
Transcripts of the Prostate Cancer-Associated Gene ANO7 Are Retained in the Nuclei of Prostatic Epithelial Cells
 , ,
, ,
Abstract
:1. Introduction
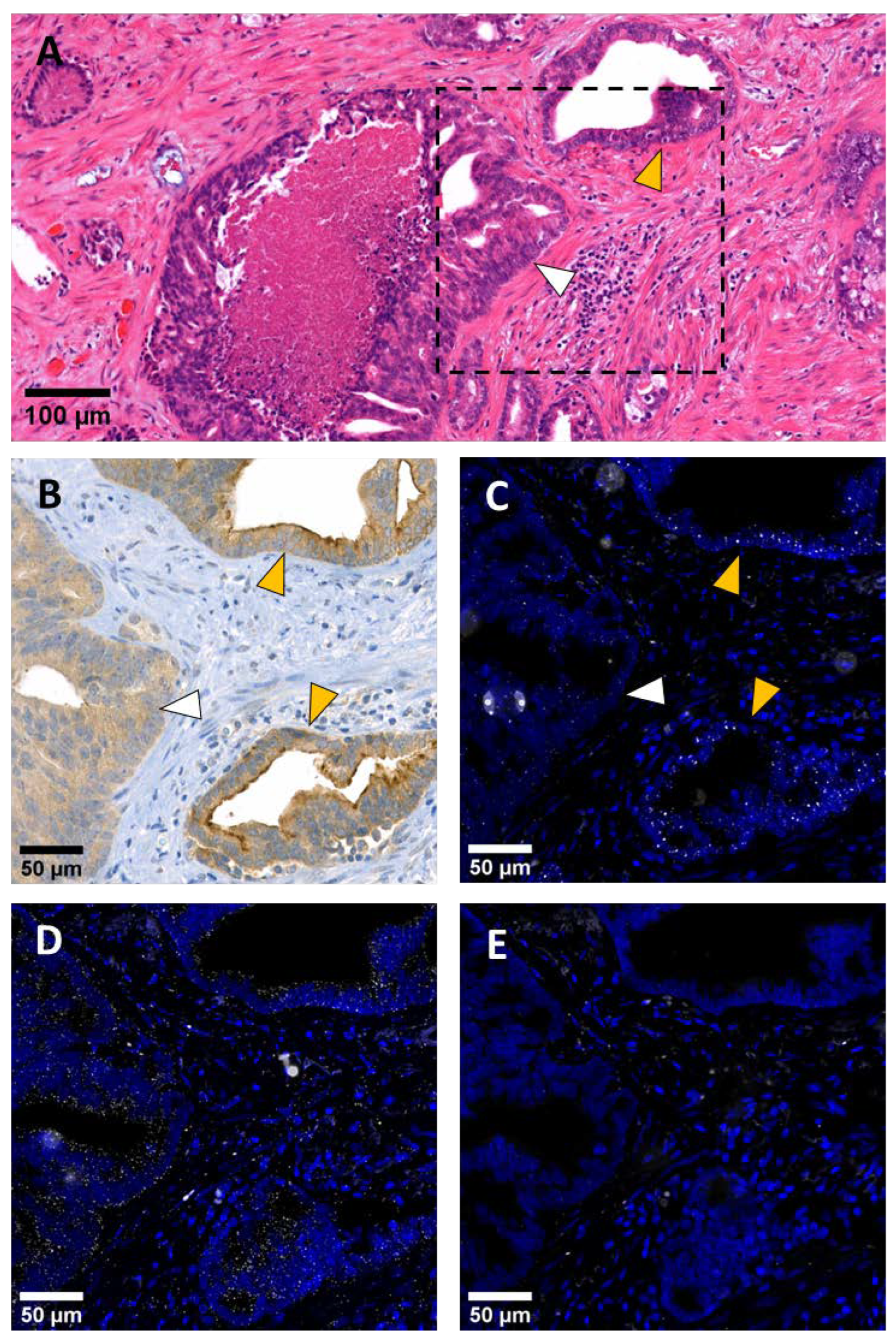
2. Results
2.1. ANO7 Transcripts Are Expressed in Luminal Cells of the Prostatic Epithelium
2.2. ANO7 Transcripts Are Enriched in the Nucleus of Benign Prostate Epithelial Cells and Cancer Cells
2.3. ANO7 Transcripts and the Nucleus-Enriched MALAT1 Are Distributed Similarly between Nuclear and Cytoplasmic Compartments in Prostate Cancer Cell Lines
3. Discussion
4. Materials and Methods
4.1. Patient Samples
4.2. Fluorescent In Situ Hybridization
4.3. Immunohistochemistry
4.4. Microscopy
4.5. Image Analysis
4.6. Cell Culture
4.7. Fractionation
4.8. Western Blot
4.9. RNA Isolation
4.10. ddPCR
4.11. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Mucci, L.A.; Hjelmborg, J.B.; Harris, J.R.; Czene, K.; Havelick, D.J.; Scheike, T.; Graff, R.E.; Holst, K.; Möller, S.; Unger, R.H.; et al. Familial Risk and Heritability of Cancer Among Twins in Nordic Countries. JAMA 2016, 315, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Conti, D.V.; Darst, B.F.; Moss, L.C.; Saunders, E.J.; Sheng, X.; Chou, A.; Schumacher, F.R.; Olama, A.A.A.; Benlloch, S.; Dadaev, T.; et al. Trans-Ancestry Genome-Wide Association Meta-Analysis of Prostate Cancer Identifies New Susceptibility Loci and Informs Genetic Risk Prediction. Nat. Genet. 2021, 53, 65–75. [Google Scholar] [CrossRef]
- Kaikkonen, E.; Rantapero, T.; Zhang, Q.; Taimen, P.; Laitinen, V.; Kallajoki, M.; Jambulingam, D.; Ettala, O.; Knaapila, J.; Boström, P.J.; et al. ANO7 Is Associated with Aggressive Prostate Cancer. Int. J. Cancer 2018, 143, 2479–2487. [Google Scholar] [CrossRef]
- Cropp, C.D.; Simpson, C.L.; Wahlfors, T.; Ha, N.; George, A.; Jones, M.S.; Harper, U.; Ponciano-Jackson, D.; Green, T.A.; Tammela, T.L.J.; et al. Genome-Wide Linkage Scan for Prostate Cancer Susceptibility in Finland: Evidence for a Novel Locus on 2q37.3 and Confirmation of Signal on 17q21-q22. Int. J. Cancer J. 2011, 129, 2400–2407. [Google Scholar] [CrossRef] [Green Version]
- Kalienkova, V.; Clerico Mosina, V.; Paulino, C. The Groovy TMEM16 Family: Molecular Mechanisms of Lipid Scrambling and Ion Conduction. J. Mol. Biol. 2021, 433, 166941. [Google Scholar] [CrossRef]
- Bera, T.K.; Das, S.; Maeda, H.; Beers, R.; Wolfgang, C.D.; Kumar, V.; Hahn, Y.; Lee, B.; Pastan, I. NGEP, a Gene Encoding a Membrane Protein Detected Only in Prostate Cancer and Normal Prostate. Proc. Natl. Acad. Sci. USA 2004, 101, 3059–3064. [Google Scholar] [CrossRef] [Green Version]
