
A Low-Fat/Sucrose Diet Rich in Complex Carbohydrates Reverses High-Fat/Sucrose Diet-Induced Corneal Dysregulation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. High-Fat/Sucrose Diet Feeding for 10 Weeks Causes Significant Adiposity and Impaired Insulin Metabolism
2.2. High-Fat/Sucrose Feeding for 10 Weeks Impairs Corneal Wound Healing and Dysregulates Abrasion-Induced Inflammation
2.3. Diet Reversal Attenuates High-Fat/Sucrose Diet-Induced Adiposity and Liver Weight
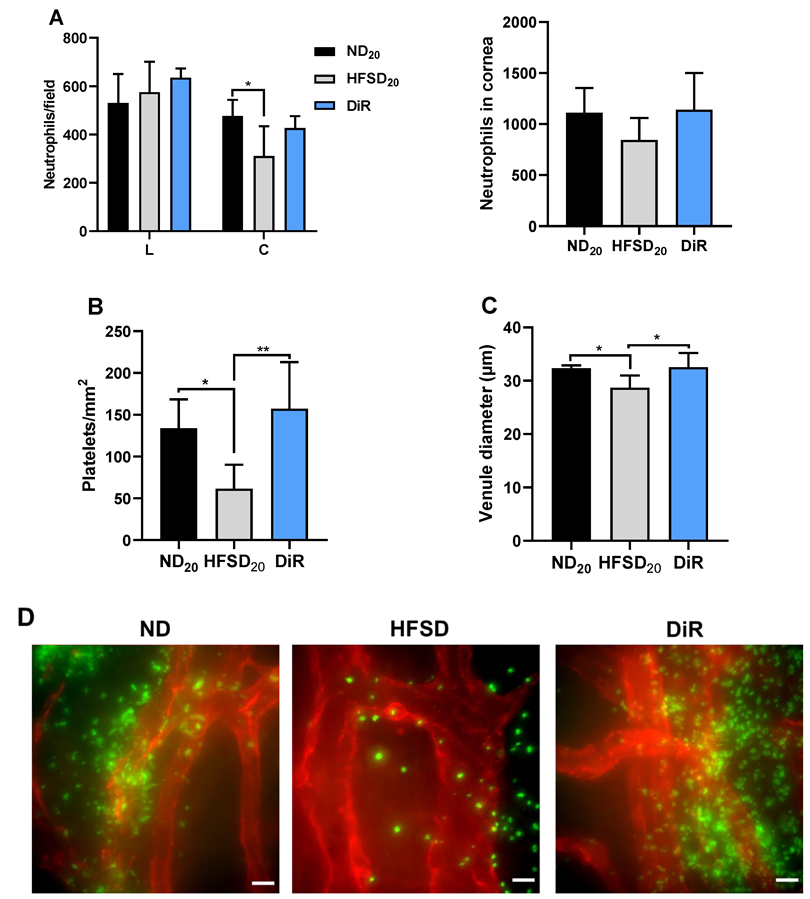
2.4. Diet Reversal Halts Progression of Corneal Nerve Function Loss and Restored Normal Corneal Wound Healing
3. Discussion
4. Materials and Methods
4.1. Mice, Diet, and Experimental Design
4.2. Body Adiposity and Fatty Liver
4.3. Fasting Blood Glucose and Plasma Insulin
4.4. Corneal Nerve Function
4.5. Corneal Wound Healing
4.6. Immunofluorescence Staining
4.7. Morphometric Analysis of Neutrophil and Platelet Recruitment and Blood Vessel Diameter
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Swinburn, B.; Sacks, G.; Hall, K.; McPherson, K.; Finegood, D.; Moodie, M.; Gortmaker, S.L. The Global Obesity Pandemic: Shaped By Global Drivers and Local Environments. Lancet 2011, 378, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Romieu, I.; Dossus, L.; Barquera, S.; Blottière, H.M.; Franks, P.W.; Gunter, M.; Hwalla, N.; Hursting, S.D.; Leitzmann, M.; Margetts, B.; et al. Energy Balance and Obesity: What Are the Main Drivers? Cancer Causes Control. 2017, 28, 247–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, J.O.; Wyatt, H.R.; Peters, J.C. Energy Balance and Obesity. Circulation 2012, 126, 126–132. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Obesity and Overweight: Key Facts. WHO 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 11 February 2021).
