
Simulating Space Conditions Evokes Different DNA Damage Responses in Immature and Mature Cells of the Human Hematopoietic System
 , ,
, , 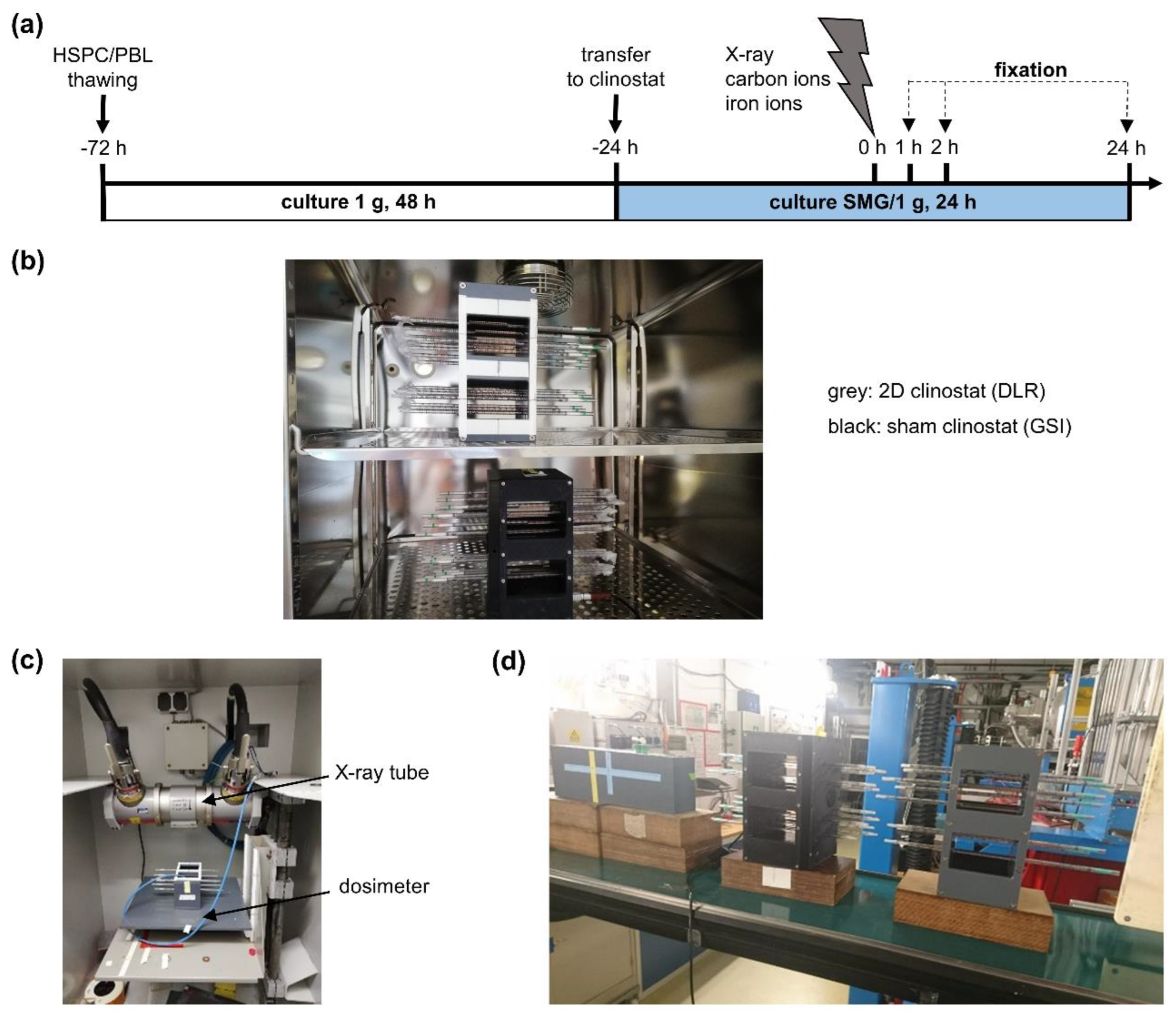{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Simulated Microgravity Accelerates Repair of Low LET Radiation-Induced DNA Damage in PBL
2.2. Simulated Microgravity Does Not Modulate X-ray-Induced Effects on Genomic Integrity, Survival or Differentiation Potential
2.3. Simulated Microgravity Does Not Affect Cell Cycle Progression
2.4. Simulated Microgravity Reduces End Processing and Homology-Directed DSB Repair
2.5. Simulated Microgravity Induces Replication Stress
2.6. NF-κB Signaling Contributes to the Accelerated Repair of DSBs in PBL Cultured in SMG
3. Discussion
3.1. SMG Accelerates Repair of Simple DSBs in PBL
3.2. Simulated Microgravity Cell Type Specifically Modulates DSB Repair
3.3. Replication Stress as DSB Repair Pre-Activating Mechanism under SMG Conditions
4. Methods and Materials
4.1. Isolation of Primary Hematopoietic Cells
4.2. Simulated Microgravity Conditions (Clinostat)
4.3. Irradiation with Photons and Heavy Ions and Drug Treatment
4.4. Quantitative Immunofluorescence Microscopy
4.5. Micronucleus Test
4.6. Colony-Forming-Unit-Assay
4.7. Cell Cycle and Sub G1 Analysis
4.8. EdU Staining
4.9. Immunoblotting
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Durante, M.; Cucinotta, F.A. Physical Basis of Radiation Protection in Space Travel. Rev. Mod. Phys. 2011, 83, 1245–1281. [Google Scholar] [CrossRef]
- Yatagai, F.; Honma, M.; Dohmae, N.; Ishioka, N. Biological Effects of Space Environmental Factors: A Possible Interaction between Space Radiation and Microgravity. Life Sci. Sp. Res. 2019, 20, 113–123. [Google Scholar] [CrossRef]
- Guéguinou, N.; Huin-Schohn, C.; Bascove, M.; Bueb, J.-L.; Tschirhart, E.; Legrand-Frossi, C.; Frippiat, J.-P. Could Spaceflight-Associated Immune System Weakening Preclude the Expansion of Human Presence beyond Earth’s Orbit? J. Leukoc. Biol. 2009, 86, 1027–1038. [Google Scholar] [CrossRef]
- Moreno-Villanueva, M.; Wong, M.; Lu, T.; Zhang, Y.; Wu, H. Interplay of Space Radiation and Microgravity in DNA Damage and DNA Damage Response. NPJ Microgravity 2017, 3, 14. [Google Scholar] [CrossRef]
- George, K.; Chappell, L.J.; Cucinotta, F.A. Persistence of Space Radiation Induced Cytogenetic Damage in the Blood Lymphocytes of Astronauts. Mutat. Res.—Genet. Toxicol. Environ. Mutagen. 2010, 701, 75–79. [Google Scholar] [CrossRef]
- Ali, Y.F.; Cucinotta, F.A.; Ning-Ang, L.; Zhou, G. Cancer Risk of Low Dose Ionizing Radiation. Front. Phys. 2020, 8, 234. [Google Scholar] [CrossRef]
- Durante, M.; Cucinotta, F.A. Heavy Ion Carcinogenesis and Human Space Exploration. Nat. Rev. Cancer 2008, 8, 465–472. [Google Scholar] [CrossRef]
- Cucinotta, F.A. Space Radiation Risks for Astronauts on Multiple International Space Station Missions. PLoS ONE 2014, 9, e96099. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; To, K.; Cacao, E. Predictions of Space Radiation Fatality Risk for Exploration Missions. Life Sci. Sp. Res. 2017, 13, 1–11. [Google Scholar] [CrossRef]
