
Differential Influences of Endogenous and Exogenous Sensory Neuropeptides on the ATP Metabolism by Soluble Ectonucleotidases in the Murine Bladder Lamina Propria
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
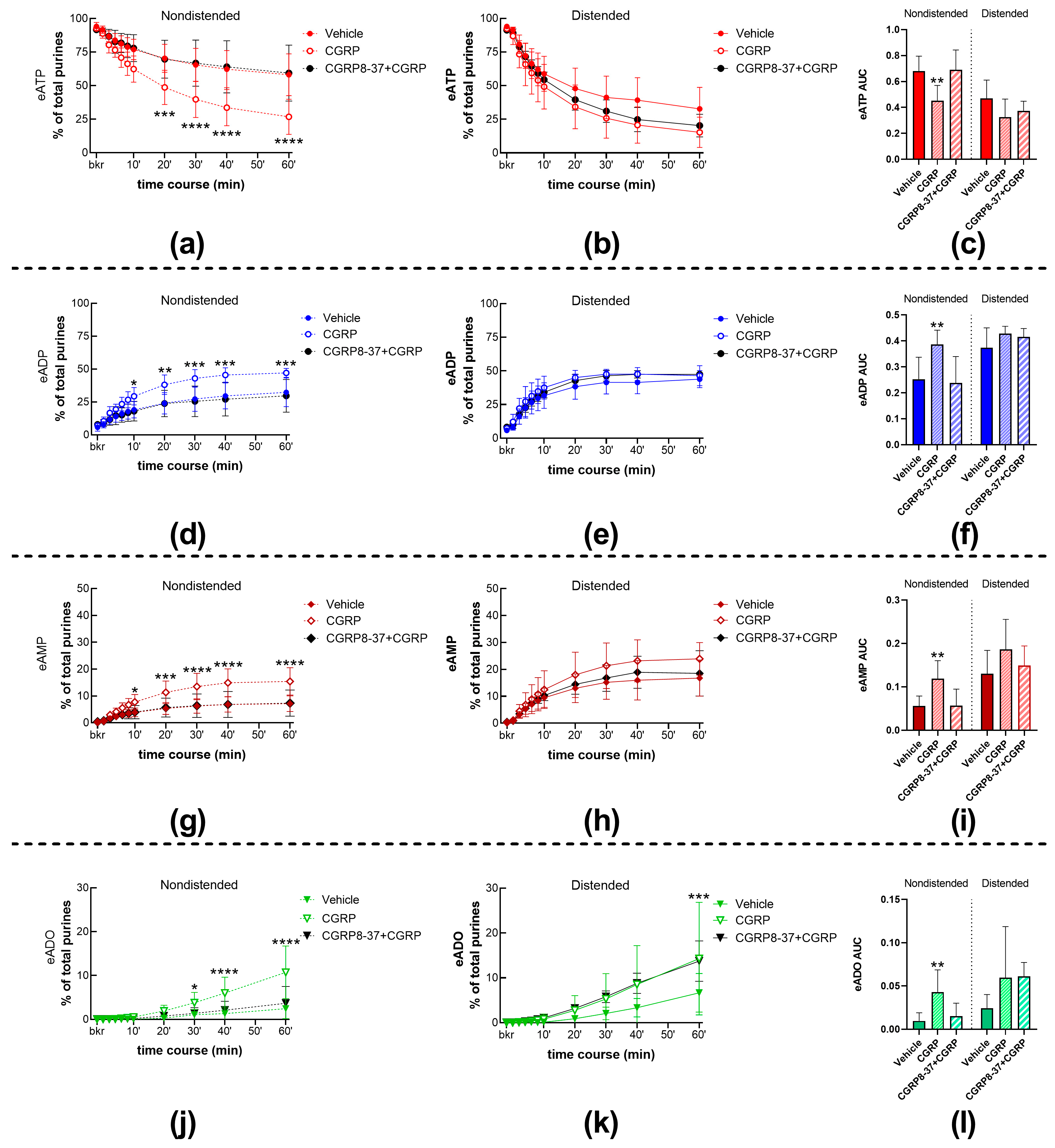
2.1. Role of CGRP in Spontaneous and Distention-Induced Release of s-ENTDs in the LP
2.1.1. Exogenous CGRP Increases the Spontaneous but Not the Distention-Induced Hydrolysis of eATP by s-ENTDs
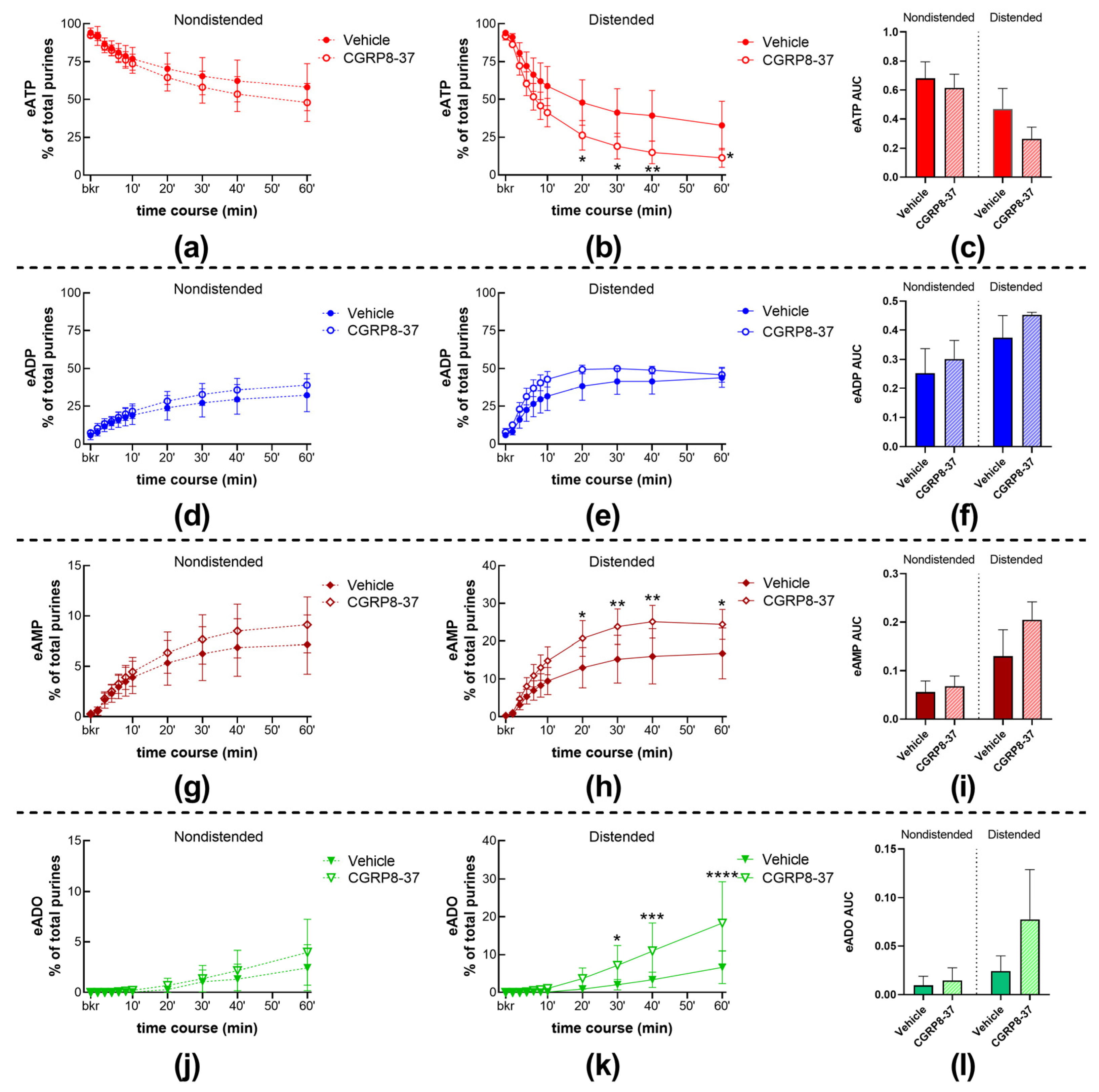
2.1.2. The Effect of CGRP on Spontaneous s-ENTD Release Is Mediated by Receptors Sensitive to CGRP8-37
2.1.3. Inhibition of CGRP Receptors with CGRP8-37 Increased the Distention-Induced Release of s-ENTDs but Not the Spontaneous Release of Enzymes
2.2. Role of the NK1 Receptor in Spontaneous and Distention-Induced Release of s-ENTDs in the LP
2.2.1. Exogenous Substance P Increases the Spontaneous but Not the Distention-Induced Hydrolysis of eATP by s-ENTDs
2.2.2. The NK1 Receptor Inhibitor CP122721 Increases eATP Hydrolysis in the LP of Nondistended and Distended Bladders
2.3. Role of the NK2 Receptor on Spontaneous and Distention-Induced Release of s-ENTDs in the LP
2.3.1. The NK2 Receptor Agonist NKA Increases eATP Hydrolysis in the LP of Nondistended and Distended Bladders
2.3.2. The NK2 Receptor Antagonist MEN 10376 Does Not Affect the Spontaneous or Distention-Induced Release of s-ENTDs in the LP
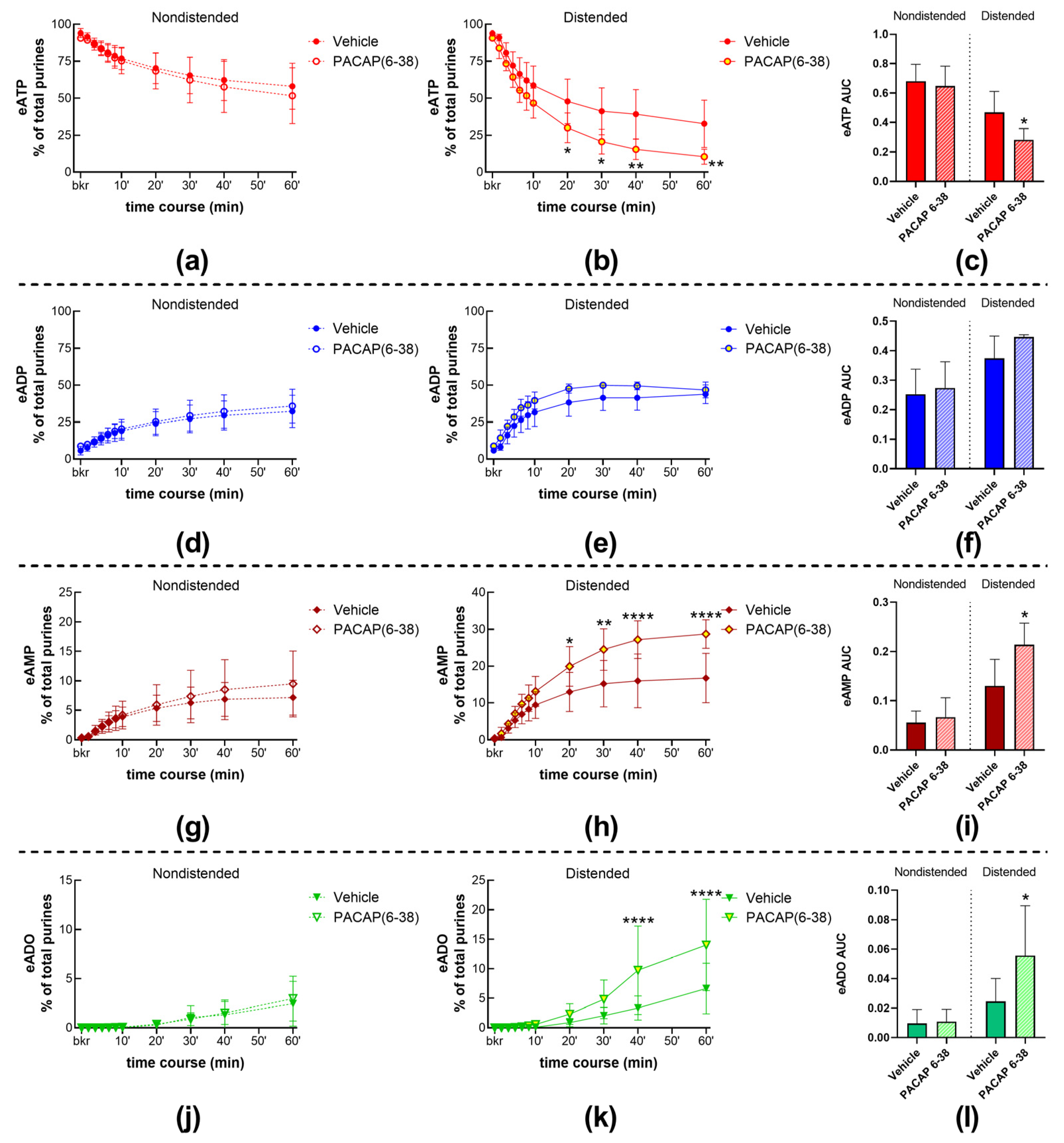
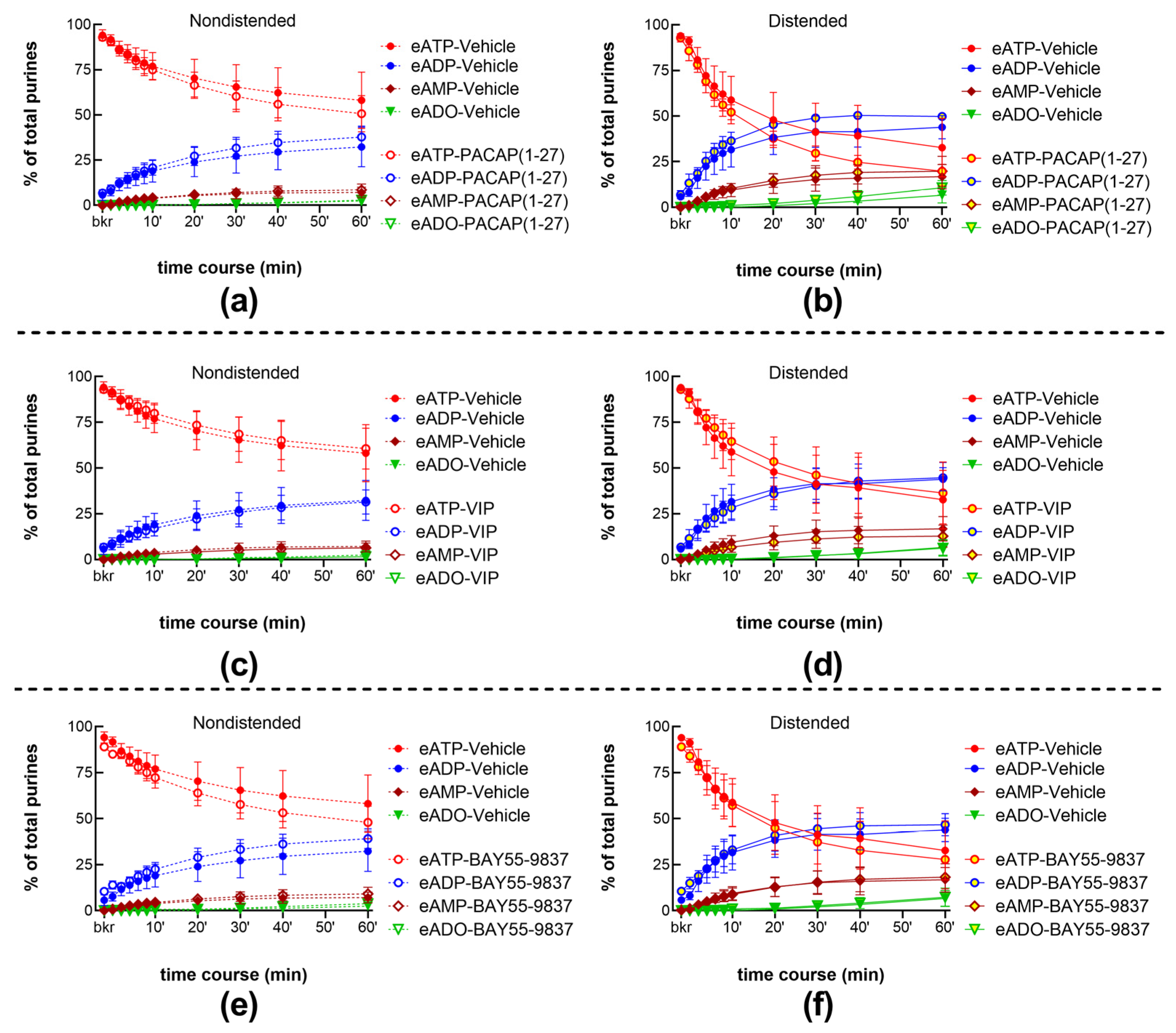
2.4. Role of the PAC1 Receptor in s-ENTD Release in the LP
2.4.1. Exogenous PACAP (1-38) Increases the Distention-Induced Release of s-ENTDs but Not the Spontaneous Release of Enzymes in the LP
2.4.2. Inhibition of the PAC1 Receptor with PACAP (6-38) Increased the Distention-Induced Release of s-ENTDs but Not the Spontaneous Release of s-ENTDs in the LP
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Ex Vivo Detrusor-Free Bladder Preparation
4.3. Collection of Extraluminal Solutions Containing s-ENTDs
4.4. Preparation of Reaction Solutions Containing Released s-ENTDs
4.5. Time-Course of eATP Hydrolysis by s-ENTDs in cELS from Nondistended and Distended Bladder Preparations
4.6. Chemical Synthesis of the 1,N6-Etheno-ATP (eATP) Substrate