- Katoh, M.; Katoh, M. Characterization of Human TMEM16G Gene in Silico. Int. J. Mol. Med. 2004, 14, 759–764. [Google Scholar] [CrossRef]
- Guo, J.; Wang, D.; Dong, Y.; Gao, X.; Tong, H.; Liu, W.; Zhang, L.; Sun, M. ANO7: Insights into Topology, Function, and Potential Applications as a Biomarker and Immunotherapy Target. Tissue Cell 2021, 72, 101546. [Google Scholar] [CrossRef]
- Suzuki, J.; Fujii, T.; Imao, T.; Ishihara, K.; Kuba, H.; Nagata, S. Calcium-Dependent Phospholipid Scramblase Activity of TMEM16 Protein Family Members. J. Biol. Chem. 2013, 288, 13305–13316. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, R.; Uliyakina, I.; Kongsuphol, P.; Warth, R.; Mirza, M.; Martins, J.R.; Kunzelmann, K. Expression and Function of Epithelial Anoctamins. J. Biol. Chem. 2010, 285, 7838–7845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Schreiber, R.; Kunzelmann, K. Anoctamins Are a Family of Ca2+-Activated Cl− Channels. J. Cell Sci. 2012, 125, 4991–4998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jhun, M.A.; Geybels, M.S.; Wright, J.L.; Kolb, S.; April, C.; Bibikova, M.; Ostrander, E.A.; Fan, J.-B.; Feng, Z.; Stanford, J.L. Gene Expression Signature of Gleason Score Is Associated with Prostate Cancer Outcomes in a Radical Prostatectomy Cohort. Oncotarget 2017, 8, 43035–43047. [Google Scholar] [CrossRef] [PubMed]
- Sinnott, J.A.; Peisch, S.F.; Tyekucheva, S.; Gerke, T.; Lis, R.; Rider, J.R.; Fiorentino, M.; Stampfer, M.J.; Mucci, L.A.; Loda, M.; et al. Prognostic Utility of a New mRNA Expression Signature of Gleason Score. Clin. Cancer Res. 2017, 23, 81–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, S.M.; Knight, L.A.; McCavigan, A.M.; Logan, G.E.; Berge, V.; Sherif, A.; Pandha, H.; Warren, A.Y.; Davidson, C.; Uprichard, A.; et al. Molecular Subgroup of Primary Prostate Cancer Presenting with Metastatic Biology. Eur. Urol. 2017, 72, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Chandran, U.R.; Ma, C.; Dhir, R.; Bisceglia, M.; Lyons-Weiler, M.; Liang, W.; Michalopoulos, G.; Becich, M.; Monzon, F.A. Gene Expression Profiles of Prostate Cancer Reveal Involvement of Multiple Molecular Pathways in the Metastatic Process. BMC Cancer 2007, 7, 64. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Hahn, Y.; Walker, D.A.; Nagata, S.; Willingham, M.C.; Peehl, D.M.; Bera, T.K.; Lee, B.; Pastan, I. Topology of NGEP, a Prostate-Specific Cell:Cell Junction Protein Widely Expressed in Many Cancers of Different Grade Level. Cancer Res. 2008, 68, 6306–6312. [Google Scholar] [CrossRef] [Green Version]
- Mohsenzadegan, M.; Madjd, Z.; Asgari, M.; Abolhasani, M.; Shekarabi, M.; Taeb, J.; Shariftabrizi, A. Reduced Expression of NGEP Is Associated with High-Grade Prostate Cancers: A Tissue Microarray Analysis. Cancer Immunol. Immunother. 2013, 62, 1609–1618. [Google Scholar] [CrossRef]