- Downing, J.R.; Wilson, R.K.; Zhang, J.; Mardis, E.R.; Pui, C.-H.; Ding, L.; Ley, T.J.; Evans, W.E. The Pediatric Cancer Genome Project. Nat. Genet. 2012, 44, 619–622. [Google Scholar] [CrossRef]
- Meyer, C.; Larghero, P.; Almeida Lopes, B.; Burmeister, T.; Gröger, D.; Sutton, R.; Venn, N.C.; Cazzaniga, G.; Corral Abascal, L.; Tsaur, G.; et al. The KMT2A Recombinome of Acute Leukemias in 2023. Leukemia 2023, 37, 988–1005. [Google Scholar] [CrossRef]
- Valikhani, M.; Rahimian, E.; Ahmadi, S.E.; Chegeni, R.; Safa, M. Involvement of Classic and Alternative Non-Homologous End Joining Pathways in Hematologic Malignancies: Targeting Strategies for Treatment. Exp. Hematol. Oncol. 2021, 10, 51. [Google Scholar] [CrossRef]
- Zhang, F.; Gu, W.; Hurles, M.E.; Lupski, J.R. Copy Number Variation in Human Health, Disease, and Evolution. Annu. Rev. Genom. Hum. Genet. 2009, 10, 451–481. [Google Scholar] [CrossRef]
- Jeggo, P.A.; Geuting, V.; Löbrich, M. The Role of Homologous Recombination in Radiation-Induced Double-Strand Break Repair. Radiother. Oncol. 2011, 101, 7–12. [Google Scholar] [CrossRef]
- Averbeck, N.B.; Ringel, O.; Herrlitz, M.; Jakob, B.; Durante, M.; Taucher-Scholz, G. DNA End Resection Is Needed for the Repair of Complex Lesions in G1-Phase Human Cells. Cell Cycle 2014, 13, 2509–2516. [Google Scholar] [CrossRef]
- Jakob, B.; Dubiak-Szepietowska, M.; Janiel, E.; Schmidt, A.; Durante, M.; Taucher-Scholz, G. Differential Repair Protein Recruitment at Sites of Clustered and Isolated DNA Double-Strand Breaks Produced by High-Energy Heavy Ions. Sci. Rep. 2020, 10, 1443. [Google Scholar] [CrossRef]
- Rall, M.; Kraft, D.; Volcic, M.; Cucu, A.; Nasonova, E.; Taucher-Scholz, G.; Bönig, H.; Wiesmüller, L.; Fournier, C. Impact of Charged Particle Exposure on Homologous DNA Double-Strand Break Repair in Human Blood-Derived Cells. Front. Oncol. 2015, 5, 250. [Google Scholar] [CrossRef]
- Bender, M.A.; Gooch, P.C.; Kondo, S. The Gemini-3 S-4 Spaceflight-Radiation Interaction Experiment. Radiat. Res. 1967, 31, 91–111. [Google Scholar] [CrossRef]
- Bender, M.A.; Gooch, P.C.; Kondo, S. The Gemini XI S-4 Spaceflight-Radiation Interaction Experiment: The Human Blood Experiment. Radiat. Res. 1968, 34, 228–238. [Google Scholar] [CrossRef]
- Horneck, G. Impact of Microgravity on Radiobiological Processes and Efficiency of DNA Repair. Mutat. Res.—Fundam. Mol. Mech. Mutagen. 1999, 430, 221–228. [Google Scholar] [CrossRef]
- Wu, H.; George, K.; Willingham, V.; Cucinotta, F.A. Comparison of Chromosome Aberration Frequencies in Pre- and Post-Flight Astronaut Lymphocytes Irradiated in Vitro with Gamma Rays. Phys. Med. 2001, 17 (Suppl. S1), 229–231. [Google Scholar] [PubMed]
- Greco, O.; Durante, M.; Gialanella, G.; Grossi, G.; Pugliese, M.; Scampoli, P.; Snigiryova, G.; Obe, G. Biological Dosimetry in Russian and Italian Astronauts. Adv. Sp. Res. 2003, 31, 1495–1503. [Google Scholar] [CrossRef]
- Lu, T.; Zhang, Y.; Kidane, Y.; Feiveson, A.; Stodieck, L.; Karouia, F.; Ramesh, G.; Rohde, L.; Wu, H. Cellular Responses and Gene Expression Profile Changes Due to Bleomycin-Induced DNA Damage in Human Fibroblasts in Space. PLoS ONE 2017, 12, e0170358. [Google Scholar] [CrossRef] [PubMed]