4.7. HPLC Analysis of 1,N6-Etheno-Nucleotides and 1,N6-Etheno-Nucleosides
4.8. Drugs
4.9. Statistical Analyses of the Results
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Durnin, L.; Kwok, B.; Kukadia, P.; McAvera, R.; Corrigan, R.D.; Ward, S.M.; Zhang, Y.; Chen, Q.; Koh, S.D.; Sanders, K.M.; et al. An ex vivo bladder model with detrusor smooth muscle removed to analyse biologically active mediators released from the suburothelium. J. Physiol. 2019, 597, 1467–1485. [Google Scholar] [CrossRef]
- Ferguson, D.R.; Kennedy, I.; Burton, T.J. ATP is released from rabbit urinary bladder epithelial cells by hydrostatic pressure changes—A possible sensory mechanism? J. Physiol. 1997, 505, 503–511. [Google Scholar] [CrossRef]
- Matsumoto-Miyai, K.; Kagase, A.; Yamada, E.; Yoshizumi, M.; Murakami, M.; Ohba, T.; Kawatani, M. Store-operated Ca2+ entry suppresses distention-induced ATP release from the urothelium. Am. J. Physiol. Renal Physiol. 2011, 300, F716–F720. [Google Scholar] [CrossRef]
- McLatchie, L.M.; Fry, C.H. ATP release from freshly isolated guinea-pig bladder urothelial cells: A quantification and study of the mechanisms involved. BJU Int. 2015, 115, 987–993. [Google Scholar] [CrossRef]
- Wang, E.C.; Lee, J.M.; Ruiz, W.G.; Balestreire, E.M.; von Bodungen, M.; Barrick, S.; Cockayne, D.A.; Birder, L.A.; Apodaca, G. ATP and purinergic receptor-dependent membrane traffic in bladder umbrella cells. J. Clin. Investig. 2005, 115, 2412–2422. [Google Scholar] [CrossRef]
- Merrill, L.; Gonzalez, E.J.; Girard, B.M.; Vizzard, M.A. Receptors, channels, and signalling in the urothelial sensory system in the bladder. Nat. Rev. Urol. 2016, 13, 193–204. [Google Scholar] [CrossRef]
- Dalghi, M.G.; Montalbetti, N.; Carattino, M.D.; Apodaca, G. The Urothelium: Life in a Liquid Environment. Physiol. Rev. 2020, 100, 1621–1705. [Google Scholar] [CrossRef]
- Burnstock, G. Purinergic signalling in the urinary tract in health and disease. Purinergic. Signal. 2014, 10, 103–155. [Google Scholar] [CrossRef]
- de Groat, W.C.; Griffiths, D.; Yoshimura, N. Neural control of the lower urinary tract. Compr. Physiol. 2015, 5, 327–396. [Google Scholar]
- Zimmermann, H.; Zebisch, M.; Strater, N. Cellular function and molecular structure of ecto-nucleotidases. Purinergic. Signal. 2012, 8, 437–502. [Google Scholar] [CrossRef]