- Marx, A.; Koopmann, L.; Höflmayer, D.; Büscheck, F.; Hube-Magg, C.; Steurer, S.; Eichenauer, T.; Clauditz, T.S.; Wilczak, W.; Simon, R.; et al. Reduced Anoctamin 7 (ANO7) Expression Is a Strong and Independent Predictor of Poor Prognosis in Prostate Cancer. Cancer Biol. Med. 2021, 18, 245–255. [Google Scholar] [CrossRef]
- Henry, G.H.; Malewska, A.; Joseph, D.B.; Malladi, V.S.; Lee, J.; Torrealba, J.; Mauck, R.J.; Gahan, J.C.; Raj, G.V.; Roehrborn, C.G.; et al. A Cellular Anatomy of the Normal Adult Human Prostate and Prostatic Urethra. Cell Rep. 2018, 25, 3530–3542.e5. [Google Scholar] [CrossRef]
- Kellokumpu-Lehtinen, P.-L.; Harmenberg, U.; Joensuu, T.; McDermott, R.; Hervonen, P.; Ginman, C.; Luukkaa, M.; Nyandoto, P.; Hemminki, A.; Nilsson, S.; et al. 2-Weekly versus 3-Weekly Docetaxel to Treat Castration-Resistant Advanced Prostate Cancer: A Randomised, Phase 3 Trial. Lancet Oncol. 2013, 14, 117–124. [Google Scholar] [CrossRef]
- Wang, F.; Flanagan, J.; Su, N.; Wang, L.-C.; Bui, S.; Nielson, A.; Wu, X.; Vo, H.-T.; Ma, X.-J.; Luo, Y. RNAscope. J. Mol. Diagn. JMD 2012, 14, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Hahn, Y.; Nagata, S.; Willingham, M.C.; Bera, T.K.; Lee, B.; Pastan, I. NGEP, a Prostate-Specific Plasma Membrane Protein That Promotes the Association of LNCaP Cells. Cancer Res. 2007, 67, 1594–1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildyard, J.C.W.; Rawson, F.; Wells, D.J.; Piercy, R.J. Multiplex in Situ Hybridization within a Single Transcript: RNAscope Reveals Dystrophin mRNA Dynamics. PLoS ONE 2020, 15, e0239467. [Google Scholar] [CrossRef] [PubMed]
- Hammerich, K.H.; Ayala, G.E.; Wheeler, T.M. Anatomy of the Prostate Gland and Surgical Pathology of Prostate Cancer. In Prostate Cancer; Hricak, H., Scardino, P., Eds.; Contemporary Issues in Cancer Imaging; Cambridge University Press: Cambridge, UK, 2008; pp. 1–14. ISBN 978-0-521-88704-5. [Google Scholar]
- Cabili, M.N.; Dunagin, M.C.; McClanahan, P.D.; Biaesch, A.; Padovan-Merhar, O.; Regev, A.; Rinn, J.L.; Raj, A. Localization and Abundance Analysis of Human lncRNAs at Single-Cell and Single-Molecule Resolution. Genome Biol. 2015, 16, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samacoits, A.; Chouaib, R.; Safieddine, A.; Traboulsi, A.-M.; Ouyang, W.; Zimmer, C.; Peter, M.; Bertrand, E.; Walter, T.; Mueller, F. A Computational Framework to Study Sub-Cellular RNA Localization. Nat. Commun. 2018, 9, 4584. [Google Scholar] [CrossRef] [Green Version]