- Risin, D.; Pellis, N.R. Modeled Microgravity Inhibits Apoptosis in Peripheral Blood Lymphocytes1. Vitr. Cell. Dev. Biol.—Anim. 2001, 37, 66. [Google Scholar] [CrossRef]
- Mosesso, P.; Schuber, M.; Seibt, D.; Schmitz, C.; Fiore, M.; Schinoppi, A.; Penna, S.; Palitti, F. X-Ray-Induced Chromosome Aberrations in Human Lymphocytes in Vitro Are Potentiated under Simulated Microgravity Conditions (Clinostat). Phys. Med. 2001, 17 (Suppl. S1), 264–266. [Google Scholar] [CrossRef] [PubMed]
- Canova, S.; Fiorasi, F.; Mognato, M.; Grifalconi, M.; Reddi, E.; Russo, A.; Celotti, L. “Modeled Microgravity” Affects Cell Response to Ionizing Radiation and Increases Genomic Damage. Radiat. Res. 2005, 163, 191–199. [Google Scholar] [CrossRef]
- Mognato, M.; Celotti, L. Modeled Microgravity Affects Cell Survival and HPRT Mutant Frequency, but Not the Expression of DNA Repair Genes in Human Lymphocytes Irradiated with Ionising Radiation. Mutat. Res. 2005, 578, 417–429. [Google Scholar] [CrossRef]
- Mognato, M.; Girardi, C.; Fabris, S.; Celotti, L. DNA Repair in Modeled Microgravity: Double Strand Break Rejoining Activity in Human Lymphocytes Irradiated with γ-Rays. Mutat. Res.—Fundam. Mol. Mech. Mutagen. 2009, 663, 32–39. [Google Scholar] [CrossRef]
- Kumari, R.; Singh, K.P.; DuMond, J.W. Simulated Microgravity Decreases DNA Repair Capacity and Induces DNA Damage in Human Lymphocytes. J. Cell. Biochem. 2009, 107, 723–731. [Google Scholar] [CrossRef]
- Manti, L.; Durante, M.; Cirrone, G.; Grossi, G.; Lattuada, M.; Pugliese, M.; Sabini, M.; Scampoli, P.; Valastro, L.; Gialanella, G. Modelled Microgravity Does Not Modify the Yield of Chromosome Aberrations Induced by High-Energy Protons in Human Lymphocytes. Int. J. Radiat. Biol. 2005, 81, 147–155. [Google Scholar] [CrossRef]
- Dang, B.; Yang, Y.; Zhang, E.; Li, W.; Mi, X.; Meng, Y.; Yan, S.; Wang, Z.; Wei, W.; Shao, C.; et al. Simulated Microgravity Increases Heavy Ion Radiation-Induced Apoptosis in Human B Lymphoblasts. Life Sci. 2014, 97, 123–128. [Google Scholar] [CrossRef]
- Tothova, Z.; Krill-Burger, J.M.; Popova, K.D.; Landers, C.C.; Sievers, Q.L.; Yudovich, D.; Belizaire, R.; Aster, J.C.; Morgan, E.A.; Tsherniak, A.; et al. Multiplex CRISPR/Cas9-Based Genome Editing in Human Hematopoietic Stem Cells Models Clonal Hematopoiesis and Myeloid Neoplasia. Cell Stem Cell 2017, 21, 547–555.e8. [Google Scholar] [CrossRef] [PubMed]
- Plett, P.A.; Abonour, R.; Frankovitz, S.M.; Orschell, C.M. Impact of Modeled Microgravity on Migration, Differentiation, and Cell Cycle Control of Primitive Human Hematopoietic Progenitor Cells. Exp. Hematol. 2004, 32, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Hauslage, J.; Cevik, V.; Hemmersbach, R. Pyrocystis Noctiluca Represents an Excellent Bioassay for Shear Forces Induced in Ground-Based Microgravity Simulators (Clinostat and Random Positioning Machine). NPJ Microgravity 2017, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Asaithamby, A.; Chen, D.J. Mechanism of Cluster DNA Damage Repair in Response to High-Atomic Number and Energy Particles Radiation. Mutat. Res. Mol. Mech. Mutagen. 2011, 711, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.J.; Chen, D.J. Complex DSBs: A Need for Resection. Cell Cycle 2014, 13, 3796–3797. [Google Scholar] [CrossRef]