- Aresta Branco, M.S.L.; Gutierrez Cruz, A.; Dayton, J.; Perrino, B.A.; Mutafova-Yambolieva, V.N. Mechanosensitive Hydrolysis of ATP and ADP in Lamina Propria of the Murine Bladder by Membrane-Bound and Soluble Nucleotidases. Front. Physiol. 2022, 13, 918100. [Google Scholar] [CrossRef] [PubMed]
- Gabella, G. Afferent nerve fibres in the wall of the rat urinary bladder. Cell Tissue Res. 2019, 376, 25–35. [Google Scholar] [CrossRef]
- Fowler, C.J.; Griffiths, D.; de Groat, W.C. The neural control of micturition. Nat. Rev. Neurosci. 2008, 9, 453–466. [Google Scholar] [CrossRef]
- Girard, B.M.; Wolf-Johnston, A.; Braas, K.M.; Birder, L.A.; May, V.; Vizzard, M.A. PACAP-mediated ATP release from rat urothelium and regulation of PACAP/VIP and receptor mRNA in micturition pathways after cyclophosphamide (CYP)-induced cystitis. J. Mol. Neurosci. 2008, 36, 310–320. [Google Scholar] [CrossRef]
- Arms, L.; Vizzard, M.A. Neuropeptides in lower urinary tract function. Handb. Exp. Pharmacol. 2011, 202, 395–423. [Google Scholar] [CrossRef]
- Shaffer, A.D.; Ball, C.L.; Robbins, M.T.; Ness, T.J.; Randich, A. Effects of acute adult and early-in-life bladder inflammation on bladder neuropeptides in adult female rats. BMC Urol. 2011, 11, 18. [Google Scholar] [CrossRef]
- Zheng, G.; Harms, A.K.; Tail, M.; Zhang, H.; Nimmo, A.; Skutella, T.; Kiening, K.; Unterberg, A.; Zweckberger, K.; Younsi, A. Effects of a neurokinin-1 receptor antagonist in the acute phase after thoracic spinal cord injury in a rat model. Front. Mol. Neurosci. 2023, 16, 1128545. [Google Scholar] [CrossRef]
- Holzer, P. Neurogenic vasodilatation and plasma leakage in the skin. Gen. Pharmacol. 1998, 30, 5–11. [Google Scholar] [CrossRef]
- Khera, M.; Somogyi, G.T.; Kiss, S.; Boone, T.B.; Smith, C.P. Botulinum toxin A inhibits ATP release from bladder urothelium after chronic spinal cord injury. Neurochem. Int. 2004, 45, 987–993. [Google Scholar] [CrossRef]
- Smith, C.P.; Vemulakonda, V.M.; Kiss, S.; Boone, T.B.; Somogyi, G.T. Enhanced ATP release from rat bladder urothelium during chronic bladder inflammation: Effect of botulinum toxin A. Neurochem. Int. 2005, 47, 291–297. [Google Scholar] [CrossRef]