- Wahlström, G.; Heron, S.; Knuuttila, M.; Kaikkonen, E.; Tulonen, N.; Metsälä, O.; Löf, C.; Ettala, O.; Boström, P.J.; Taimen, P.; et al. The Variant rs77559646 Associated with Aggressive Prostate Cancer Disrupts ANO7 mRNA Splicing and Protein Expression. Hum. Mol. Genet. 2022, 31, 2063–2077. [Google Scholar] [CrossRef]
- Sramkoski, R.M.; Pretlow, T.G.; Giaconia, J.M.; Pretlow, T.P.; Schwartz, S.; Sy, M.-S.; Marengo, S.R.; Rhim, J.S.; Zhang, D.; Jacobberger, J.W. A New Human Prostate Carcinoma Cell Line, 22Rv1. Vitro Cell. Dev. Biol.-Anim. 1999, 35, 403–409. [Google Scholar] [CrossRef]
- Navone, N.M.; Olive, M.; Ozen, M.; Davis, R.; Troncoso, P.; Tu, S.M.; Johnston, D.; Pollack, A.; Pathak, S.; von Eschenbach, A.C.; et al. Establishment of Two Human Prostate Cancer Cell Lines Derived from a Single Bone Metastasis. Clin. Cancer Res. 1997, 3, 2493–2500. [Google Scholar]
- Popp, M.W.; Maquat, L.E. Nonsense-Mediated mRNA Decay and Cancer. Curr. Opin. Genet. Dev. 2018, 48, 44–50. [Google Scholar] [CrossRef]
- Zaghlool, A.; Niazi, A.; Björklund, Å.K.; Westholm, J.O.; Ameur, A.; Feuk, L. Characterization of the Nuclear and Cytosolic Transcriptomes in Human Brain Tissue Reveals New Insights into the Subcellular Distribution of RNA Transcripts. Sci. Rep. 2021, 11, 4076. [Google Scholar] [CrossRef] [PubMed]
- Wegener, M.; Müller-McNicoll, M. Nuclear Retention of mRNAs–Quality Control, Gene Regulation and Human Disease. Semin. Cell Dev. Biol. 2018, 79, 131–142. [Google Scholar] [CrossRef]
- Bahar Halpern, K.; Caspi, I.; Lemze, D.; Levy, M.; Landen, S.; Elinav, E.; Ulitsky, I.; Itzkovitz, S. Nuclear Retention of mRNA in Mammalian Tissues. Cell Rep. 2015, 13, 2653–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, J.M.; Phizicky, D.V.; Neugebauer, K.M. Nuclear Mechanisms of Gene Expression Control: Pre-mRNA Splicing as a Life or Death Decision. Curr. Opin. Genet. Dev. 2021, 67, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Palazzo, A.F.; Lee, E.S. Sequence Determinants for Nuclear Retention and Cytoplasmic Export of mRNAs and lncRNAs. Front. Genet. 2018, 9, 440. [Google Scholar] [CrossRef] [Green Version]
- Braunschweig, U.; Barbosa-Morais, N.L.; Pan, Q.; Nachman, E.N.; Alipanahi, B.; Gonatopoulos-Pournatzis, T.; Frey, B.; Irimia, M.; Blencowe, B.J. Widespread Intron Retention in Mammals Functionally Tunes Transcriptomes. Genome Res. 2014, 24, 1774–1786. [Google Scholar] [CrossRef] [Green Version]
- Dvinge, H.; Bradley, R.K. Widespread Intron Retention Diversifies Most Cancer Transcriptomes. Genome Med. 2015, 7, 45. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.J.-L.; Schmitz, U. Intron Retention: Importance, Challenges, and Opportunities. Trends Genet. 2022, 38, 789–792. [Google Scholar] [CrossRef]