- Mladenova, V.; Mladenov, E.; Stuschke, M.; Iliakis, G. DNA Damage Clustering after Ionizing Radiation and Consequences in the Processing of Chromatin Breaks. Molecules 2022, 27, 1540. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, X.; Wang, P.; Yu, X.; Essers, J.; Chen, D.; Kanaar, R.; Takeda, S.; Wang, Y. Characteristics of DNA-Binding Proteins Determine the Biological Sensitivity to High-Linear Energy Transfer Radiation. Nucleic Acids Res. 2010, 38, 3245–3251. [Google Scholar] [CrossRef]
- Hada, M.; Ikeda, H.; Rhone, J.R.; Beitman, A.J.; Plante, I.; Souda, H.; Yoshida, Y.; Held, K.D.; Fujiwara, K.; Saganti, P.B.; et al. Increased Chromosome Aberrations in Cells Exposed Simultaneously to Simulated Microgravity and Radiation. Int. J. Mol. Sci. 2019, 20, 43. [Google Scholar] [CrossRef]
- Yamanouchi, S.; Rhone, J.; Mao, J.H.; Fujiwara, K.; Saganti, P.B.; Takahashi, A.; Hada, M. Simultaneous Exposure of Cultured Human Lymphoblastic Cells to Simulated Microgravity and Radiation Increases Chromosome Aberrations. Life 2020, 10, 187. [Google Scholar] [CrossRef]
- Kraft, D.; Ritter, S.; Durante, M.; Seifried, E.; Fournier, C.; Tonn, T. Transmission of Clonal Chromosomal Abnormalities in Human Hematopoietic Stem and Progenitor Cells Surviving Radiation Exposure. Mutat. Res. 2015, 777, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Symington, L.S. Mechanism and Regulation of DNA End Resection in Eukaryotes. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 195–212. [Google Scholar] [CrossRef] [PubMed]
- Kais, Z.; Rondinelli, B.; Holmes, A.; O’Leary, C.; Kozono, D.; D’Andrea, A.D.; Ceccaldi, R. FANCD2 Maintains Fork Stability in BRCA1/2-Deficient Tumors and Promotes Alternative End-Joining DNA Repair. Cell Rep. 2016, 15, 2488–2499. [Google Scholar] [CrossRef] [PubMed]
- Raghunandan, M.; Chaudhury, I.; Kelich, S.L.; Hanenberg, H.; Sobeck, A. FANCD2, FANCJ and BRCA2 Cooperate to Promote Replication Fork Recovery Independently of the Fanconi Anemia Core Complex. Cell Cycle 2015, 14, 342–353. [Google Scholar] [CrossRef]
- Kanao, R.; Masutani, C. Regulation of DNA Damage Tolerance in Mammalian Cells by Post-Translational Modifications of PCNA. Mutat. Res. 2017, 803, 82–88. [Google Scholar] [CrossRef]
- Volcic, M.; Karl, S.; Baumann, B.; Salles, D.; Daniel, P.; Fulda, S.; Wiesmuller, L. NF-KappaB Regulates DNA Double-Strand Break Repair in Conjunction with BRCA1-CtIP Complexes. Nucleic Acids Res. 2012, 40, 181–195. [Google Scholar] [CrossRef]
- Kraft, D.; Rall, M.; Volcic, M.; Metzler, E.; Groo, A.; Stahl, A.; Bauer, L.; Nasonova, E.; Salles, D.; Taucher-Scholz, G.; et al. NF-KappaB-Dependent DNA Damage-Signaling Differentially Regulates DNA Double-Strand Break Repair Mechanisms in Immature and Mature Human Hematopoietic Cells. Leukemia 2015, 29, 1543–1554. [Google Scholar] [CrossRef]
