
CD8 and CD4 Positive NKT Subpopulations and Immune-Checkpoint Pathways in Early-Onset Preeclampsia and Healthy Pregnancy

Abstract
:1. Introduction
2. Results
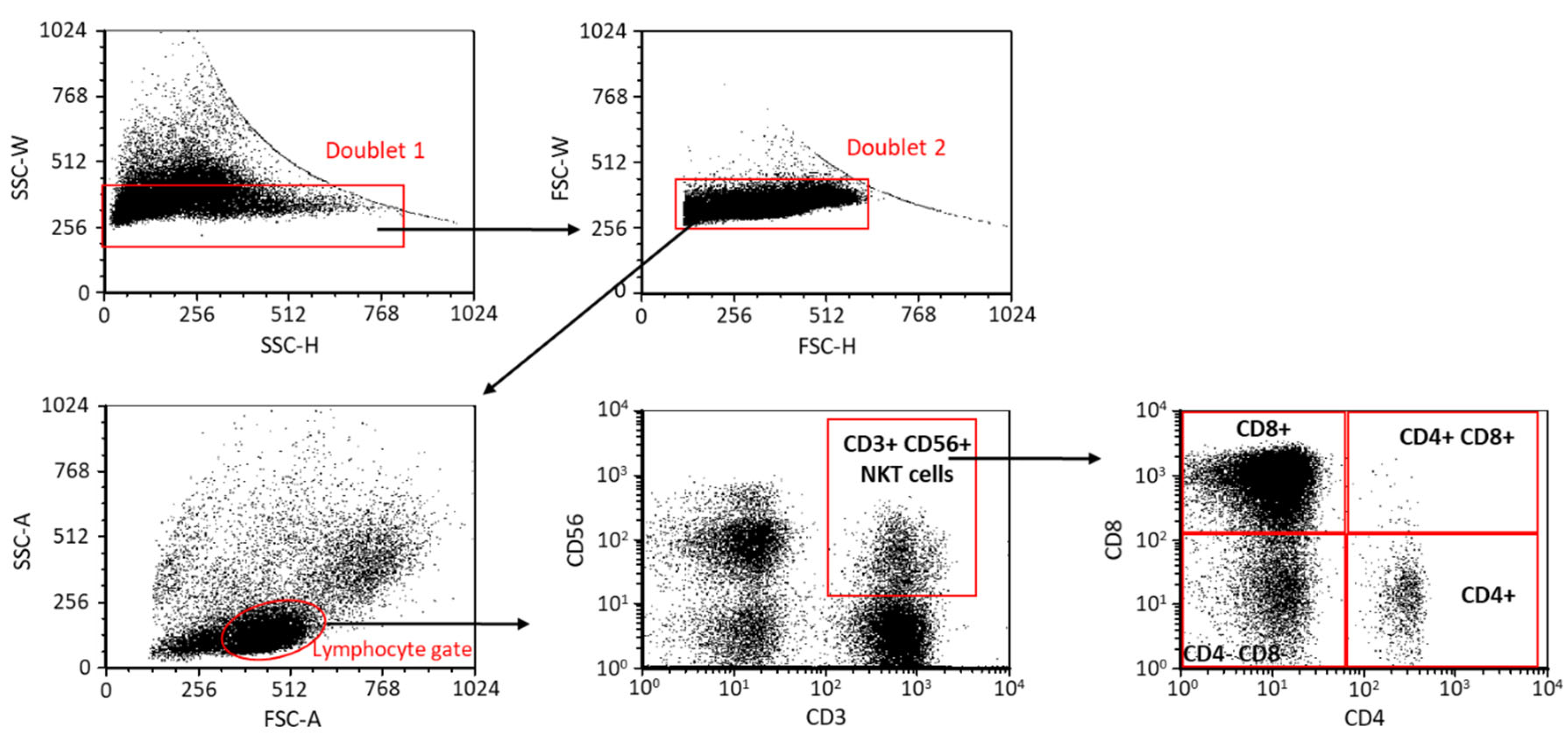
2.1. The Frequency of NKT Cell Subpopulations in EOP Patients and Healthy Pregnant Women
2.2. PD-1 Receptor and PD-L1 Ligand Expression by the Four NKT Cell Subpopulations in EOP Patients and Healthy Pregnant Women
2.3. LAG-3 Receptor and Gal-3 Ligand Expression by the Four NKT Cell Subpopulations in EOP Patients and Healthy Pregnant Women
2.4. TIGIT and CD226 Receptor Expression by the Four NKT Cell Subpopulations in EOP Patients and Healthy Pregnant Women
2.5. CD112 and CD155 Ligand Expression by the Four NKT Cell Subpopulations in EOP Patients and Healthy Pregnant Women
2.6. Relative TIGIT and CD226 Expression by the Four NKT Cell Subpopulations in EOP Patients and Healthy Pregnant Women
2.7. The Serum Level of CD226 and the Relationship between the Surface and the Relative Expression of CD226 by NKT Cell Subsets in EOP Patients and Healthy Pregnant Women
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Participants and Sample Collection
4.3. Lymphocyte Separation, Cryopreservation, and Thawing
4.4. Flow Cytometric Measurement
4.5. Intracellular Staining
4.6. Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, M.A.; Magee, L.A.; Kenny, L.C.; Karumanchi, S.A.; McCarthy, F.P.; Saito, S.; Hall, D.R.; Warren, C.E.; Adoyi, G.; Ishaku, S. The hypertensive disorders of pregnancy: ISSHP classification, diagnosis & management recommendations for international practice. Pregnancy Hypertens. 2018, 13, 291–310. [Google Scholar]
- Umesawa, M.; Kobashi, G. Epidemiology of hypertensive disorders in pregnancy: Prevalence, risk factors, predictors and prognosis. Hypertens. Res. 2017, 40, 213–220. [Google Scholar] [CrossRef]
- Li, X.L.; Guo, P.L.; Xue, Y.; Gou, W.L.; Tong, M.; Chen, Q. An analysis of the differences between early and late preeclampsia with severe hypertension. Pregnancy Hypertens. 2016, 6, 47–52. [Google Scholar] [CrossRef]
- Raymond, D.; Peterson, E. A critical review of early-onset and late-onset preeclampsia. Obstet. Gynecol. Surv. 2011, 66, 497–506. [Google Scholar] [CrossRef]
- Powers, R.W.; Jeyabalan, A.; Clifton, R.G.; van Dorsten, P.; Hauth, J.C.; Klebanoff, M.A.; Lindheimer, M.D.; Sibai, B.; Landon, M.; Miodovnik, M. Soluble fms-Like Tyrosine Kinase 1 (sFlt1), endoglin and placental growth factor (PlGF) in preeclampsia among high risk pregnancies. PLoS ONE 2010, 5, e13263. [Google Scholar] [CrossRef] [Green Version]
- Redman, C.W.G.; Sargent, I.L. Pre-eclampsia, the placenta and the maternal systemic inflammatory response—A review. Placenta 2003, 24 (Suppl. A), S21–S27. [Google Scholar] [CrossRef]
