear Factor NF-KappaB Pathway in Inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T.; Fong, C. The Resolution of Inflammation: Anti-Inflammatory Roles for NF-KappaB. Int. J. Biochem. Cell Biol. 2010, 42, 519–523. [Google Scholar] [CrossRef]
- Barkett, M.; Gilmore, T.D. Control of Apoptosis by Rel/NF-KappaB Transcription Factors. Oncogene 1999, 18, 6910–6924. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, D.; Textor, B.; Pein, O.T.; Licht, A.H.; Andrecht, S.; Sator-Schmitt, M.; Fusenig, N.E.; Angel, P.; Schorpp-Kistner, M. Critical Role for NF-KappaB-Induced JunB in VEGF Regulation and Tumor Angiogenesis. EMBO J. 2007, 26, 710–719. [Google Scholar] [CrossRef] [Green Version]
- Hinz, M.; Krappmann, D.; Eichten, A.; Heder, A.; Scheidereit, C.; Strauss, M. NF-KappaB Function in Growth Control: Regulation of Cyclin D1 Expression and G0/G1-to-S-Phase Transition. Mol. Cell. Biol. 1999, 19, 2690–2698. [Google Scholar] [CrossRef] [Green Version]
- Blackwell, T.S.; Hipps, A.N.; Yamamoto, Y.; Han, W.; Barham, W.J.; Ostrowski, M.C.; Yull, F.E.; Prince, L.S. NF-ΚB Signaling in Fetal Lung Macrophages Disrupts Airway Morphogenesis. J. Immunol. 2011, 187, 2740–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvira, C.M.; Abate, A.; Yang, G.; Dennery, P.A.; Rabinovitch, M. Nuclear Factor-KappaB Activation in Neonatal Mouse Lung Protects against Lipopolysaccharide-Induced Inflammation. Am. J. Respir. Crit. Care Med. 2007, 175, 805–815. [Google Scholar] [CrossRef] [Green Version]
- Iosef, C.; Alastalo, T.P.; Hou, Y.; Chen, C.; Adams, E.S.; Lyu, S.C.; Cornfield, D.N.; Alvira, C.M. Inhibiting NF-KappaB in the Developing Lung Disrupts Angiogenesis and Alveolarization. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 302, L1023–L1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourbia, A.; Cruz, M.A.; Rozycki, H.J. NF-KappaB in Tracheal Lavage Fluid from Intubated Premature Infants: Association with Inflammation, Oxygen, and Outcome. Arch. Dis. Child Fetal Neonatal Ed. 2006, 91, F36–F39. [Google Scholar] [CrossRef] [Green Version]
- Alvira, C.M. Nuclear Factor-Kappa-B Signaling in Lung Development and Disease: One Pathway, Numerous Functions. Birth Defects Res. Part A Clin. Mol. Teratol. 2014, 100, 202–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Hirschfeld, A.F.; Mayer, M.L.; Fortuno, E.S., 3rd; Corbett, N.; Kaplan, M.; Wang, S.; Schneiderman, J.; Fjell, C.D.; Yan, J.; et al. Functional Genetic Variation in NFKBIA and Susceptibility to Childhood Asthma, Bronchiolitis, and Bronchopulmonary Dysplasia. J. Immunol. 2013, 190, 3949–3958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reicherzer, T.; Häffner, S.; Shahzad, T.; Gronbach, J.; Mysliwietz, J.; Hübener, C.; Hasbargen, U.; Gertheiss, J.; Schulze, A.; Bellusci, S.; et al. Activation of the NF-KB Pathway Alters the Phenotype of MSCs in the Tracheal Aspirates of Preterm Infants with Severe BPD. Am. J. Physiol. Lung Cell. Mol. Physiol. 2018, 315, L87–L101. [Google Scholar] [CrossRef] [PubMed]
- Kaltschmidt, C.; Greiner, J.F.W.; Kaltschmidt, B. The Transcription Factor Nf-Κb in Stem Cells and Development. Cells 2021, 10, 2042. [Google Scholar] [CrossRef]
- Joel, M.D.M.; Yuan, J.; Wang, J.; Yan, Y.; Qian, H.; Zhang, X.; Xu, W.; Mao, F. MSC: Immunoregulatory Effects, Roles on Neutrophils and Evolving Clinical Potentials. Am. J. Transl. Res. 2019, 11, 3890. [Google Scholar] [PubMed]
