
Cannabidiol Modulates Emotional Function and Brain-Derived Neurotrophic Factor Expression in Middle-Aged Female Rats Exposed to Social Isolation

Abstract
:1. Introduction
2. Results
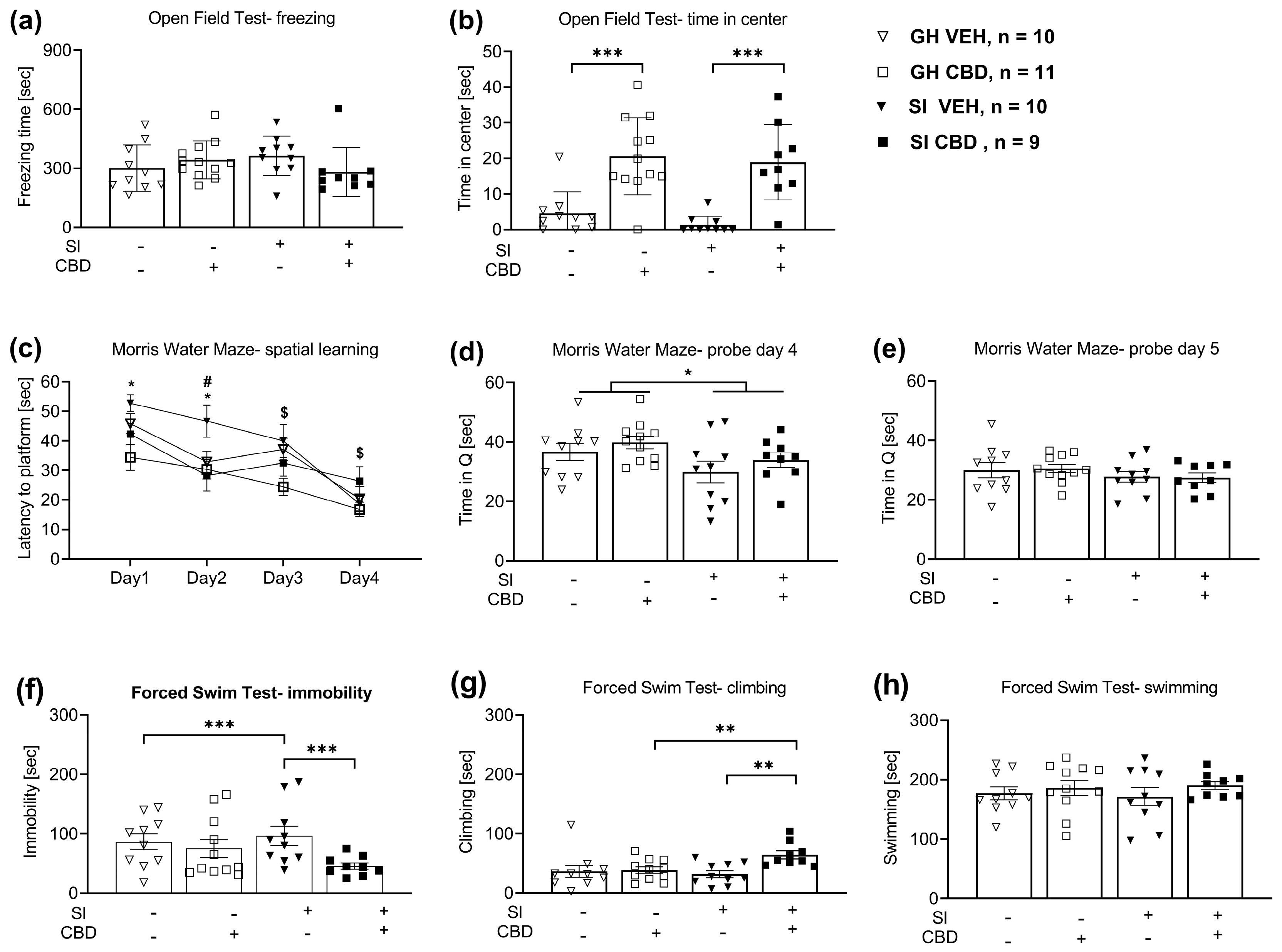
2.1. The Effects of CBD and SI on Behavior
2.1.1. Open Field Test (OFT)
2.1.2. Morris Water Maze (MWM)
2.1.3. Forced Swim Test (FST)
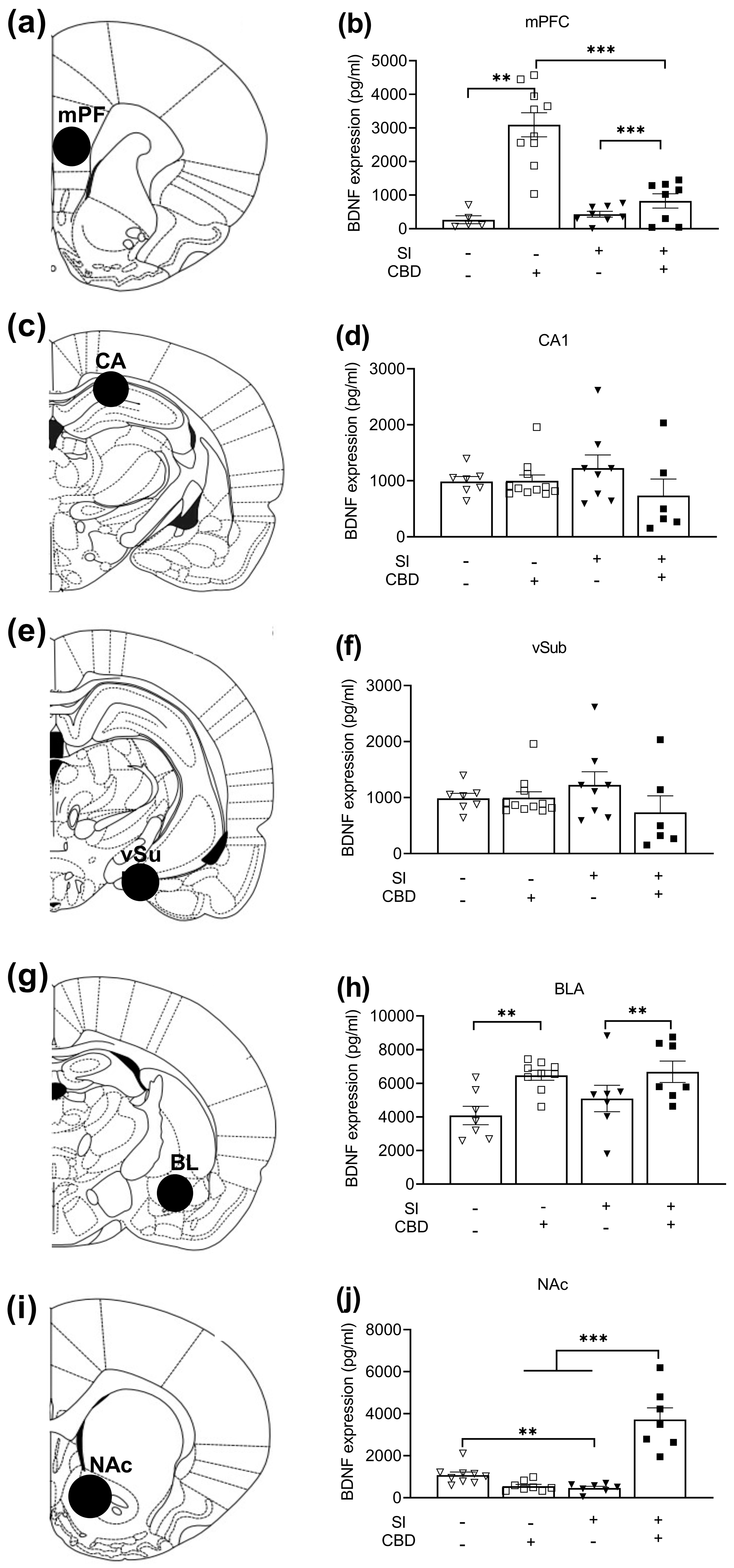
2.2. The Effects of CBD and SI on BDNF Protein Expression
2.2.1. Medial PFC (mPFC)
2.2.2. CA1
2.2.3. Ventral Subiculum (VSUB)
2.2.4. Basolateral Amygdala (BLA)
2.2.5. Nucleus Accumbens (NAc)
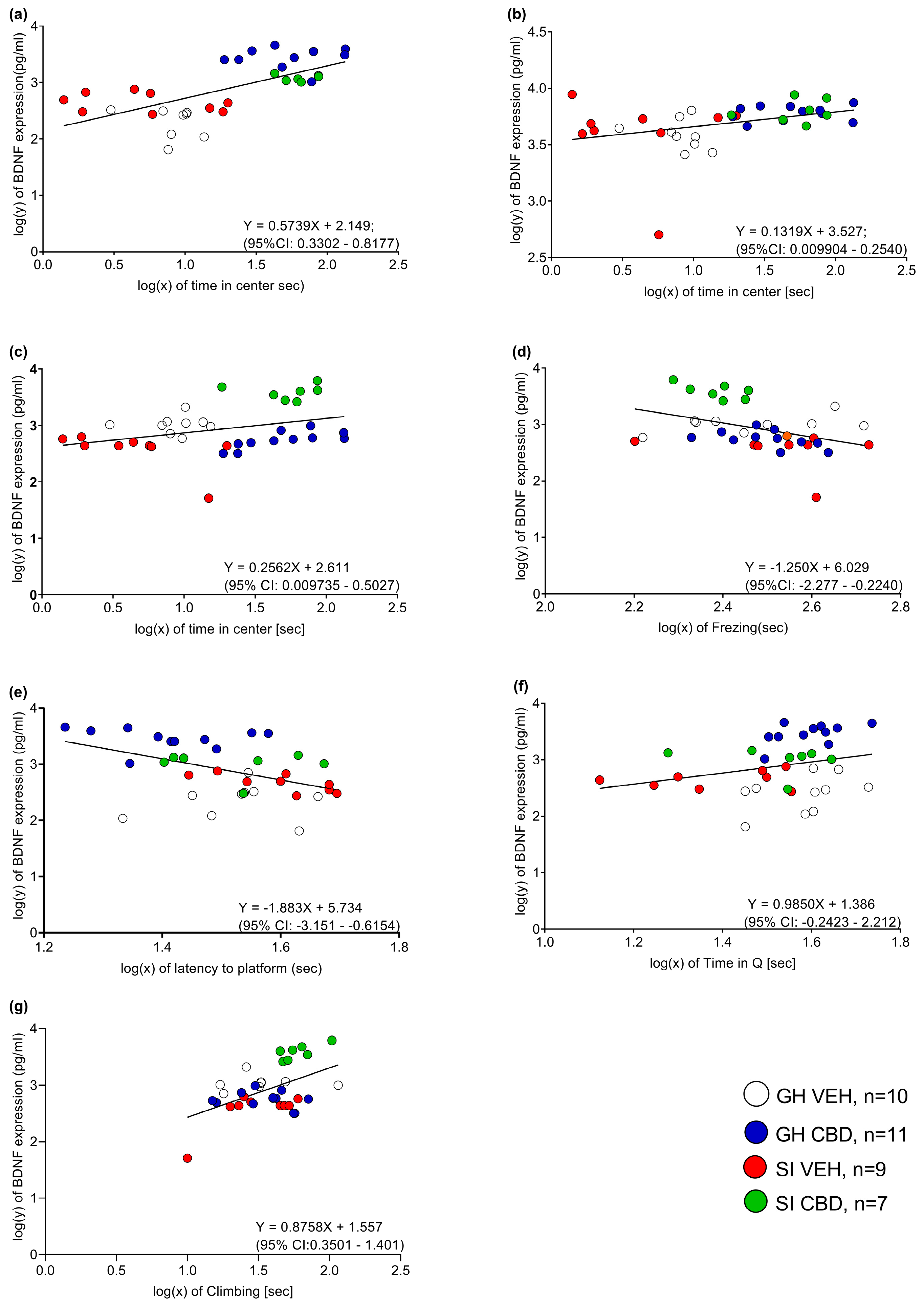
2.3. Correlations between BDNF Levels and Behavior
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Drug Treatment
4.3. Social Isolation Stress Paradigm (SI)
4.4. Behavioral Tests
4.4.1. Open Field (OFT)
4.4.2. Morris Water Maze Test (MWM)
4.4.3. Forced Swim Test (FST)
4.5. BDNF Protein Measurement via Enzyme-Linked Immunosorbent Assay (ELISA)
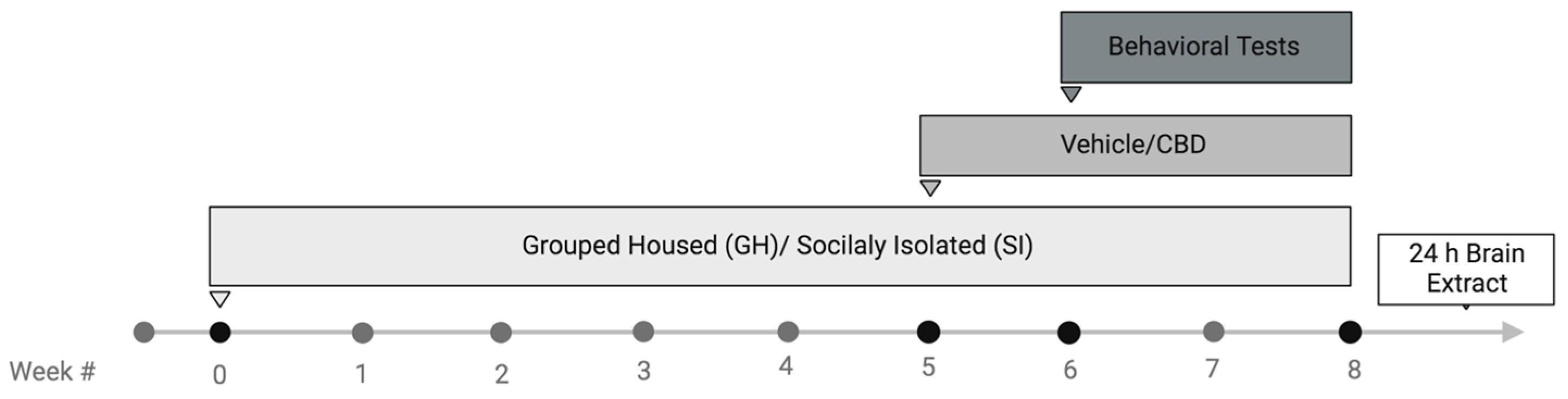
4.6. Experimental Design
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bektas, A.; Schurman, S.H.; Sen, R.; Ferrucci, L. Aging, Inflammation and the Environment. Exp. Gerontol. 2018, 105, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, K.R.; Scruggs, B.; Aniya, J.; Wright, K.C.; Ontl, T.; Xing, Y.; Bai, L. Age-related deficits in mice performing working memory tasks in a water maze. Behav. Neurosci. 2003, 117, 485. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Hernández, E.; García-Fuster, M.J. Dose-dependent antidepressant-like effects of cannabidiol in aged rats. Front. Pharmacol. 2022, 13, 891842. [Google Scholar] [CrossRef]
- McKinney, B.C.; Sibille, E. The age-by-disease interaction hypothesis of late-life depression. Am. J. Geriatr. Psychiatry 2013, 21, 418–432. [Google Scholar] [CrossRef] [PubMed]
- Sabolek, H.R.; Bunce, J.G.; Giuliana, D.; Chrobak, J.J. Within-subject memory decline in middle-aged rats: Effects of intraseptal tacrine. Neurobiol. Aging 2004, 25, 1221–1229. [Google Scholar] [CrossRef]
- Fischer, W.; Chen, K.S.; Gage, F.H.; Björklund, A. Progressive decline in spatial learning and integrity of forebrain cholinergic neurons in rats during aging. Neurobiol. Aging 1992, 13, 9–23. [Google Scholar] [CrossRef]
- Muir, J.L.; Fischer, W.; Björklund, A. Decline in visual attention and spatial memory in aged rats. Neurobiol. Aging 1999, 20, 605–615. [Google Scholar] [CrossRef]
- Moretti, M.; de Souza, A.G.; de Chaves, G.; de Andrade, V.M.; Romao, P.R.T.; Gavioli, E.C.; Boeck, C.R. Emotional behavior in middle-aged rats: Implications for geriatric psychopathologies. Physiol. Behav. 2011, 102, 115–120. [Google Scholar] [CrossRef]