- Zhang, Y.; Maria-Villanueva, M.; Krieger, S.; Ramesh, G.T.; Neelam, S.; Wu, H. Transcriptomics, NF-ΚB Pathway, and Their Potential Spaceflight-Related Health Consequences. Int. J. Mol. Sci. 2017, 18, 1166. [Google Scholar] [CrossRef]
- He, Y.; Pasupala, N.; Zhi, H.; Dorjbal, B.; Hussain, I.; Shih, H.M.; Bhattacharyya, S.; Biswas, R.; Miljkovic, M.; Semmes, O.J.; et al. NF-ΚB–Induced R-Loop Accumulation and DNA Damage Select for Nucleotide Excision Repair Deficiencies in Adult T Cell Leukemia. Proc. Natl. Acad. Sci. USA 2021, 118, e2005568118. [Google Scholar] [CrossRef]
- Ragu, S.; Droin, N.; Matos-Rodrigues, G.; Barascu, A.; Caillat, S.; Zarkovic, G.; Siberchicot, C.; Dardillac, E.; Gelot, C.; Guirouilh-Barbat, J.; et al. A Noncanonical Response to Replication Stress Protects Genome Stability through ROS Production, in an Adaptive Manner. Cell Death Differ. 2023, 30, 1349–1365. [Google Scholar] [CrossRef]
- Rithidech, K.N.; Honikel, L.; Whorton, E.B. MFISH Analysis of Chromosomal Damage in Bone Marrow Cells Collected from CBA/CaJ Mice Following Whole Body Exposure to Heavy Ions (56Fe Ions). Radiat. Environ. Biophys. 2007, 46, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Rajput, M.; Singh, R.P. Simulated Microgravity Triggers DNA Damage and Mitochondria-Mediated Apoptosis through ROS Generation in Human Promyelocytic Leukemic Cells. Mitochondrion 2021, 61, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Löbrich, M.; Jeggo, P. A Process of Resection-Dependent Nonhomologous End Joining Involving the Goddess Artemis. Trends Biochem. Sci. 2017, 42, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Bartosova, Z.; Krejci, L. Nucleases in Homologous Recombination as Targets for Cancer Therapy. FEBS Lett. 2014, 588, 2446–2456. [Google Scholar] [CrossRef]
- Trenner, A.; Sartori, A.A. Harnessing DNA Double-Strand Break Repair for Cancer Treatment. Front. Oncol. 2019, 9, 1388. [Google Scholar] [CrossRef]
- Murina, O.; von Aesch, C.; Karakus, U.; Ferretti, L.P.; Bolck, H.A.; Hänggi, K.; Sartori, A.A. FANCD2 and CtIP Cooperate to Repair DNA Interstrand Crosslinks. Cell Rep. 2014, 7, 1030–1038. [Google Scholar] [CrossRef]
- Claesson, K.; Magnander, K.; Kahu, H.; Lindegren, S.; Hultborn, R.; Elmroth, K. RBE of α-Particles from 211At for Complex DNA Damage and Cell Survival in Relation to Cell Cycle Position. Int. J. Radiat. Biol. 2011, 87, 372–384. [Google Scholar] [CrossRef]
- Pinto, M.; Prise, K.M.; Michael, B.D. Evidence for Complexity at the Nanometer Scale of Radiation-Induced DNA DSBs as a Determinant of Rejoining Kinetics. Radiat. Res. 2005, 164, 73–85. [Google Scholar] [CrossRef]
- Anderson, J.A.; Harper, J.V.; Cucinotta, F.A.; O’Neill, P. Participation of DNA-PKcs in DSB Repair after Exposure to High-and Low-Let Radiation. Radiat. Res. 2010, 174, 195–205. [Google Scholar] [CrossRef]
- Nickoloff, J.A.; Sharma, N.; Taylor, L. Clustered DNA Double-Strand Breaks: Biological Effects and Relevance to Cancer Radiotherapy. Genes. 2020, 11, 99. [Google Scholar] [CrossRef]