- Aresta Branco, M.S.L.; Gutierrez Cruz, A.; Borhani Peikani, M.; Mutafova-Yambolieva, V.N. Sensory Neurons, PIEZO Channels and PAC1 Receptors Regulate the Mechanosensitive Release of Soluble Ectonucleotidases in the Murine Urinary Bladder Lamina Propria. Int. J. Mol. Sci. 2023, 24, 7322. [Google Scholar] [CrossRef]
- Aresta Branco, M.S.L.; Gutierrez Cruz, A.; Peri, L.E.; Mutafova-Yambolieva, V.N. The Pannexin 1 Channel and the P2X7 Receptor Are in Complex Interplay to Regulate the Release of Soluble Ectonucleotidases in the Murine Bladder Lamina Propria. Int. J. Mol. Sci. 2023, 24, 9964. [Google Scholar] [CrossRef]
- Chiba, T.; Yamaguchi, A.; Yamatani, T.; Nakamura, A.; Morishita, T.; Inui, T.; Fukase, M.; Noda, T.; Fujita, T. Calcitonin gene-related peptide receptor antagonist human CGRP-(8–37). Am. J. Physiol. 1989, 256, E331–E335. [Google Scholar] [CrossRef]
- McLean, S.; Ganong, A.; Seymour, P.A.; Bryce, D.K.; Crawford, R.T.; Morrone, J.; Reynolds, L.S.; Schmidt, A.W.; Zorn, S.; Watson, J.; et al. Characterization of CP-122,721; a nonpeptide antagonist of the neurokinin NK1 receptor. J. Pharmacol. Exp. Ther. 1996, 277, 900–908. [Google Scholar]
- Maggi, C.A.; Giuliani, S.; Ballati, L.; Lecci, A.; Manzini, S.; Patacchini, R.; Renzetti, A.R.; Rovero, P.; Quartara, L.; Giachetti, A. In vivo evidence for tachykininergic transmission using a new NK-2 receptor-selective antagonist, MEN 10,376. J. Pharmacol. Exp. Ther. 1991, 257, 1172–1178. [Google Scholar]
- Robberecht, P.; Gourlet, P.; De Neef, P.; Woussen-Colle, M.C.; Vandermeers-Piret, M.C.; Vandermeers, A.; Christophe, J. Structural requirements for the occupancy of pituitary adenylate-cyclase-activating-peptide (PACAP) receptors and adenylate cyclase activation in human neuroblastoma NB-OK-1 cell membranes. Discovery of PACAP(6-38) as a potent antagonist. Eur. J. Biochem. 1992, 207, 239–246. [Google Scholar] [CrossRef]
- Smith-Anttila, C.J.A.; Morrison, V.; Keast, J.R. Spatiotemporal mapping of sensory and motor innervation of the embryonic and postnatal mouse urinary bladder. Dev. Biol. 2021, 476, 18–32. [Google Scholar] [CrossRef]
- Schueth, A.; Spronck, B.; van Zandvoort, M.; van Koeveringe, G.A. Computer-assisted three-dimensional tracking of sensory innervation in the murine bladder mucosa with two-photon microscopy. J. Chem. Neuroanat. 2017, 85, 43–49. [Google Scholar] [CrossRef]