- Boguszewski, P.; Zagrodzka, J. Emotional changes related to age in rats—A behavioral analysis. Behav. Brain Res. 2002, 133, 323–332. [Google Scholar] [CrossRef]
- Cacioppo, J.T.; Hawkley, L.C. Perceived social isolation and cognition. Trends Cogn. Sci. 2009, 13, 447–454. [Google Scholar] [CrossRef]
- Wilson, R.S.; Krueger, K.R.; Arnold, S.E.; Schneider, J.A.; Kelly, J.F.; Barnes, L.L.; Tang, Y.; Bennett, D.A. Loneliness and risk of Alzheimer disease. Arch. Gen. Psychiatry 2007, 64, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Landmann, H.; Rohmann, A. When loneliness dimensions drift apart: Emotional, social and physical loneliness during the COVID-19 lockdown and its associations with age, personality, stress and well-being. Int. J. Psychol. 2021, 57, 63–72. [Google Scholar] [CrossRef]
- Hawton, A.; Green, C.; Dickens, A.P.; Richards, S.H.; Taylor, R.S.; Edwards, R.; Greaves, C.J.; Campbell, J.L. The impact of social isolation on the health status and health-related quality of life of older people. Qual. Life Res. 2011, 20, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Patterson-Kane, E.G.; Hunt, M.; Harper, D. Rats demand social contact. Anim. Welf. 2002, 11, 327–332. [Google Scholar] [CrossRef]
- Arakawa, H. Ethological approach to social isolation effects in behavioral studies of laboratory rodents. Behav. Brain Res. 2018, 341, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Cacioppo, J.T.; Hawkley, L.C.; Norman, G.J.; Berntson, G.G. Social isolation. Ann. N. Y. Acad. Sci. 2011, 1231, 17. [Google Scholar] [CrossRef]
- Martin, A.L.; Brown, R.E. The lonely mouse: Verification of a separation-induced model of depression in female mice. Behav. Brain Res. 2010, 207, 196–207. [Google Scholar] [CrossRef]
- Ren, Q.G.; Gong, W.G.; Wang, Y.J.; Zhou, Q.D.; Zhang, Z.J. Citalopram attenuates tau hyperphosphorylation and spatial memory deficit induced by social isolation rearing in middle-aged rats. J. Mol. Neurosci. 2015, 56, 145–153. [Google Scholar] [CrossRef]
- Gong, W.G.; Wang, Y.J.; Zhou, H.; Li, X.L.; Bai, F.; Ren, Q.G.; Zhang, Z.J. Citalopram ameliorates synaptic plasticity deficits in different cognition-associated brain regions induced by social isolation in middle-aged rats. Mol. Neurobiol. 2017, 54, 1927–1938. [Google Scholar] [CrossRef]
- Pereda-Pérez, I.; Valencia, A.; Baliyan, S.; Núñez, Á.; Sanz-García, A.; Zamora, B.; Rodríguez-Fernández, R.; Esteban, J.A.; Venero, C. Systemic administration of a fibroblast growth factor receptor 1 agonist rescues the cognitive deficit in aged socially isolated rats. Neurobiol. Aging 2019, 78, 155–165. [Google Scholar] [CrossRef]
- Park, S.S.; Park, H.S.; Kim, T.W.; Lee, S.J. Effects of swimming exercise on social isolation-induced memory impairment and apoptosis in old rats. J. Exerc. Rehabil. 2020, 16, 234. [Google Scholar] [CrossRef] [PubMed]