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity and Severe Obesity among Adults: United States, 2017–2018; NCHSNCHS Data Brief, no 360; National Center for Health Statistics: Hyattsville, MD, USA, 2020. [Google Scholar]
- Pi-Sunyer, X. The medical risks of obesity. Postgrad. Med. 2009, 121, 21–33. [Google Scholar] [CrossRef]
- Ginsberg, H.N.; Maccallum, P.R. The Obesity, Metabolic Syndrome, and Type 2 Diabetes Mellitus Pandemic: Part I. Increased Cardiovascular Disease Risk and The Importance of Atherogenic Dyslipidemia in Persons with the Metabolic Syndrome and Type 2 Diabetes Mellitus. J. Cardiometabolic Syndr. 2009, 4, 113–119. [Google Scholar] [CrossRef]
- Han, T.S.; Lean, M.E. A Clinical Perspective of Obesity, Metabolic Syndrome and Cardiovascular Disease. JRSM Cardiovasc. Dis. 2016, 5, 204800401663337. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-Y.; McCulloch, C.E.; Iribarren, C.; Darbinian, J.; Go, A.S. Body Mass Index and Risk for End-Stage Renal Disease. Ann. Intern. Med. 2006, 144, 21–28. [Google Scholar] [CrossRef]
- Leitner, D.R.; Frühbeck, G.; Yumuk, V.; Schindler, K.; Micic, D.; Woodward, E.; Toplak, H. Obesity and Type 2 Diabetes: Two Diseases with a Need for Combined Treatment Strategies—EASO Can Lead the Way. Obes. Facts 2017, 10, 483–492. [Google Scholar] [CrossRef]
- Wu, Y.; Ding, Y.; Tanaka, Y.; Zhang, W. Risk Factors Contributing to Type 2 Diabetes and Recent Advances in the Treatment and Prevention. Int. J. Med. Sci. 2014, 11, 1185–1200. [Google Scholar] [CrossRef] [Green Version]
- Barnes, A.S. The epidemic of obesity and diabetes: Trends and treatments. Tex. Heart Inst. J. 2011, 38, 142–144. [Google Scholar]
- Priyadarsini, S.; Whelchel, A.; Nicholas, S.; Sharif, R.; Riaz, K.; Karamichos, D. Diabetic Keratopathy: Insights and Challenges. Surv. Ophthalmol. 2020, 65, 513–529. [Google Scholar] [CrossRef] [PubMed]
- Ljubimov, A.V. Diabetic Complications in the Cornea. Vis. Res. 2017, 139, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.P.; Coppey, L.J.; Kardon, R.; Yorek, M.A. Differences and Similarities in Development of Corneal Nerve Damage and Peripheral Neuropathy and in Diet- Induced Obesity and Type 2 Diabetic Rats. Investig. Ophthalmol. Vis. Sci. 2014, 55, 1222–1230. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Huang, J.; Chen, C.; Gao, N.; Wang, F.; Yu, F.-S.X. Corneal Complications in Streptozocin-Induced Type I Diabetic Rats. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6589–6596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matlock, H.G.; Qiu, F.; Malechka, V.; Zhou, K.; Cheng, R.; Benyajati, S.; Whelchel, A.; Karamichos, D.; Ma, J.-X. Pathogenic Role of PPARα Downregulation in Corneal Nerve Degeneration and Impaired Corneal Sensitivity in Diabetes. Diabetes 2020, 69, 1279–1291. [Google Scholar] [CrossRef]
- Hargrave, A.; Courson, J.A.; Pham, V.; Landry, P.; Magadi, S.; Shankar, P.; Hanlon, S.; Das, A.; Rumbaut, R.E.; Smith, C.W.; et al. Corneal Dysfunction Precedes the Onset of Hyperglycemia in a Mouse Model of Diet-Induced Obesity. PLoS ONE 2020, 15, e0238750. [Google Scholar] [CrossRef]