- Almehmadi, M.; Flanagan, B.F.; Khan, N.; Alomar, S.; Christmas, S.E. Increased numbers and functional activity of CD56+ T cells in healthy cytomegalovirus positive subjects. Immunology 2014, 142, 258–268. [Google Scholar] [CrossRef]
- Montoya, C.J.; Pollard, D.; Martinson, J.; Kumari, K.; Wasserfall, C.; Mulder, C.B.; Rugeles, M.T.; Atkinson, M.A.; Landay, A.L.; Wilson, S.B. Characterization of human invariant natural killer T subsets in health and disease using a novel invariant natural killer T cell-clonotypic monoclonal antibody, 6B11. Immunology 2007, 122, 1–14. [Google Scholar] [CrossRef]
- Godfrey, D.I.; Stankovic, S.; Baxter, A.G. Raising the NKT cell family. Nat. Immunol. 2010, 11, 197–206. [Google Scholar] [CrossRef]
- Brigl, M.; Brenner, M.B. CD1: Antigen presentation and T cell function. Annu. Rev. Immunol. 2004, 22, 817–890. [Google Scholar] [CrossRef]
- Gálvez, N.M.S.; Bohmwald, K.; Pacheco, G.A.; Andrade, C.A.; Carreño, L.J.; Kalergis, A.M. Type I natural killer T cells as key regulators of the immune response to infectious diseases. Clin. Microbiol. Rev. 2021, 34, e00232-20. [Google Scholar] [CrossRef]
- Pellicci, D.G.; Koay, H.F.; Berzins, S.P. Thymic development of unconventional T cells: How NKT cells, MAIT cells and γδ T cells emerge. Nat. Rev. Immunol. 2020, 20, 756–770. [Google Scholar] [CrossRef] [PubMed]
- Terabe, M.; Swann, J.; Ambrosino, E.; Sinha, P.; Takaku, S.; Hayakawa, Y.; Godfrey, D.I.; Ostrand-Rosenberg, S.; Smyth, M.J.; Berzofsky, J.A. A nonclassical non-Vα14Jα18 CD1d-restricted (type II) NKT cell is sufficient for down-regulation of tumor immunosurveillance. J. Exp. Med. 2005, 202, 1627–1633. [Google Scholar] [CrossRef] [Green Version]
- Renukaradhya, G.J.; Khan, M.A.; Vieira, M.; Du, W.; Gervay-Hague, J.; Brutkiewicz, R.R. Type i NKT cells protect (and type II NKT cells suppress) the host’s innate antitumor immune response to a B-cell lymphoma. Blood 2008, 111, 5637–5645. [Google Scholar] [CrossRef] [Green Version]
- Sasmal, P.; Kumar Babasahib, S.; Prashantha Kumar, B.R.; Manjunathaiah Raghavendra, N. Biphenyl-based small molecule inhibitors: Novel cancer immunotherapeutic agents targeting PD-1/PD-L1 interaction. Bioorg. Med. Chem. 2022, 73, 117001. [Google Scholar] [CrossRef]
- Ge, Y.; Zhang, Y.; Zhao, K.N.; Zhu, H. Emerging Therapeutic Strategies of Different Immunotherapy Approaches Combined with PD-1/PD-L1 Blockade in Cervical Cancer. Drug Des. Dev. Ther. 2022, 16, 3055–3070. [Google Scholar] [CrossRef]
- Mucileanu, A.; Chira, R.; Mircea, P.A. PD-1/PD-L1 expression in pancreatic cancer and its implication in novel therapies. Med. Pharm. Rep. 2021, 94, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Zhu, G.; Tamada, K.; Chen, L. B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion. Nat. Med. 1999, 5, 1365–1369. [Google Scholar] [CrossRef]