- Pisu, M.G.; Garau, A.; Boero, G.; Biggio, F.; Pibiri, V.; Dore, R.; Locci, V.; Paci, E.; Porcu, P.; Serra, M. Sex differences in the outcome of juvenile social isolation on HPA axis function in rats. Neuroscience 2016, 320, 172–182. [Google Scholar] [CrossRef]
- Hermes, G.L.; Rosenthal, L.; Montag, A.; McClintock, M.K. Social isolation and the inflammatory response: Sex differences in the enduring effects of a prior stressor. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2006, 290, R273–R282. [Google Scholar] [CrossRef]
- Weintraub, A.; Singaravelu, J.; Bhatnagar, S. Enduring and sex-specific effects of adolescent social isolation in rats on adult stress reactivity. Brain Res. 2010, 1343, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Colucci-D’Amato, L.; Speranza, L.; Volpicelli, F. Neurotrophic factor BDNF, physiological functions and therapeutic potential in depression, neurodegeneration and brain cancer. Int. J. Mol. Sci. 2020, 21, 7777. [Google Scholar] [CrossRef] [PubMed]
- Zaletel, I.; Filipović, D.; Puškaš, N. Hippocampal BDNF in physiological conditions and social isolation. Rev. Neurosci. 2017, 28, 675–692. [Google Scholar] [CrossRef]
- Dwivedi, Y. Involvement of brain-derived neurotrophic factor in late-life depression. Am. J. Geriatr. Psychiatry 2013, 21, 433–449. [Google Scholar] [CrossRef]
- Şahin, T.D.; Karson, A.; Balcı, F.; Yazır, Y.; Bayramgürler, D.; Utkan, T. TNF-alpha inhibition prevents cognitive decline and maintains hippocampal BDNF levels in the unpredictable chronic mild stress rat model of depression. Behav. Brain Res. 2015, 292, 233–240. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, F.; Chen, J.; Dong, W. Chronic antidepressant administration alleviates frontal and hippocampal BDNF deficits in CUMS rat. Brain Res. 2010, 1366, 141–148. [Google Scholar] [CrossRef]
- Fang, K.; Li, H.R.; Chen, X.X.; Gao, X.R.; Huang, L.L.; Du, A.Q.; Jiang, C.; Li, H.; Ge, J.F. Quercetin alleviates LPS-induced depression-like behavior in rats via regulating BDNF-related imbalance of Copine 6 and TREM1/2 in the hippocampus and PFC. Front. Pharmacol. 2020, 10, 1544. [Google Scholar] [CrossRef]
- Rattiner, L.M.; Davis, M.; French, C.T.; Ressler, K.J. Brain-derived neurotrophic factor and tyrosine kinase receptor B involvement in amygdala-dependent fear conditioning. J. Neurosci. 2004, 24, 4796–4806. [Google Scholar] [CrossRef] [PubMed]
- Kozlovsky, N.; Matar, M.A.; Kaplan, Z.; Kotler, M.; Zohar, J.; Cohen, H. Long-term down-regulation of BDNF mRNA in rat hippocampal CA1 subregion correlates with PTSD-like behavioural stress response. Int. J. Neuropsychopharmacol. 2007, 10, 741–758. [Google Scholar] [CrossRef] [PubMed]