- Akowuah, P.K.; Hargrave, A.; Rumbaut, R.E.; Burns, A.R. Dissociation between Corneal and Cardiometabolic Changes in Response to a Time-Restricted Feeding of a High Fat Diet. Nutrients 2021, 14, 139. [Google Scholar] [CrossRef]
- Yorek, M.S.; Obrosov, A.; Shevalye, H.; Holmes, A.; Harper, M.M.; Kardon, R.H.; Yorek, M.A. Effect of Diet-Induced Obesity or Type 1 or Type 2 Diabetes on Corneal Nerves and Peripheral Neuropathy in C57Bl/6J Mice. J. Peripher. Nerv. Syst. 2015, 20, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, Z.; Kalteniece, A.; Ferdousi, M.; Adam, S.; D’Onofrio, L.; Ho, J.H.; Rao, A.P.; Dhage, S.; Azmi, S.; Liu, Y.; et al. Corneal Keratocyte Density and Corneal Nerves Are Reduced in Patients With Severe Obesity and Improve After Bariatric Surgery. Investig. Ophthalmol. Vis. Sci. 2021, 62, 20. [Google Scholar] [CrossRef]
- Azmi, S.; Ferdousi, M.; Liu, Y.; Adam, S.; Iqbal, Z.; Dhage, S.; Ponirakis, G.; Siahmansur, T.; Marshall, A.; Petropoulos, I.; et al. Bariatric Surgery Leads to an Improvement in Small Nerve Fibre Damage in Subjects With Obesity. Int. J. Obes. 2021, 45, 631–638. [Google Scholar] [CrossRef]
- O’Donnell, C.; Efron, N. Diabetes and Contact Lens Wear. Clin. Exp. Optom. 2012, 95, 328–337. [Google Scholar] [CrossRef]
- Vieira-Potter, V.J.; Karamichos, D.; Lee, D. Ocular Complications of Diabetes and Therapeutic Approaches. BioMed Res. Int. 2016, 2016, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Schulze, S.D.; Sekundo, W.; Kroll, P. Autologous Serum for the Treatment of Corneal Epithelial Abrasions in Diabetic Patients Undergoing Vitrectomy. Am. J. Ophthalmol. 2006, 142, 207–211. [Google Scholar] [CrossRef]
- McTigue, K.M.; Harris, R.; Hemphill, B.; Lux, L.; Sutton, S.; Bunton, A.J.; Lohr, K.N. Screening and Interventions for Obesity in Adults: Summary of the Evidence for the U.S. Preventive Services Task Force. Ann. Intern. Med. 2003, 139, 933–949. [Google Scholar] [CrossRef] [PubMed]
- Sims-Robinson, C.; Bakeman, A.; Bruno, E.; Jackson, S.; Glasser, R.; Murphy, G.G.; Feldman, E.L. Dietary Reversal Ameliorates Short- and Long-Term Memory Deficits Induced by High-fat Diet Early in Life. PLoS ONE 2016, 11, e0163883. [Google Scholar] [CrossRef] [Green Version]
- Kalita, H.; Hazarika, A.; Devi, R. Withdrawal of High-Carbohydrate, High-Fat Diet Alters Status of Trace Elements to Ameliorate Metabolic Syndrome in Rats With Type 2 Diabetes Mellitus. Can. J. Diabetes 2020, 44, 317–326.e1. [Google Scholar] [CrossRef]
- Coppey, L.; Davidson, E.; Shevalye, H.; Obrosov, A.; Torres, M.; Yorek, M.A. Progressive Loss of Corneal Nerve Fibers and Sensitivity in Rats Modeling Obesity and Type 2 Diabetes is Reversible with Omega-3 Fatty Acid Intervention: Supporting Cornea Analyses as a Marker for Peripheral Neuropathy and Treatment. Diabetes Metab. Syndr. Obes. 2020, 13, 1367–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branecky, K.L.; Niswender, K.D.; Pendergast, J.S. Disruption of Daily Rhythms by High-Fat Diet Is Reversible. PLoS ONE 2015, 10, e0137970. [Google Scholar] [CrossRef] [PubMed]