- Meggyes, M.; Miko, E.; Lajko, A.; Csiszar, B.; Sandor, B.; Matrai, P.; Tamas, P.; Szereday, L. Involvement of the PD-1/PD-L1 Co-Inhibitory Pathway in the Pathogenesis of the Inflammatory Stage of Early-Onset Preeclampsia. Int. J. Mol. Sci. 2019, 20, 583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecocq, Q.; Keyaerts, M.; Devoogdt, N.; Breckpot, K. The next-generation immune checkpoint lag-3 and its therapeutic potential in oncology: Third time’s a charm. Int. J. Mol. Sci. 2021, 22, 75. [Google Scholar] [CrossRef]
- Huang, R.Y.; Francois, A.; McGray, A.R.; Miliotto, A.; Odunsi, K. Compensatory upregulation of PD-1, LAG-3, and CTLA-4 limits the efficacy of single-agent checkpoint blockade in metastatic ovarian cancer. Oncoimmunology 2017, 6, e1249561. [Google Scholar] [CrossRef] [PubMed]
- Bruniquel, D.; Borie, N.; Hannier, S.; Triebel, F. Regulation of expression of the human lymphocyte activation gene-3 (LAG-3) molecule, a ligand for MHC class II. Immunogenetics 1998, 48, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Hemon, P.; Jean-Louis, F.; Ramgolam, K.; Brignone, C.; Viguier, M.; Bachelez, H.; Triebel, F.; Charron, D.; Aoudjit, F.; Al-Daccak, R.; et al. MHC Class II Engagement by Its Ligand LAG-3 (CD223) Contributes to Melanoma Resistance to Apoptosis. J. Immunol. 2011, 186, 5173–5183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huard, B.; Tournier, M.; Hercend, T.; Triebel, F.; Faure, F. Lymphocyte-activation gene 3/major histocompatibility complex class II interaction modulates the antigenic response of CD4+ T lymphocytes. Eur. J. Immunol. 1994, 24, 3216–3221. [Google Scholar] [CrossRef]
- Ruvolo, P.P. Galectin 3 as a guardian of the tumor microenvironment. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Stanietsky, N.; Simic, H.; Arapovic, J.; Toporik, A.; Levy, O.; Novik, A.; Levine, Z.; Beiman, M.; Dassa, L.; Achdout, H.; et al. The interaction of TIGIT with PVR and PVRL2 inhibits human NK cell cytotoxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 17858–17863. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Harden, K.; Gonzalez, L.C.; Francesco, M.; Chiang, E.; Irving, B.; Tom, I.; Ivelja, S.; Refino, C.J.; Clark, H.; et al. The surface protein TIGIT suppresses T cell activation by promoting the generation of mature immunoregulatory dendritic cells. Nat. Immunol. 2009, 10, 48–57. [Google Scholar] [CrossRef]
- Huang, Z.; Qi, G.; Miller, J.S.; Zheng, S.G. CD226: An Emerging Role in Immunologic Diseases. Front. Cell Dev. Biol. 2020, 8, 564. [Google Scholar] [CrossRef]
- Nabekura, T.; Kanaya, M.; Shibuya, A.; Fu, G.; Gascoigne, N.R.J.; Lanier, L.L. Costimulatory molecule DNAM-1 is essential for optimal differentiation of memory natural killer cells during mouse cytomegalovirus infection. Immunity 2014, 40, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Yue, C.; Gao, S.; Li, S.; Xing, Z.; Qian, H.; Hu, Y.; Wang, W.; Hua, C. TIGIT as a Promising Therapeutic Target in Autoimmune Diseases. Front. Immunol. 2022, 13, 911919. [Google Scholar] [CrossRef]