- Andersson, K.E.; McCloskey, K.D. Lamina propria: The functional center of the bladder? Neurourol. Urodyn. 2014, 33, 9–16. [Google Scholar] [CrossRef]
- Steinhoff, M.S.; von Mentzer, B.; Geppetti, P.; Pothoulakis, C.; Bunnett, N.W. Tachykinins and their receptors: Contributions to physiological control and the mechanisms of disease. Physiol. Rev. 2014, 94, 265–301. [Google Scholar] [CrossRef]
- Birder, L.A.; Kullmann, F.A. Role of neurogenic inflammation in local communication in the visceral mucosa. Semin. Immunopathol. 2018, 40, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Birder, L.; Andersson, K.E. Urothelial signaling. Physiol. Rev. 2013, 93, 653–680. [Google Scholar] [CrossRef] [PubMed]
- Hammond, T.G.; Saban, R.; Bost, K.L.; Harris, H.W., Jr.; Kaysen, J.H.; Goda, F.O.; Wang, X.C.; Lewis, F.C.; Navar, G.L.; Campbell, W.C.; et al. Substance P dependence of endosomal fusion during bladder inflammation. Am. J. Physiol. Renal Physiol. 2000, 278, F440–F451. [Google Scholar] [CrossRef]
- Ojala, J.; Tooke, K.; Hsiang, H.; Girard, B.M.; May, V.; Vizzard, M.A. PACAP/PAC1 Expression and Function in Micturition Pathways. J. Mol. Neurosci. 2019, 68, 357–367. [Google Scholar] [CrossRef]
- McLatchie, L.M.; Fraser, N.J.; Main, M.J.; Wise, A.; Brown, J.; Thompson, N.; Solari, R.; Lee, M.G.; Foord, S.M. RAMPs regulate the transport and ligand specificity of the calcitonin-receptor-like receptor. Nature 1998, 393, 333–339. [Google Scholar] [CrossRef]
- Roehrkasse, A.M.; Booe, J.M.; Lee, S.M.; Warner, M.L.; Pioszak, A.A. Structure-function analyses reveal a triple β-turn receptor-bound conformation of adrenomedullin 2/intermedin and enable peptide antagonist design. J. Biol. Chem. 2018, 293, 15840–15854. [Google Scholar] [CrossRef]
- Doods, H.; Hallermayer, G.; Wu, D.; Entzeroth, M.; Rudolf, K.; Engel, W.; Eberlein, W. Pharmacological profile of BIBN4096BS, the first selective small molecule CGRP antagonist. Br. J. Pharmacol. 2000, 129, 420–423. [Google Scholar] [CrossRef]