- Bray, J.K.; Chiu, G.S.; McNeil, L.K.; Moon, M.L.; Wall, R.; Towers, A.; Freund, G.G. Switching From a High-Fat Cellulose Diet to a High-Fat Pectin Diet Reverses Certain Obesity-Related Morbidities. Nutr. Metab. 2018, 15, 55. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Rumbaut, R.E.; Burns, A.R.; Smith, C.W. Platelet Response to Corneal Abrasion is Necessary for Acute Inflammation and Efficient Re-Epithelialization. Investig. Ophthalmol. Vis. Sci. 2006, 47, 4794–4802. [Google Scholar] [CrossRef]
- Li, Z.; Burns, A.R.; Smith, C.W. Two Waves of Neutrophil Emigration in Response to Corneal Epithelial Abrasion: Distinct Adhesion Molecule Requirements. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1947–1955. [Google Scholar] [CrossRef]
- Della Vedova, M.C.; Muñoz, M.D.; Santillan, L.D.; Plateo-Pignatari, M.G.; Germanó, M.J.; Rinaldi Tosi, M.E.; Garcia, S.; Gomez, N.N.; Fornes, M.W.; Gomez Mejiba, S.E.; et al. A Mouse Model of Diet-Induced Obesity Resembling Most Features of Human Metabolic Syndrome. Nutr. Metab. Insights 2016, 9, 93–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Perrard, X.D.; Wang, Q.; Perrard, J.L.; Polsani, V.R.; Jones, P.H.; Smith, C.W.; Ballantyne, C.M. CD11c Expression in Adipose Tissue and Blood and its Role in Diet-Induced Obesity. Arter. Thromb. Vasc. Biol. 2010, 30, 186–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osae, E.A.; Bullock, T.; Chintapalati, M.; Brodesser, S.; Hanlon, S.; Redfern, R.; Steven, P.; Smith, C.W.; Rumbaut, R.E.; Burns, A.R. Obese Mice with Dyslipidemia Exhibit Meibomian Gland Hypertrophy and Alterations in Meibum Composition and Aqueous Tear Production. Int. J. Mol. Sci. 2020, 21, 8772. [Google Scholar] [CrossRef]
- Roberts, C.K.; Vaziri, N.D.; Liang, K.H.; Barnard, R.J. Reversibility of chronic experimental syndrome X by diet modification. Hypertension 2001, 37, 1323–1328. [Google Scholar] [CrossRef] [Green Version]
- Parekh, P.I.; Petro, A.E.; Tiller, J.M.; Feinglos, M.N.; Surwit, R.S. Reversal of Diet-Induced Obesity and Diabetes in C57BL/6J Mice. Metabolism 1998, 47, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Pendergast, J.S.; Branecky, K.L.; Yang, W.; Ellacott, K.L.J.; Niswender, K.D.; Yamazaki, S. High-Fat Diet Acutely Affects Circadian Organisation and Eating Behavior. Eur. J. Neurosci. 2013, 37, 1350–1356. [Google Scholar] [CrossRef] [Green Version]
- Kohsaka, A.; Laposky, A.D.; Ramsey, K.M.; Estrada, C.; Joshu, C.; Kobayashi, Y.; Turek, F.W.; Bass, J. High-Fat Diet Disrupts Behavioral and Molecular Circadian Rhythms in Mice. Cell Metab. 2007, 6, 414–421. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.; Zhou, Q.; McGinn, L.; Nguyen, T.; Luo, Y.; Djalilian, A.; Rosenblatt, M. High Fat Diet Induced Gut Dysbiosis Alters Corneal Epithelial Injury Response in Mice. Ocul. Surf. 2022, 23, 49–59. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Carmody, R.N.; Gerber, G.K.; Luevano, J.M., Jr.; Gatti, D.M.; Somes, L.; Svenson, K.L.; Turnbaugh, P.J. Diet Dominates Host Genotype in Shaping the Murine Gut Microbiota. Cell Host Microbe 2015, 17, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Carmody, R.N.; Gerber, G.K.; Luevano, J.M., Jr.; Gatti, D.M.; Somes, L.; Svenson, K.L.; Turnbaugh, P.J. Environment Dominates Over Host Genetics in Shaping Human Gut Microbiota. Nature 2018, 555, 210–215. [Google Scholar]