- Panier, S.; Boulton, S.J. Double-Strand Break Repair: 53BP1 Comes into Focus. Nat. Rev. Mol. Cell Biol. 2014, 15, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Elsasser, T.; Tonn, T.; Seifried, E.; Durante, M.; Ritter, S.; Fournier, C. Response of Human Hematopoietic Stem and Progenitor Cells to Energetic Carbon Ions. Int. J. Radiat. Biol. 2009, 85, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Ritter, S.; Durante, M. Heavy-Ion Induced Chromosomal Aberrations: A Review. Mutat. Res. 2010, 701, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Iliakis, G.; Mladenov, E.; Mladenova, V. Necessities in the Processing of DNA Double Strand Breaks and Their Effects on Genomic Instability and Cancer. Cancers 2019, 11, 1671. [Google Scholar] [CrossRef]
- Cornforth, M.N. Occam’s Broom and the Dirty DSB: Cytogenetic Perspectives on Cellular Response to Changes in Track Structure and Ionization Density. Int. J. Radiat. Biol. 2021, 97, 1099–1108. [Google Scholar] [CrossRef]
- Mangala, L.S.; Zhang, Y.; He, Z.; Emami, K.; Ramesh, G.T.; Story, M.; Rohde, L.H.; Wu, H. Effects of Simulated Microgravity on Expression Profile of MicroRNA in Human Lymphoblastoid Cells. J. Biol. Chem. 2011, 286, 32483–32490. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, T.; Wong, M.; Wang, X.; Stodieck, L.; Karouia, F.; Story, M.; Wu, H. Transient Gene and MicroRNA Expression Profile Changes of Confluent Human Fibroblast Cells in Spaceflight. FASEB J. 2016, 30, 2211–2224. [Google Scholar] [CrossRef]
- Zwart, S.R.; Pierson, D.; Mehta, S.; Gonda, S.; Smith, S.M. Capacity of Omega-3 Fatty Acids or Eicosapentaenoic Acid to Counteract Weightlessness-Induced Bone Loss by Inhibiting NF-ΚB Activation: From Cells to Bed Rest to Astronauts. J. Bone Miner. Res. 2010, 25, 1049–1057. [Google Scholar] [CrossRef]
- Chang, T.T.; Walther, I.; Li, C.-F.; Boonyaratanakornkit, J.; Galleri, G.; Meloni, M.A.; Pippia, P.; Cogoli, A.; Hughes-Fulford, M. The Rel/NF-ΚB Pathway and Transcription of Immediate Early Genes in T Cell Activation Are Inhibited by Microgravity. J. Leukoc. Biol. 2012, 92, 1133–1145. [Google Scholar] [CrossRef]
- Boonyaratanakornkit, J.B.; Cogoli, A.; Li, C.-F.; Schopper, T.; Pippia, P.; Galleri, G.; Meloni, M.A.; Hughes-Fulford, M. Key Gravity-sensitive Signaling Pathways Drive T-cell Activation. FASEB J. 2005, 19, 2020–2022. [Google Scholar] [CrossRef]
- Paulsen, K.; Thiel, C.; Timm, J.; Schmidt, P.M.; Huber, K.; Tauber, S.; Hemmersbach, R.; Seibt, D.; Kroll, H.; Grote, K.H.; et al. Microgravity-Induced Alterations in Signal Transduction in Cells of the Immune System. Acta Astronaut. 2010, 67, 1116–1125. [Google Scholar] [CrossRef]
- Horneck, G.; Rettberg, P.; Kozubek, S.; Baumstark-Khan, C.; Rink, H.; Schäfer, M.; Schmitz, C. The Influence of Microgravity on Repair of Radiation-Induced DNA Damage in Bacteria and Human Fibroblasts. Radiat. Res. 1997, 147, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Berti, M.; Cortez, D.; Lopes, M. The Plasticity of DNA Replication Forks in Response to Clinically Relevant Genotoxic Stress. Nat. Rev. Mol. Cell Biol. 2020, 21, 633–651. [Google Scholar] [CrossRef]