- Prince, L.S. FGF10 and Human Lung Disease Across the Life Spectrum. Front. Genet. 2018, 9, 517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popova, A.P.; Bentley, J.K.; Cui, T.X.; Richardson, M.N.; Linn, M.J.; Lei, J.; Chen, Q.; Goldsmith, A.M.; Pryhuber, G.S.; Hershenson, M.B. Reduced Platelet-Derived Growth Factor Receptor Expression Is a Primary Feature of Human Bronchopulmonary Dysplasia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 307, L231–L239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierro, M.; Ionescu, L.; Montemurro, T.; Vadivel, A.; Weissmann, G.; Oudit, G.; Emery, D.; Bodiga, S.; Eaton, F.; Peault, B.; et al. Short-Term, Long-Term and Paracrine Effect of Human Umbilical Cord-Derived Stem Cells in Lung Injury Prevention and Repair in Experimental Bronchopulmonary Dysplasia. Thorax 2013, 68, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Chu, X.; Lingampally, A.; Moiseenko, A.; Kheirollahi, V.; Vazquez-Armendariz, A.I.; Koepke, J.; Khadim, A.; Kiliaris, G.; Shahriari Felordi, M.; Zabihi, M.; et al. GLI1+ Cells Are a Source of Repair-Supportive Mesenchymal Cells (RSMCs) during Airway Epithelial Regeneration. Cell. Mol. Life Sci. 2022, 79, 581. [Google Scholar] [CrossRef]
- Agostini, C. Stem Cell Therapy for Chronic Lung Diseases: Hope and Reality. Respir. Med. 2010, 104 (Suppl. 1), S86–S91. [Google Scholar] [CrossRef]
- Blaisdell, C.J.; Gail, D.B.; Nabel, E.G. National Heart, Lung, and Blood Institute Perspective: Lung Progenitor and Stem Cells—Gaps in Knowledge and Future Opportunities. Stem Cells 2009, 27, 2263–2270. [Google Scholar] [CrossRef] [Green Version]
- Sueblinvong, V.; Weiss, D.J. Stem Cells and Cell Therapy Approaches in Lung Biology and Diseases. Transl. Res. 2010, 156, 188–205. [Google Scholar] [CrossRef]
- Murphy, S.; Lim, R.; Dickinson, H.; Acharya, R.; Rosli, S.; Jenkin, G.; Wallace, E. Human Amnion Epithelial Cells Prevent Bleomycin-Induced Lung Injury and Preserve Lung Function. Cell Transplant. 2011, 20, 909–923. [Google Scholar] [CrossRef]
- Aguilar, S.; Scotton, C.J.; McNulty, K.; Nye, E.; Stamp, G.; Laurent, G.; Bonnet, D.; Janes, S.M. Bone Marrow Stem Cells Expressing Keratinocyte Growth Factor via an Inducible Lentivirus Protects against Bleomycin-Induced Pulmonary Fibrosis. PLoS ONE 2009, 4, e8013. [Google Scholar] [CrossRef]
- Gupta, N.; Su, X.; Popov, B.; Lee, J.W.; Serikov, V.; Matthay, M.A. Intrapulmonary Delivery of Bone Marrow-Derived Mesenchymal Stem Cells Improves Survival and Attenuates Endotoxin-Induced Acute Lung Injury in Mice. J. Immunol. 2007, 179, 1855–1863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, S.S.; Torres-Gonzalez, E.; Neujahr, D.C.; Kwon, M.; Brigham, K.L.; Jones, D.P.; Mora, A.L.; Rojas, M. Effect of Bone Marrow-Derived Mesenchymal Stem Cells on Endotoxin-Induced Oxidation of Plasma Cysteine and Glutathione in Mice. Stem Cells Int. 2010, 2010, 868076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tropea, K.A.; Leder, E.; Aslam, M.; Lau, A.N.; Raiser, D.M.; Lee, J.H.; Balasubramaniam, V.; Fredenburgh, L.E.; Alex Mitsialis, S.; Kourembanas, S.; et al. Bronchioalveolar Stem Cells Increase after Mesenchymal Stromal Cell Treatment in a Mouse Model of Bronchopulmonary Dysplasia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 302, L829–L837. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Baveja, R.; Liang, O.D.; Fernandez-Gonzalez, A.; Lee, C.; Mitsialis, S.A.; Kourembanas, S. Bone Marrow Stromal Cells Attenuate Lung Injury in a Murine Model of Neonatal Chronic Lung Disease. Am. J. Respir. Crit. Care Med. 2009, 180, 1122–1130. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Wang, H.; Shi, Y.; Peng, W.; Zhang, S.; Zhang, W.; Xu, J.; Mei, Y.; Feng, Z. Role of Bone Marrow-Derived Mesenchymal Stem Cells in the Prevention of Hyperoxia-Induced Lung Injury in Newborn Mice. Cell Biol. Int. 2012, 36, 589–594. [Google Scholar] [CrossRef]
- Hansmann, G.; Fernandez-Gonzalez, A.; Aslam, M.; Vitali, S.H.; Martin, T.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal Stem Cell-Mediated Reversal of Bronchopulmonary Dysplasia and Associated Pulmonary Hypertension. Pulm. Circ. 2012, 2, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Vosdoganes, P.; Hodges, R.J.; Lim, R.; Westover, A.J.; Acharya, R.Y.; Wallace, E.M.; Moss, T.J. Human Amnion Epithelial Cells as a Treatment for Inflammation-Induced Fetal Lung Injury in Sheep. Am. J. Obstet. Gynecol. 2011, 205, 156-e26. [Google Scholar] [CrossRef]
- Popova, A.P.; Bentley, J.K.; Anyanwu, A.C.; Richardson, M.N.; Linn, M.J.; Lei, J.; Wong, E.J.; Goldsmith, A.M.; Pryhuber, G.S.; Hershenson, M.B. Glycogen Synthase Kinase-3beta/Beta-Catenin Signaling Regulates Neonatal Lung Mesenchymal Stromal Cell Myofibroblastic Differentiation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 303, L439–L448. [Google Scholar] [CrossRef]
- Chang, Y.S.; Ahn, S.Y.; Yoo, H.S.; Sung, S.I.; Choi, S.J.; Oh, W.I.; Park, W.S. Mesenchymal Stem Cells for Bronchopulmonary Dysplasia: Phase 1 Dose-Escalation Clinical Trial. J. Pediatr. 2014, 164, 966–972.e6. [Google Scholar] [CrossRef]
- Lim, R.; Malhotra, A.; Tan, J.; Chan, S.T.; Lau, S.; Zhu, D.; Mockler, J.C.; Wallace, E.M. First-In-Human Administration of Allogeneic Amnion Cells in Premature Infants With Bronchopulmonary Dysplasia: A Safety Study. Stem Cells Transl. Med. 2018, 7, 628–635. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, A.; Lim, R.; Mockler, J.C.; Wallace, E.M. Two-Year Outcomes of Infants Enrolled in the First-in-Human Study of Amnion Cells for Bronchopulmonary Dysplasia. Stem Cells Transl. Med. 2020, 9, 289–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, E.K.; Malhotra, A.; Lim, R.; Jacobs, S.E.; Hooper, S.B.; Davis, P.G.; Wallace, E.M. Human Amnion Cells for the Prevention of Bronchopulmonary Dysplasia: A Protocol for a Phase I Dose Escalation Study. BMJ Open 2019, 9, e026265. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marega, M.; El-Merhie, N.; Gökyildirim, M.Y.; Orth, V.; Bellusci, S.; Chao, C.-M. Stem/Progenitor Cells and Related Therapy in Bronchopulmonary Dysplasia. Int. J. Mol. Sci. 2023, 24, 11229. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241311229
Marega M, El-Merhie N, Gökyildirim MY, Orth V, Bellusci S, Chao C-M. Stem/Progenitor Cells and Related Therapy in Bronchopulmonary Dysplasia. International Journal of Molecular Sciences. 2023; 24(13):11229. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241311229
Chicago/Turabian StyleMarega, Manuela, Natalia El-Merhie, Mira Y. Gökyildirim, Valerie Orth, Saverio Bellusci, and Cho-Ming Chao. 2023. "Stem/Progenitor Cells and Related Therapy in Bronchopulmonary Dysplasia" International Journal of Molecular Sciences 24, no. 13: 11229. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241311229