- David, L.A.; Materna, A.C.; Friedman, J.; Campos-Baptista, M.I.; Blackburn, M.C.; Perrotta, A.; Erdman, S.E.; Alm, E.J. Host Lifestyle Affects Human Microbiota on Daily Timescales. Genome Biol. 2014, 15, R89. [Google Scholar] [CrossRef] [Green Version]
- De Wit, N.; Derrien, M.; Bosch-Vermeulen, H.; Oosterink, E.; Keshtkar, S.; Duval, C.; de Vogel-van den Bosch, J.; Kleerebezem, M.; Müller, M.; van der Meer, R. Saturated Fat Stimulates Obesity and Hepatic Steatosis and Affects Gut Microbiota Composition by an Enhanced Overflow of Dietary Fat to the Distal Intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G589–G599. [Google Scholar] [CrossRef] [Green Version]
- Hildebrandt, M.A.; Hoffmann, C.; Sherrill–Mix, S.A.; Keilbaugh, S.A.; Hamady, M.; Chen, Y.-Y.; Knight, R.; Ahima, R.S.; Bushman, F.; Wu, G.D. High-Fat Diet Determines the Composition of the Murine Gut Microbiome Independently of Obesity. Gastroenterology 2009, 137, 1716–1724.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosas-Villegas, A.; Sánchez-Tapia, M.; Avila-Nava, A.; Ramírez, V.; Tovar, A.R.; Torres, N. Differential Effect of Sucrose and Fructose in Combination with a High Fat Diet on Intestinal Microbiota and Kidney Oxidative Stress. Nutrients 2017, 9, 393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity Alters Gut Microbial Ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Bäckhed, F.; Fulton, L.; Gordon, J.I. Diet-Induced Obesity Is Linked to Marked but Reversible Alterations in the Mouse Distal Gut Microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Roszkowska, A.M.; Colosi, P.; Ferreri, F.M.; Galasso, S. Age-Related Modifications of Corneal Sensitivity. Ophthalmologica 2004, 218, 350–355. [Google Scholar] [CrossRef]
- Mirzajan, A.; Khezri, F.; Jafarzadehpur, E.; Karimian, F.; Khabazkhoob, M. Normal Corneal Sensitivity and Its Changes With Age in Tehran, Iran. Clin. Exp. Optom. 2015, 98, 54–57. [Google Scholar] [CrossRef] [PubMed]
- De Silva, M.E.H.; Hill, L.J.; Downie, L.E.; Chinnery, H.R. The Effects of Aging on Corneal and Ocular Surface Homeostasis in Mice. Investig. Ophthalmol. Vis. Sci. 2019, 60, 2705–2715. [Google Scholar] [CrossRef] [PubMed]
- Coppey, L.; Davidson, E.; Shevalye, H.; Torres, M.E.; Yorek, M.A. Effect of Dietary Oils on Peripheral Neuropathy-Related Endpoints in Dietary Obese Rats. Diabetes Metab. Syndr. Obes. 2018, 11, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Rumora, A.E.; Lograsso, G.; Hayes, J.M.; Mendelson, F.E.; Tabbey, M.A.; Haidar, J.A.; Lentz, S.I.; Feldman, E.L. The Divergent Roles of Dietary Saturated and Monounsaturated Fatty Acids on Nerve Function in Murine Models of Obesity. J. Neurosci. 2019, 39, 3770–3781. [Google Scholar] [CrossRef] [Green Version]
- Hinder, L.M.; O’Brien, P.D.; Hayes, J.M.; Backus, C.; Solway, A.P.; Sims-Robinson, C.; Feldman, E.L. Dietary Reversal of Neuropathy in a Murine Model of Prediabetes and Metabolic Syndrome. Dis. Model. Mech. 2017, 10, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, T.L. Mitochondrial Trafficking in Neurons. Cold Spring Harb. Perspect. Biol. 2013, 5, a011304. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Z.H. Mitochondrial Trafficking and Anchoring in Neurons: New Insight and Implications. J. Cell Biol. 2014, 204, 1087–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumora, A.E.; Lentz, S.I.; Hinder, L.M.; Jackson, S.W.; Valesano, A.; Levinson, G.E.; Feldman, E.L. Dyslipidemia Impairs Mitochondrial Trafficking and Function in Sensory Neurons. FASEB J. 2018, 32, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Rumora, A.E.; LoGrasso, G.; Haidar, J.A.; Dolkowski, J.J.; Lentz, S.I.; Feldman, E.L. Chain Length of Saturated Fatty Acids Regulates Mitochondrial Trafficking and Function in Sensory Neurons. J. Lipid Res. 2019, 60, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Burhans, M.S.; Flowers, M.T.; Harrington, K.; Bond, L.M.; Guo, C.-A.; Anderson, R.M.; Ntambi, J.M. Hepatic Oleate Regulates Adipose Tissue Lipogenesis and Fatty Acid Oxidation. J. Lipid Res. 2015, 56, 304–318. [Google Scholar] [CrossRef] [Green Version]
- Henique, C.; Mansouri, A.; Fumey, G.; Lenoir, V.; Girard, J.; Bouillaud, F.; Prip-Buus, C.; Cohen, I. Increased Mitochondrial Fatty Acid Oxidation Is Sufficient To Protect Skeletal Muscle Cells From Palmitate-Induced Apoptosis. J. Biol. Chem. 2010, 285, 36818–36827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, S.K.; Mondal, A.K.; Elbein, S.C. Distinct Gene Expression Profiles Characterize Cellular Responses to Palmitate and Oleate. J. Lipid Res. 2010, 51, 2121–2131. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.J.H.; Perkins, B.A.; Lovblom, L.E.; Bazinet, R.P.; Wolever, T.M.S.; Bril, V. Effect of Omega-3 Supplementation on Neuropathy in Type 1 Diabetes: A 12-Month Pilot Trial. Neurology 2017, 88, 2294–2301. [Google Scholar] [CrossRef] [PubMed]
- Nickelson, K.J.; Stromsdorfer, K.L.; Pickering, R.; Liu, T.-W.; Ortinau, L.C.; Keating, A.F.; Perfield, J.W. A Comparison of Inflammatory and Oxidative Stress Markers in Adipose Tissue from Weight-Matched Obese Male and Female Mice. Exp. Diabetes Res. 2012, 2012, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Stubbins, R.E.; Najjar, K.; Holcomb, V.B.; Hong, J.; Núñez, N.P. Oestrogen Alters Adipocyte Biology and Protects Female Mice from Adipocyte Inflammation and Insulin Resistance. Diabetes Obes. Metab. 2012, 14, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stubbins, R.E.; Holcomb, V.B.; Hong, J.; Núñez, N.P. Estrogen Modulates Abdominal Adiposity and Protects Female Mice from Obesity and Impaired Glucose Tolerance. Eur. J. Nutr. 2012, 51, 861–870. [Google Scholar] [CrossRef]
- Chaix, A.; Zarrinpar, A.; Miu, P.; Panda, S. Time-Restricted Feeding Is a Preventative and Therapeutic Intervention Against Diverse Nutritional Challenges. Cell Metab. 2014, 20, 991–1005. [Google Scholar] [CrossRef] [Green Version]
- Avtanski, D.; Pavlov, V.A.; Tracey, K.J.; Poretsky, L. Characterization of Inflammation and Insulin Resistance in High-Fat Diet-Induced Male C57BL/6J Mouse Model of Obesity. Anim. Model. Exp. Med. 2019, 2, 252–258. [Google Scholar] [CrossRef] [Green Version]
- Hargrave, S.L.; Davidson, T.L.; Zheng, W.; Kinzig, K.P. Western Diets Induce Blood-Brain Barrier Leakage and Alter Spatial Strategies in Rats. Behav. Neurosci. 2016, 130, 123–135. [Google Scholar] [CrossRef]