- Ihle, M.; Biber, S.; Schroeder, I.S.; Blattner, C.; Deniz, M.; Damia, G.; Gottifredi, V.; Wiesmuller, L. Impact of the Interplay between Stemness Features, P53 and Pol Iota on Replication Pathway Choices. Nucleic Acids Res. 2021, 49, 7457–7475. [Google Scholar] [CrossRef]
- Schlacher, K.; Wu, H.; Jasin, M. A Distinct Replication Fork Protection Pathway Connects Fanconi Anemia Tumor Suppressors to RAD51-BRCA1/2. Cancer Cell 2012, 22, 106–116. [Google Scholar] [CrossRef]
- Petropoulos, M.; Champeris Tsaniras, S.; Taraviras, S.; Lygerou, Z. Replication Licensing Aberrations, Replication Stress, and Genomic Instability. Trends Biochem. Sci. 2019, 44, 752–764. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Li, R.; Tan, X.; Zhang, J.; Fan, C.; Zhao, Q.; Deng, Y.; Xu, A.; Lukong, K.E.; Genth, H.; et al. Simulated Microgravity Reduces Focal Adhesions and Alters Cytoskeleton and Nuclear Positioning Leading to Enhanced Apoptosis via Suppressing FAK/Rhoa-Mediated MTORC1/NF-ΚB and ERK1/2 Pathways. Int. J. Mol. Sci. 2018, 19, 1994. [Google Scholar] [CrossRef] [PubMed]
- Mahen, R.; Hattori, H.; Lee, M.; Sharma, P.; Jeyasekharan, A.D.; Venkitaraman, A.R. A-Type Lamins Maintain the Positional Stability of DNA Damage Repair Foci in Mammalian Nuclei. PLoS ONE 2013, 8, e61893. [Google Scholar] [CrossRef]
- Simon, M.; Yang, J.; Gigas, J.; Earley, E.J.; Hillpot, E.; Zhang, L.; Zagorulya, M.; Tombline, G.; Gilbert, M.; Yuen, S.L.; et al. A Rare Human Centenarian Variant of SIRT6 Enhances Genome Stability and Interaction with Lamin A. EMBO J. 2022, 41, e110393. [Google Scholar] [CrossRef]
- Barravecchia, I.; De Cesari, C.; Forcato, M.; Scebba, F.; Pyankova, O.V.; Bridger, J.M.; Foster, H.A.; Signore, G.; Borghini, A.; Andreassi, M.; et al. Microgravity and Space Radiation Inhibit Autophagy in Human Capillary Endothelial Cells, through Either Opposite or Synergistic Effects on Specific Molecular Pathways. Cell. Mol. Life Sci. 2022, 79, 28. [Google Scholar] [CrossRef]
- Schreck, R.; Rieber, P.; Baeuerle, P.A. Reactive Oxygen Intermediates as Apparently Widely Used Messengers in the Activation of the NF-ΚB Transcription Factor and HIV-1. EMBO J. 1991, 10, 2247–2258. [Google Scholar] [CrossRef]
- Albi, E.; Krüger, M.; Hemmersbach, R.; Lazzarini, A.; Cataldi, S.; Codini, M.; Beccari, T.; Ambesi-Impiombato, F.S.; Curcio, F. Impact of Gravity on Thyroid Cells. Int. J. Mol. Sci. 2017, 18, 972. [Google Scholar] [CrossRef] [PubMed]
- Eiermann, P.; Kopp, S.; Hauslage, J.; Hemmersbach, R.; Gerzer, R.; Ivanova, K. Adaptation of a 2-D Clinostat for Simulated Microgravity Experiments with Adherent Cells. Microgravity Sci. Technol. 2013, 25, 153–159. [Google Scholar] [CrossRef]
- Kamentsky, L.; Jones, T.R.; Fraser, A.; Bray, M.-A.; Logan, D.J.; Madden, K.L.; Ljosa, V.; Rueden, C.; Eliceiri, K.W.; Carpenter, A.E. Improved Structure, Function and Compatibility for CellProfiler: Modular High-Throughput Image Analysis Software. Bioinformatics 2011, 27, 1179–1180. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M. Cytokinesis-Block Micronucleus Cytome Assay. Nat. Protoc. 2007, 2, 1084–1104. [Google Scholar] [CrossRef]