- Mendelsohn, C.L.; Wimmer, E.; Racaniello, V.R. Cellular receptor for poliovirus: Molecular cloning, nucleotide sequence, and expression of a new member of the immunoglobulin superfamily. Cell 1989, 56, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Eberlé, F.; Dubreuil, P.; Mattei, M.G.; Devilard, E.; Lopez, M. The human PRR2 gene, related to the human poliovirus receptor gene (PVR), is the true homolog of the murine MPH gene. Gene 1995, 159, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.; Aoubala, M.; Jordier, F.; Isnardon, D.; Gomez, S.; Dubreuil, P. The human poliovirus receptor related 2 protein is a new hematopoietic/endothelial homophilic adhesion molecule. Blood 1998, 92, 4602–4611. Available online: https://pubmed.ncbi.nlm.nih.gov/9845526/ (accessed on 24 October 2022). [CrossRef] [PubMed]
- Blake, S.J.; Stannard, K.; Liu, J.; Allen, S.; Yong, M.C.R.; Mittal, D.; Aguilera, A.R.; Miles, J.J.; Lutzky, V.P.; de Andrade, L.F.; et al. Suppression of metastases using a new lymphocyte checkpoint target for cancer immunotherapy. Cancer Discov. 2016, 6, 446–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, R.J.; Comps-Agrar, L.; Hackney, J.; Yu, X.; Huseni, M.; Yang, Y.; Park, S.; Javinal, V.; Chiu, H.; Irving, B.; et al. The immunoreceptor TIGIT regulates antitumor and antiviral CD8(+) T cell effector function. Cancer Cell 2014, 26, 923–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, S.; Ophir, E.; Kotturi, M.F.; Levy, O.; Ganguly, S.; Leung, L.; Vaknin, I.; Kumar, S.; Dassa, L.; Hansen, K.; et al. PVRIG and PVRL2 are induced in cancer and inhibit CD8 + T-cell function. Cancer Immunol. Res. 2019, 7, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Yusrawati; Saputra, N.P.K.; Lipoeto, N.I.; Machmud, R. Analyses of Nutrients and Body Mass Index as Risk Factor for Preeclampsia. J. Obstet. Gynecol. India 2017, 67, 409–413. [Google Scholar] [CrossRef]
- González-Comadran, M.; Avila, J.U.; Tascón, A.S.; Jimenéz, R.; Solà, I.; Brassesco, M.; Carreras, R.; Checa, M.Á. The impact of donor insemination on the risk of preeclampsia: A systematic review and meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 182, 160–166. [Google Scholar] [CrossRef]
- Saftlas, A.F.; Rubenstein, L.; Prater, K.; Harland, K.K.; Field, E.; Triche, E.W. Cumulative exposure to paternal seminal fluid prior to conception and subsequent risk of preeclampsia. J. Reprod. Immunol. 2014, 101–102, 104–110. [Google Scholar] [CrossRef]
- Valencia-Ortega, J.; Zárate, A.; Saucedo, R.; Hernández-Valencia, M.; Cruz, J.G.; Puello, E. Placental Proinflammatory State and Maternal Endothelial Dysfunction in Preeclampsia. Gynecol. Obstet. Investig. 2019, 84, 12–19. [Google Scholar] [CrossRef]