- Bortolin, R.C.; Vargas, A.R.; Gasparotto, J.; Chaves, P.R.; Schnorr, C.E.; Martinello, K.B.; Silveira, A.K.; Rabelo, T.K.; Gelain, D.P.; Moreira, J.C.F. A new animal diet based on human Western diet is a robust diet-induced obesity model: Comparison to high-fat and cafeteria diets in term of metabolic and gut microbiota disruption. Int. J. Obes. 2018, 42, 525–534. [Google Scholar] [CrossRef]
- Woodie, L.N.; Johnson, R.M.; Ahmed, B.; Fowler, S.; Haynes, W.; Carmona, B.; Reed, M.; Suppiramaniam, V.; Greene, M.W. Western diet-induced obesity disrupts the diurnal rhythmicity of hippocampal core clock gene expression in a mouse model. Brain Behav. Immun. 2020, 88, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.; et al. Time-Restricted Feeding Without Reducing Caloric Intake Prevents Metabolic Diseases in Mice Fed a High-Fat Diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [PubMed]
- Chaix, A.; Lin, T.; Le, H.D.; Chang, M.W.; Panda, S. Time-Restricted Feeding Prevents Obesity and Metabolic Syndrome in Mice Lacking a Circadian Clock. Cell Metab. 2019, 29, 303–319.e4. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.; Elkfury, J.L.; Jornada, M.N.; Foletto, K.C.; Bertoluci, M.C. Validation of HOMA-IR in a Model of Insulin-Resistance Induced by a High-Fat Diet in Wistar Rats. Arch. Endocrinol. Metab. 2016, 60, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Fraulob, J.C.; Ogg-Diamantino, R.; Fernandes-Santos, C.; Aguila, M.B.; Mandarim-De-Lacerda, C.A. A Mouse Model of Metabolic Syndrome: Insulin Resistance, Fatty Liver and Non-Alcoholic Fatty Pancreas Disease (NAFPD) in C57BL/6 Mice Fed a High Fat Diet. J. Clin. Biochem. Nutr. 2010, 46, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Akowuah, P.K.; De La Cruz, A.; Smith, C.W.; Rumbaut, R.E.; Burns, A.R. An Epithelial Abrasion Model for Studying Corneal Wound Healing. J. Vis. Exp. 2021, 178, e63112. [Google Scholar] [CrossRef]
- Li, Z.; Rivera, C.A.; Burns, A.R.; Smith, C.W. Hind limb Unloading Depresses Corneal Epithelial Wound Healing in Mice. J. Appl. Physiol. 2004, 97, 641–647. [Google Scholar] [CrossRef] [Green Version]
- la Cruz, A.D.; Hargrave, A.; Magadi, S.; Courson, J.A.; Landry, P.T.; Zhang, W.; Lam, F.W.; Bray, M.A.; Smith, C.W.; Burns, A.R.; et al. Platelet and Erythrocyte Extravasation Across Inflamed Corneal Venules Depend on CD18, Neutrophils, and Mast Cell Degranulation. Int. J. Mol. Sci. 2021, 22, 7360. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akowuah, P.K.; Lema, C.; Rumbaut, R.E.; Burns, A.R. A Low-Fat/Sucrose Diet Rich in Complex Carbohydrates Reverses High-Fat/Sucrose Diet-Induced Corneal Dysregulation. Int. J. Mol. Sci. 2023, 24, 931. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24020931
Akowuah PK, Lema C, Rumbaut RE, Burns AR. A Low-Fat/Sucrose Diet Rich in Complex Carbohydrates Reverses High-Fat/Sucrose Diet-Induced Corneal Dysregulation. International Journal of Molecular Sciences. 2023; 24(2):931. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24020931
Chicago/Turabian StyleAkowuah, Prince K., Carolina Lema, Rolando E. Rumbaut, and Alan R. Burns. 2023. "A Low-Fat/Sucrose Diet Rich in Complex Carbohydrates Reverses High-Fat/Sucrose Diet-Induced Corneal Dysregulation" International Journal of Molecular Sciences 24, no. 2: 931. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24020931