- Rall-Scharpf, M.; Friedl, T.W.P.; Biechonski, S.; Denkinger, M.; Milyavsky, M.; Wiesmüller, L. Sex-Specific Differences in DNA Double-Strand Break Repair of Cycling Human Lymphocytes during Aging. Aging 2021, 13, 21066–21089. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Kim, H.; Kim, J.M.; Primack, B.; Vidal-Cardenas, S.; Xu, Y.; Price, B.D.; Mills, A.A.; D’Andrea, A.D. FANCD2 Activates Transcription of TAp63 and Suppresses Tumorigenesis. Mol. Cell 2013, 50, 908–918. [Google Scholar] [CrossRef] [PubMed]
- Helbling-Leclerc, A.; Dessarps-Freichey, F.; Evrard, C.; Rosselli, F. Fanconi Anemia Proteins Counteract the Implementation of the Oncogene-Induced Senescence Program. Sci. Rep. 2019, 9, 17024. [Google Scholar] [CrossRef]
- Flach, J.; Bakker, S.T.; Mohrin, M.; Conroy, P.C.; Pietras, E.M.; Reynaud, D.; Alvarez, S.; Diolaiti, M.E.; Ugarte, F.; Forsberg, E.C.; et al. Replication Stress Is a Potent Driver of Functional Decline in Ageing Haematopoietic Stem Cells. Nature 2014, 512, 198–202. [Google Scholar] [CrossRef]
- Walter, D.; Lier, A.; Geiselhart, A.; Thalheimer, F.B.; Huntscha, S.; Sobotta, M.C.; Moehrle, B.; Brocks, D.; Bayindir, I.; Kaschutnig, P.; et al. Exit from Dormancy Provokes DNA-Damage-Induced Attrition in Haematopoietic Stem Cells. Nature 2015, 520, 549–552. [Google Scholar] [CrossRef]
- Sotnezova, E.V.; Markina, E.A.; Andreeva, E.R.; Buravkova, L.B. Myeloid Precursors in the Bone Marrow of Mice after a 30-Day Space Mission on a Bion-M1 Biosatellite. Bull. Exp. Biol. Med. 2017, 162, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Boerma, M.; Rodriguez, D.; Campbell-Beachler, M.; Jones, T.; Stanbouly, S.; Sridharan, V.; Nishiyama, N.C.; Wroe, A.; Nelson, G.A. Combined Effects of Low-Dose Proton Radiation and Simulated Microgravity on the Mouse Retina and the Hematopoietic System. Radiat. Res. 2019, 192, 241–250. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Handwerk, L.; Schreier, H.K.; Kraft, D.; Shreder, K.; Hemmersbach, R.; Hauslage, J.; Bonig, H.; Wiesmüller, L.; Fournier, C.; Rall-Scharpf, M. Simulating Space Conditions Evokes Different DNA Damage Responses in Immature and Mature Cells of the Human Hematopoietic System. Int. J. Mol. Sci. 2023, 24, 13761. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241813761
Handwerk L, Schreier HK, Kraft D, Shreder K, Hemmersbach R, Hauslage J, Bonig H, Wiesmüller L, Fournier C, Rall-Scharpf M. Simulating Space Conditions Evokes Different DNA Damage Responses in Immature and Mature Cells of the Human Hematopoietic System. International Journal of Molecular Sciences. 2023; 24(18):13761. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241813761
Chicago/Turabian StyleHandwerk, Leonie, Heike Katrin Schreier, Daniela Kraft, Kateryna Shreder, Ruth Hemmersbach, Jens Hauslage, Halvard Bonig, Lisa Wiesmüller, Claudia Fournier, and Melanie Rall-Scharpf. 2023. "Simulating Space Conditions Evokes Different DNA Damage Responses in Immature and Mature Cells of the Human Hematopoietic System" International Journal of Molecular Sciences 24, no. 18: 13761. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms241813761