- Figueiredo, A.S.; Schumacher, A. The T helper type 17/regulatory T cell paradigm in pregnancy. Immunology 2016, 148, 13–21. [Google Scholar] [CrossRef]
- Hosseini, A.; Dolati, S.; Hashemi, V.; Abdollahpour-Alitappeh, M.; Yousefi, M. Regulatory T and T helper 17 cells: Their roles in preeclampsia. J. Cell. Physiol. 2018, 233, 6561–6573. [Google Scholar] [CrossRef]
- Robertson, S.A.; Green, E.S.; Care, A.S.; Moldenhauer, L.M.; Prins, J.R.; Louise Hull, M.; Barry, S.C.; Dekker, G. Therapeutic potential of regulatory T cells in preeclampsia-opportunities and challenges. Front. Immunol. 2019, 10, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, A.E.; Fraser, R.; Cartwright, J.E. Extravillous trophoblast and decidual natural killer cells: A remodelling partnership. Hum. Reprod. Update 2012, 18, 458–471. [Google Scholar] [CrossRef] [Green Version]
- Bendelac, A.; Savage, P.B.; Teyton, L. The biology of NKT cells. Annu. Rev. Immunol. 2007, 25, 297–336. [Google Scholar] [CrossRef] [Green Version]
- Tao, L.; Wang, S.; Kang, G.; Jiang, S.; Yin, W.; Zong, L.; Li, J.; Wang, X. PD-1 blockade improves the anti-tumor potency of exhausted CD3+CD56+ NKT-like cells in patients with primary hepatocellular carcinoma. Oncoimmunology 2021, 10, 2002068. [Google Scholar] [CrossRef]
- Hakanen, H.; Hernberg, M.; Mäkelä, S.; Yadav, B.; Brück, O.; Juteau, S.; Kohtamäki, L.; Ilander, M.; Mustjoki, S.; Kreutzman, A. Anti-PD1 therapy increases peripheral blood NKT cells and chemokines in metastatic melanoma patients. Ann. Oncol. 2018, 29, x3. [Google Scholar] [CrossRef]
- Cullen, R.; Germanov, E.; Shimaoka, T.; Johnston, B. Enhanced Tumor Metastasis in Response to Blockade of the Chemokine Receptor CXCR6 Is Overcome by NKT Cell Activation. J. Immunol. 2009, 183, 5807–5815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coquet, J.M.; Chakravarti, S.; Kyparissoudis, K.; McNab, F.W.; Pitt, L.A.; McKenzie, B.S.; Berzins, S.P.; Smyth, M.J.; Godfrey, D.I. Diverse cytokine production by NKT cell subsets and identification of an IL-17-producing CD4-NK1.1-NKT cell population. Proc. Natl. Acad. Sci. USA 2008, 105, 11287–11292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaki, J.; Gnjatic, S.; Mhawech-Fauceglia, P.; Beck, A.; Miller, A.; Tsuji, T.; Eppolito, C.; Qian, F.; Lele, S.; Shrikant, P.; et al. Tumor-infiltrating NY-ESO-1-specific CD8+ T cells are negatively regulated by LAG-3 and PD-1 in human ovarian cancer. Proc. Natl. Acad. Sci. USA 2010, 107, 7875–7880. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, N.; Sugaya, M.; Suga, H.; Oka, T.; Kawaguchi, M.; Miyagaki, T.; Fujita, H.; Inozume, T.; Sato, S. Increased Soluble CD226 in Sera of Patients with Cutaneous T-Cell Lymphoma Mediates Cytotoxic Activity against Tumor Cells via CD155. J. Invest. Dermatol. 2017, 137, 1766–1773. [Google Scholar] [CrossRef] [PubMed]