- Rudolf, K.; Eberlein, W.; Engel, W.; Pieper, H.; Entzeroth, M.; Hallermayer, G.; Doods, H. Development of human calcitonin gene-related peptide (CGRP) receptor antagonists. 1. Potent and selective small molecule CGRP antagonists. 1-[N2-[3,5-dibromo-N-[[4-(3,4-dihydro-2(1H)-oxoquinazolin-3-yl)-1-piperidinyl]carbonyl]-d-tyrosyl]-l-lysyl]-4-(4-pyridinyl)piperazine: The first CGRP antagonist for clinical trials in acute migraine. J. Med. Chem. 2005, 48, 5921–5931. [Google Scholar] [CrossRef]
- Harmar, A.J.; Fahrenkrug, J.; Gozes, I.; Laburthe, M.; May, V.; Pisegna, J.R.; Vaudry, D.; Vaudry, H.; Waschek, J.A.; Said, S.I. Pharmacology and functions of receptors for vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide: IUPHAR review 1. Br. J. Pharmacol. 2012, 166, 4–17. [Google Scholar] [CrossRef]
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- Tsutsumi, M.; Claus, T.H.; Liang, Y.; Li, Y.; Yang, L.; Zhu, J.; Dela Cruz, F.; Peng, X.; Chen, H.; Yung, S.L.; et al. A potent and highly selective VPAC2 agonist enhances glucose-induced insulin release and glucose disposal: A potential therapy for type 2 diabetes. Diabetes 2002, 51, 1453–1460. [Google Scholar] [CrossRef] [PubMed]
- Braas, K.M.; May, V.; Zvara, P.; Nausch, B.; Kliment, J.; Dunleavy, J.D.; Nelson, M.T.; Vizzard, M.A. Role for pituitary adenylate cyclase activating polypeptide in cystitis-induced plasticity of micturition reflexes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290, R951–R962. [Google Scholar] [CrossRef] [PubMed]
- Girard, B.M.; Malley, S.E.; Braas, K.M.; May, V.; Vizzard, M.A. PACAP/VIP and receptor characterization in micturition pathways in mice with overexpression of NGF in urothelium. J. Mol. Neurosci. 2010, 42, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Rapp, D.E.; Turk, K.W.; Bales, G.T.; Cook, S.P. Botulinum toxin type a inhibits calcitonin gene-related peptide release from isolated rat bladder. J. Urol. 2006, 175, 1138–1142. [Google Scholar] [CrossRef]
- Lucioni, A.; Bales, G.T.; Lotan, T.L.; McGehee, D.S.; Cook, S.P.; Rapp, D.E. Botulinum toxin type A inhibits sensory neuropeptide release in rat bladder models of acute injury and chronic inflammation. BJU Int. 2008, 101, 366–370. [Google Scholar] [CrossRef]
- Sun, Y.; Chai, T.C. Augmented extracellular ATP signaling in bladder urothelial cells from patients with interstitial cystitis. Am. J. Physiol. Cell Physiol. 2006, 290, C27–C34. [Google Scholar] [CrossRef]