- Borges, J.V.; Pires, V.N.; de Freitas, B.S.; Rübensam, G.; Vieira, V.C.; Dos Santos, C.D.S.; Schröder, N.; Bromberg, E. Behavior, BDNF and epigenetic mechanisms in response to social isolation and social support in middle aged rats exposed to chronic stress. Behav. Brain Res. 2023, 441, 114303. [Google Scholar] [CrossRef] [PubMed]
- Giacobbo, B.L.; de Freitas, B.S.; Vedovelli, K.; Schlemmer, L.M.; Pires, V.N.; Antoniazzi, V.; Dos Santos, C.D.S.; Paludo, L.; Borges, J.V.; de Lima, D.B.; et al. Long-term environmental modifications affect BDNF concentrations in rat hippocampus, but not in serum. Behav. Brain Res. 2019, 372, 111965. [Google Scholar] [CrossRef]
- Bath, K.G.; Schilit, A.; Lee, F.S. Stress effects on BDNF expression: Effects of age, sex, and form of stress. Neuroscience 2013, 239, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Ter Horst, G.J.; Wichmann, R.; Bakker, P.; Liu, A.; Li, X.; Westenbroek, C. Sex differences in the effects of acute and chronic stress and recovery after long-term stress on stress-related brain regions of rats. Cereb. Cortex 2009, 19, 1978–1989. [Google Scholar] [CrossRef]
- Weisbrod, A.S.; Barry, E.S.; Graham, A.M.; Eklund, M.; Grunberg, N.E. Decreased BDNF in female but not male rats after exposure to stress: A sex-sensitive rat model of stress? Stress 2019, 22, 581–591. [Google Scholar] [CrossRef]
- Polyakova, M.; Stuke, K.; Schuemberg, K.; Mueller, K.; Schoenknecht, P.; Schroeter, M.L. BDNF as a biomarker for successful treatment of mood disorders: A systematic & quantitative meta-analysis. J. Affect. Disord. 2015, 174, 432–440. [Google Scholar]
- Murínová, J.; Hlaváčová, N.; Chmelová, M.; Riečanský, I. The evidence for altered BDNF expression in the brain of rats reared or housed in social isolation: A systematic review. Front. Behav. Neurosci. 2017, 11, 101. [Google Scholar] [CrossRef]
- Crippa, J.A.; Guimarães, F.S.; Campos, A.C.; Zuardi, A.W. Translational investigation of the therapeutic potential of cannabidiol (CBD): Toward a new age. Front. Immunol. 2018, 9, 2009. [Google Scholar] [CrossRef]
- Nahler, G.; Grotenhermen, F.; Zuardi, A.W.; Crippa, J.A. A conversion of oral cannabidiol to delta9-tetrahydrocannabinol seems not to occur in humans. Cannabis Cannabinoid Res. 2017, 2, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Bahji, A.; Meyyappan, A.C.; Hawken, E.R. Efficacy and acceptability of cannabinoids for anxiety disorders in adults: A systematic review & meta-analysis. J. Psychiatr. Res. 2020, 129, 257–264. [Google Scholar] [PubMed]
- Machado Bergamaschi, M.; Helena Costa Queiroz, R.; Waldo Zuardi, A.; Crippa, A.S. Safety and side effects of cannabidiol, a Cannabis sativa constituent. Curr. Drug Saf. 2011, 6, 237–249. [Google Scholar] [CrossRef]