- Cereda, V.; Poole, D.J.; Palena, C.; Das, S.; Bera, T.K.; Remondo, C.; Gulley, J.L.; Arlen, P.M.; Yokokawa, J.; Pastan, I.; et al. New Gene Expressed in Prostate (NGEP): A Potential Target for T Cell-Mediated Prostate Cancer Immunotherapy. Cancer Immunol. Immunother. CII 2010, 59, 63–71. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nuclear % | SD | SEM | n Subjects 1 | n ROIs | n Cells | n Spots 2 | |
|---|---|---|---|---|---|---|---|
| Benign glands and low-grade cancer | |||||||
| ANO7 | 88.8 | 3.8 | 1.4 | 5 | 7 | 644 | 1163 |
| PPIB | 47.7 | 6.3 | 2.4 | 5 | 7 | 608 | 4337 |
| dabB | 67.4 | 20.7 | 9.3 | 4 | 5 | 280 | 53 |
| High-grade cancer | |||||||
| ANO7 | 77.7 | 9.3 | 3.5 | 5 | 7 | 817 | 975 |
| PPIB | 46.7 | 10.8 | 4.1 | 5 | 7 | 834 | 8985 |
| dabB | 57.8 | 23.7 | 10.6 | 4 | 5 | 437 | 228 |
| Cytoplasm | Nucleus | Whole Cell | Nuclear Enrichment | ||||
|---|---|---|---|---|---|---|---|
| Copies per 1 µg RNA | SD | Copies per 1 µg RNA | SD | Copies per 1 µg RNA | SD | Log2 Nucleus/Cytoplasm | |
| 22Rv1 | |||||||
| ANO7 | 117 | 30 | 1616 | 690 | 479 | 134 | 3.75 |
| MALAT1 | 1.0 × 106 | 0.4 × 106 | 15.2 × 106 | 2.9 × 106 | 10.1 × 106 | 5.6 × 106 | 3.93 |
| GAPDH | 5.6 × 106 | 1.8 × 106 | 1.3 × 106 | 0.4 × 106 | 7.2 × 106 | 2.3 × 106 | −2.07 |
| MDA PCa 2b | |||||||
| ANO7 | 3.6 × 103 | 1.7 × 103 | 22.0 × 103 | 9.5 × 103 | 6.5 × 103 | 2.8 × 103 | 2.63 |
| MALAT1 | 9.4 × 106 | 2.4 × 106 | 102.7 × 106 | 27.6 × 106 | 30.1 × 106 | 2.6 × 106 | 3.44 |
| GAPDH | 41.8 × 105 | 14.7 × 105 | 7.3 × 105 | 2.1 × 105 | 25.5 × 105 | 2.1 × 105 | −2.52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metsälä, O.; Wahlström, G.; Taimen, P.; Kellokumpu-Lehtinen, P.-L.; Schleutker, J. Transcripts of the Prostate Cancer-Associated Gene ANO7 Are Retained in the Nuclei of Prostatic Epithelial Cells. Int. J. Mol. Sci. 2023, 24, 1052. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021052
Metsälä O, Wahlström G, Taimen P, Kellokumpu-Lehtinen P-L, Schleutker J. Transcripts of the Prostate Cancer-Associated Gene ANO7 Are Retained in the Nuclei of Prostatic Epithelial Cells. International Journal of Molecular Sciences. 2023; 24(2):1052. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021052
Chicago/Turabian StyleMetsälä, Olli, Gudrun Wahlström, Pekka Taimen, Pirkko-Liisa Kellokumpu-Lehtinen, and Johanna Schleutker. 2023. "Transcripts of the Prostate Cancer-Associated Gene ANO7 Are Retained in the Nuclei of Prostatic Epithelial Cells" International Journal of Molecular Sciences 24, no. 2: 1052. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021052