- Szereday, L.; Nagy, D.U.; Csiszar, B.; Kevey, D.; Feik, T.; Meggyes, M. Examination of the tigit, cd226, cd112, and cd155 immune checkpoint molecules in peripheral blood mononuclear cells in women diagnosed with early-onset preeclampsia. Biomedicines 2021, 9, 1608. [Google Scholar] [CrossRef] [PubMed]
- Meggyes, M.; Nagy, D.U.; Feik, T.; Boros, A.; Polgar, B.; Szereday, L. Examination of the TIGIT-CD226-CD112-CD155 Immune Checkpoint Network during a Healthy Pregnancy. Int. J. Mol. Sci. 2022, 23, 10776. [Google Scholar] [CrossRef] [PubMed]
- Crawley, M.J. Statistics: An Introduction Using R; Wiley: New York, NY, USA, 2014; ISBN 9781118941096. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Pregnant Women | Early-Onset Preeclamptic Patients | |
|---|---|---|
| No. of patients | 51 | 32 |
| Age (years) | 32.84 (23–44) | 29.81 (18–43) |
| Gestational age at birth (weeks) | 39.06 ± 0.97 | 31.31 ± 3.17 * |
| Gestational age at sampling (weeks) | 33.11 ± 3.91 | 30.36 ± 2.53 |
| Birth weight (gram) | 3447.38 ± 411.04 | 1429.14 ± 599.97 * |
| Antigen | Format | Clone | Isotype | Company | CAT |
|---|---|---|---|---|---|
| CD112 | PE | R2.525 | Mouse IgG1, κ | BD Biosciences | 551057 |
| CD155 | APC | SKII.4 | Mouse IgG1, κ | Biolegend | 337618 |
| CD3 | BV510 | UCHT1 | Mouse BALB/c IgG1, κ | BD Biosciences | 563109 |
| CD4 | FITC | RPA-T4 | Mouse IgG1, κ | BD Biosciences | 555346 |
| CD8 | APC-H7 | SK1 | Mouse BALB/c IgG1, κ | BD Biosciences | 560179 |
| CD56 | APC | B159 | Mouse IgG1, κ | BD Biosciences | 555518 |
| CD226 | BV421 | DX11 | Mouse BALB/c IgG1, κ | BD Biosciences | 742493 |
| Galectin-3 | PE | B2C10 | Mouse BALB/c IgG1, κ | BD Biosciences | 565676 |
| LAG-3 | PerCp Cy5.5 | 11C3C65 | Mouse IgG1, κ | Biolegend | 369312 |
| TIGIT | PE | A1553G | Mouse IgG2a, κ | Biolegend | 372704 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meggyes, M.; Feik, T.; Nagy, D.U.; Polgar, B.; Szereday, L. CD8 and CD4 Positive NKT Subpopulations and Immune-Checkpoint Pathways in Early-Onset Preeclampsia and Healthy Pregnancy. Int. J. Mol. Sci. 2023, 24, 1390. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021390
Meggyes M, Feik T, Nagy DU, Polgar B, Szereday L. CD8 and CD4 Positive NKT Subpopulations and Immune-Checkpoint Pathways in Early-Onset Preeclampsia and Healthy Pregnancy. International Journal of Molecular Sciences. 2023; 24(2):1390. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021390
Chicago/Turabian StyleMeggyes, Matyas, Timoteus Feik, David U. Nagy, Beata Polgar, and Laszlo Szereday. 2023. "CD8 and CD4 Positive NKT Subpopulations and Immune-Checkpoint Pathways in Early-Onset Preeclampsia and Healthy Pregnancy" International Journal of Molecular Sciences 24, no. 2: 1390. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24021390