- Blázquez, C.; Chiarlone, A.; Bellocchio, L.; Resel, E.; Pruunsild, P.; García-Rincón, D.; Sendtner, M.; Timmusk, T.; Lutz, B.; Galve-Roperh, I.; et al. The CB1 cannabinoid receptor signals striatal neuroprotection via a PI3K/Akt/mTORC1/BDNF pathway. Cell Death Differ. 2015, 22, 1618–1629. [Google Scholar] [CrossRef] [PubMed]
- Maison, P.; Walker, D.J.; Walsh, F.S.; Williams, G.; Doherty, P. BDNF regulates neuronal sensitivity to endocannabinoids. Neurosci. Lett. 2009, 467, 90–94. [Google Scholar] [CrossRef]
- Sales, A.J.; Fogaça, M.V.; Sartim, A.G.; Pereira, V.S.; Wegener, G.; Guimarães, F.S.; Joca, S.R. Cannabidiol induces rapid and sustained antidepressant-like effects through increased BDNF signaling and synaptogenesis in the prefrontal cortex. Mol. Neurobiol. 2019, 56, 1070–1081. [Google Scholar] [CrossRef]
- Bright, U.; Akirav, I. Cannabidiol Modulates Alterations in PFC microRNAs in a Rat Model of Depression. Int. J. Mol. Sci. 2023, 24, 2052. [Google Scholar] [CrossRef]
- Réus, G.Z.; Stringari, R.B.; Ribeiro, K.F.; Luft, T.; Abelaira, H.M.; Fries, G.R.; Aguiar, B.W.; Kapczinski, F.; Hallak, J.E.; Zuardi, A.W.; et al. Administration of cannabidiol and imipramine induces antidepressant-like effects in the forced swimming test and increases brain-derived neurotrophic factor levels in the rat amygdala. Acta Neuropsychiatr. 2011, 23, 241–248. [Google Scholar] [CrossRef]
- Matheson, J.; Bourgault, Z.; Le Foll, B. Sex differences in the neuropsychiatric effects and pharmacokinetics of cannabidiol: A scoping review. Biomolecules 2022, 12, 1462. [Google Scholar] [CrossRef]
- Yankelevitch-Yahav, R.; Franko, M.; Huly, A.; Doron, R. The forced swim test as a model of depressive-like behavior. JoVE J. Vis. Exp. 2015, 97, e52587. [Google Scholar]
- Palanza, P. Animal models of anxiety and depression: How are females different? Neurosci. Biobehav. Rev. 2001, 25, 219–233. [Google Scholar] [CrossRef]
- Campos, A.C.; Fogaça, M.V.; Aguiar, D.C.; Guimaraes, F.S. Animal models of anxiety disorders and stress. Braz. J. Psychiatry 2013, 35, S101–S111. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Tuohimaa, P. Experimental modeling of anxiety and depression. Acta Neurobiol. Exp. 2004, 64, 439–448. [Google Scholar]
- Deussing, J.M. Animal models of depression. Drug Discov. Today Dis. Models 2006, 3, 375–383. [Google Scholar] [CrossRef]
- Shbiro, L.; Hen-Shoval, D.; Hazut, N.; Rapps, K.; Dar, S.; Zalsman, G.; Mechoulam, R.; Weller, A.; Shoval, G. Effects of cannabidiol in males and females in two different rat models of depression. Physiol. Behav. 2019, 201, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Gáll, Z.; Farkas, S.; Albert, Á.; Ferencz, E.; Vancea, S.; Urkon, M.; Kolcsár, M. Effects of chronic cannabidiol treatment in the rat chronic unpredictable mild stress model of depression. Biomolecules 2020, 10, 801. [Google Scholar] [CrossRef]