- Taidi, Z.; Mansfield, K.J.; Bates, L.; Sana-Ur-Rehman, H.; Liu, L. Purinergic P2X7 receptors as therapeutic targets in interstitial cystitis/bladder pain syndrome; key role of ATP signaling in inflammation. Bladder 2019, 6, e38. [Google Scholar] [CrossRef]
- Lichtenthaler, S.F.; Lemberg, M.K.; Fluhrer, R. Proteolytic ectodomain shedding of membrane proteins in mammals-hardware, concepts, and recent developments. EMBO J. 2018, 37, e99456. [Google Scholar] [CrossRef]
- Weston, C.; Winfield, I.; Harris, M.; Hodgson, R.; Shah, A.; Dowell, S.J.; Mobarec, J.C.; Woodlock, D.A.; Reynolds, C.A.; Poyner, D.R.; et al. Receptor Activity-modifying Protein-directed G Protein Signaling Specificity for the Calcitonin Gene-related Peptide Family of Receptors. J. Biol. Chem. 2016, 291, 21925–21944. [Google Scholar] [CrossRef]
- Sun, J.; Ramnath, R.D.; Tamizhselvi, R.; Bhatia, M. Neurokinin A engages neurokinin-1 receptor to induce NF-kappaB-dependent gene expression in murine macrophages: Implications of ERK1/2 and PI 3-kinase/Akt pathways. Am. J. Physiol. Cell Physiol. 2008, 295, C679–C691. [Google Scholar] [CrossRef]
- Sánchez, M.L.; Rodríguez, F.D.; Coveñas, R. Peptidergic Systems and Cancer: Focus on Tachykinin and Calcitonin/Calcitonin Gene-Related Peptide Families. Cancers 2023, 15, 1694. [Google Scholar] [CrossRef] [PubMed]
- Reiter, E.; Ahn, S.; Shukla, A.K.; Lefkowitz, R.J. Molecular mechanism of β-arrestin-biased agonism at seven-transmembrane receptors. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 179–197. [Google Scholar] [CrossRef] [PubMed]
- May, V.; Parsons, R.L. G Protein-Coupled Receptor Endosomal Signaling and Regulation of Neuronal Excitability and Stress Responses: Signaling Options and Lessons from the PAC1 Receptor. J. Cell. Physiol. 2017, 232, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Alexander, T.I.; Tasma, Z.; Siow, A.; Rees, T.A.; Brimble, M.A.; Harris, P.W.R.; Hay, D.L.; Walker, C.S. Novel Fluorescently Labeled PACAP and VIP Highlight Differences between Peptide Internalization and Receptor Pharmacology. ACS Pharmacol. Transl. Sci. 2023, 6, 52–64. [Google Scholar] [CrossRef]