- Moreira, F.A.; Aguiar, D.C.; Guimarães, F.S. Anxiolytic-like effect of cannabidiol in the rat Vogel conflict test. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2006, 30, 1466–1471. [Google Scholar] [CrossRef]
- Shoval, G.; Shbiro, L.; Hershkovitz, L.; Hazut, N.; Zalsman, G.; Mechoulam, R.; Weller, A. Prohedonic effect of cannabidiol in a rat model of depression. Neuropsychobiology 2016, 73, 123–129. [Google Scholar] [CrossRef]
- Gomes, F.V.; Reis, D.G.; Alves, F.H.; Corrêa, F.M.; Guimaraes, F.S.; Resstel, L.B. Cannabidiol injected into the bed nucleus of the stria terminalis reduces the expression of contextual fear conditioning via 5-HT1A receptors. J. Psychopharmacol. 2012, 26, 104–113. [Google Scholar] [CrossRef]
- Franzen, J.M.; Werle, I.; Vanz, F.; de Oliveira, B.B.; Nascimento, L.M.M.; Guimarães, F.S.; Bertoglio, L.J. Cannabidiol attenuates fear memory expression in female rats via hippocampal 5-HT1A but not CB1 or CB2 receptors. Neuropharmacology 2023, 223, 109316. [Google Scholar] [CrossRef]
- Kreilaus, F.; Przybyla, M.; Ittner, L.; Karl, T. Cannabidiol (CBD) treatment improves spatial memory in 14-month-old female TAU58/2 transgenic mice. Behav. Brain Res. 2022, 425, 113812. [Google Scholar] [CrossRef] [PubMed]
- Franzen, J.M.; Vanz, F.; Werle, I.; Guimarães, F.S.; Bertoglio, L.J. Cannabidiol impairs fear memory reconsolidation in female rats through dorsal hippocampus CB1 but not CB2 receptor interaction. Eur. Neuropsychopharmacol. 2022, 64, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Mottarlini, F.; Fumagalli, M.; Castillo-Díaz, F.; Piazza, S.; Targa, G.; Sangiovanni, E.; Pacchetti, B.; Sodergren, M.H.; Dell’Agli, M.; Fumagalli, F.; et al. Single and repeated exposure to cannabidiol differently modulate BDNF expression and signaling in the Cortico-Striatal Brain Network. Biomedicines 2022, 10, 1853. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Kesner, R.P. Time-dependent relationship between the dorsal hippocampus and the prefrontal cortex in spatial memory. J. Neurosci. 2003, 23, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Churchwell, J.C.; Morris, A.M.; Musso, N.D.; Kesner, R.P. Prefrontal and hippocampal contributions to encoding and retrieval of spatial memory. Neurobiol. Learn. Mem. 2010, 93, 415–421. [Google Scholar] [CrossRef]
- Yu, H.; Chen, Z.Y. The role of BDNF in depression on the basis of its location in the neural circuitry. Acta Pharmacol. Sin. 2011, 32, 3–11. [Google Scholar] [CrossRef]
- Groves, J.O. Is it time to reassess the BDNF hypothesis of depression? Mol. Psychiatry 2007, 12, 1079–1088. [Google Scholar] [CrossRef]
- Berton, O.; McClung, C.A.; Dileone, R.J.; Krishnan, V.; Renthal, W.; Russo, S.J.; Graham, D.; Tsankova, N.M.; Bolanos, C.A.; Rios, M.; et al. Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science 2006, 311, 864–868. [Google Scholar] [CrossRef]
- McEwen, B.S.; Nasca, C.; Gray, J.D. Stress effects on neuronal structure: Hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology 2016, 41, 3–23. [Google Scholar] [CrossRef]
- Gray, J.D.; Milner, T.A.; McEwen, B.S. Dynamic plasticity: The role of glucocorticoids, brain-derived neurotrophic factor and other trophic factors. Neuroscience 2013, 239, 214–227. [Google Scholar] [CrossRef]
- Bennett, M.R.; Lagopoulos, J. Stress and trauma: BDNF control of dendritic-spine formation and regression. Prog. Neurobiol. 2014, 112, 80–99. [Google Scholar] [CrossRef] [PubMed]
- Mallei, A.; Ieraci, A.; Popoli, M. Chronic social defeat stress differentially regulates the expression of BDNF transcripts and epigenetic modifying enzymes in susceptible and resilient mice. World J. Biol. Psychiatry 2018, 20, 555–566. [Google Scholar] [CrossRef]
- Han, X.; Wang, W.; Xue, X.; Shao, F.; Li, N. Brief social isolation in early adolescence affects reversal learning and forebrain BDNF expression in adult rats. Brain Res. Bull. 2011, 86, 173–178. [Google Scholar] [CrossRef]
- Pizarro, J.M.; Lumley, L.A.; Medina, W.; Robison, C.L.; Chang, W.E.; Alagappan, A.; Bah, M.J.; Dawood, M.Y.; Shah, J.D.; Mark, B.; et al. Acute social defeat reduces neurotrophin expression in brain cortical and subcortical areas in mice. Brain Res. 2004, 1025, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Fanous, S.; Hammer, R.P., Jr.; Nikulina, E.M. Short- and long-term effects of intermittent social defeat stress on brain-derived neurotrophic factor expression in mesocorticolimbic brain regions. Neuroscience 2010, 167, 598–607. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.; Sun, Y.; McGregor, A.; Connor, B. Allopregnanolone regulates neurogenesis and depressive/anxiety-like behaviour in a social isolation rodent model of chronic stress. Neuropharmacology 2012, 63, 1315–1326. [Google Scholar] [CrossRef]
- Hellweg, R.; Zueger, M.; Fink, K.; Hörtnagl, H.; Gass, P. Olfactory bulbectomy in mice leads to increased BDNF levels and decreased serotonin turnover in depression-related brain areas. Neurobiol. Dis. 2007, 25, 1–7. [Google Scholar] [CrossRef]
- Sales, A.J.; Crestani, C.C.; Guimarães, F.S.; Joca, S.R. Antidepressant-like effect induced by Cannabidiol is dependent on brain serotonin levels. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 86, 255–261. [Google Scholar] [CrossRef]
- Sales, A.J.; Guimarães, F.S.; Joca, S.R. CBD modulates DNA methylation in the prefrontal cortex and hippocampus of mice exposed to forced swim. Behav. Brain Res. 2020, 388, 112627. [Google Scholar] [CrossRef]