- Gutierrez Cruz, A.; Aresta Branco, M.S.L.; Perrino, B.A.; Sanders, K.M.; Mutafova-Yambolieva, V.N. Urinary ATP Levels Are Controlled by Nucleotidases Released from the Urothelium in a Regulated Manner. Metabolites 2022, 13, 30. [Google Scholar] [CrossRef]
- Bobalova, J.; Bobal, P.; Mutafova-Yambolieva, V.N. High-Performance Liquid Chromatographic Technique for Detection of a Fluorescent Analogue of ADP-Ribose in Isolated Blood Vessel Preparations. Anal. Biochem. 2002, 305, 269–276. [Google Scholar] [CrossRef]
- Levitt, B.; Head, R.J.; Westfall, D.P. High-pressure liquid chromatographic-fluorometric detection of adenosine and adenine nucleotides: Application to endogenous content and electrically induced release of adenyl purines in guinea pig vas deferens. Anal. Biochem. 1984, 137, 93–100. [Google Scholar] [CrossRef]
- Vollert, J.; Schenker, E.; Macleod, M.; Bespalov, A.; Wuerbel, H.; Michel, M.; Dirnagl, U.; Potschka, H.; Waldron, A.M.; Wever, K.; et al. Systematic review of guidelines for internal validity in the design, conduct and analysis of preclinical biomedical experiments involving laboratory animals. BMJ Open Sci. 2020, 4, e100046. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutierrez Cruz, A.; Aresta Branco, M.S.L.; Borhani Peikani, M.; Mutafova-Yambolieva, V.N. Differential Influences of Endogenous and Exogenous Sensory Neuropeptides on the ATP Metabolism by Soluble Ectonucleotidases in the Murine Bladder Lamina Propria. Int. J. Mol. Sci. 2023, 24, 15650. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242115650
Gutierrez Cruz A, Aresta Branco MSL, Borhani Peikani M, Mutafova-Yambolieva VN. Differential Influences of Endogenous and Exogenous Sensory Neuropeptides on the ATP Metabolism by Soluble Ectonucleotidases in the Murine Bladder Lamina Propria. International Journal of Molecular Sciences. 2023; 24(21):15650. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242115650
Chicago/Turabian StyleGutierrez Cruz, Alejandro, Mafalda S. L. Aresta Branco, Mahsa Borhani Peikani, and Violeta N. Mutafova-Yambolieva. 2023. "Differential Influences of Endogenous and Exogenous Sensory Neuropeptides on the ATP Metabolism by Soluble Ectonucleotidases in the Murine Bladder Lamina Propria" International Journal of Molecular Sciences 24, no. 21: 15650. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242115650