- Cruces, J.; Venero, C.; Pereda-Pérez, I.; De la Fuente, M. A higher anxiety state in old rats after social isolation is associated to an impairment of the immune response. J. Neuroimmunol. 2014, 277, 18–25. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, Hard Cover Edition; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Portugalov, A.; Zaidan, H.; Gaisler-Salomon, I.; Hillard, C.J.; Akirav, I. FAAH Inhibition Restores Early Life Stress-Induced Alterations in PFC microRNAs Associated with Depressive-Like Behavior in Male and Female Rats. Int. J. Mol. Sci. 2022, 23, 16101. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mPFC (n = 28) | CA1 (n = 32) | VSUB (n = 34) | BLA (n = 25) | NAc (n = 25) | |
|---|---|---|---|---|---|
| OFT—time in center | r = 0.422 p = 0.025 | r = −0.152 p = 0.406 | r = −0.172 p = 0.330 | r = 0.433 p = 0.027 | r = 0.431 p = 0.032 |
| OFT—freezing | r = 0.075 p = 0.73 | r = −0.183 p = 0.316 | r = 0.089 p = 0.617 | r = −0.214 p = 0.305 | r = −0.395 p = 0.051 |
| MWM test—training days average | r = −0.522 p = 0.004 | r = −0.101 p = 0.581 | r = −0.063 p = 0.723 | r = −0.230 p = 0.269 | r = −0.135 p = 0.519 |
| MWM probe (day 4) | r = 0.437 p = 0.02 | r = 0.105 p = 0.567 | r = −0.123 p = 0.489 | r = −0.001 p = 0.996 | r = −0.257 p = 0.215 |
| MWM probe (day 5) | r = 0.353 p = 0.065 | r = 0.027 p = 0.881 | r = 0.046 p = 0.796 | r = 0.222 p = 0.286 | r = 0.030 p = 0.887 |
| FST—immobility | r = 0.272 p = 0.161 | r = 0.005 p = 0.980 | r = 0.071 p = 0.692 | r = −0.046 p = 0.828 | r = −0.383 p = 0.059 |
| FST—climbing | r = −0.001 p = 0.995 | r = −0.14 p = 0.941 | r = 0.136 p = 0.443 | r = 0.391 p = 0.053 | r = 0.724 p = <0.001 |
| FST—swimming | r = −0.325 p = 0.092 | r = 0.004 p = 0.981 | r = −0.155 p = 0.380 | r = −0.187 p = 0.370 | r = −0.004 p = 0.984 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saad, N.; Raviv, D.; Mizrachi Zer-Aviv, T.; Akirav, I. Cannabidiol Modulates Emotional Function and Brain-Derived Neurotrophic Factor Expression in Middle-Aged Female Rats Exposed to Social Isolation. Int. J. Mol. Sci. 2023, 24, 15492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242015492
Saad N, Raviv D, Mizrachi Zer-Aviv T, Akirav I. Cannabidiol Modulates Emotional Function and Brain-Derived Neurotrophic Factor Expression in Middle-Aged Female Rats Exposed to Social Isolation. International Journal of Molecular Sciences. 2023; 24(20):15492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242015492
Chicago/Turabian StyleSaad, Nadya, Danielle Raviv, Tomer Mizrachi Zer-Aviv, and Irit Akirav. 2023. "Cannabidiol Modulates Emotional Function and Brain-Derived Neurotrophic Factor Expression in Middle-Aged Female Rats Exposed to Social Isolation" International Journal of Molecular Sciences 24, no. 20: 15492. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242015492