
Bisphenol AF Induces Prostatic Dorsal Lobe Hyperplasia in Rats through Activation of the NF-κB Signaling Pathway
 , ,
, ,
Abstract
:1. Introduction
2. Results
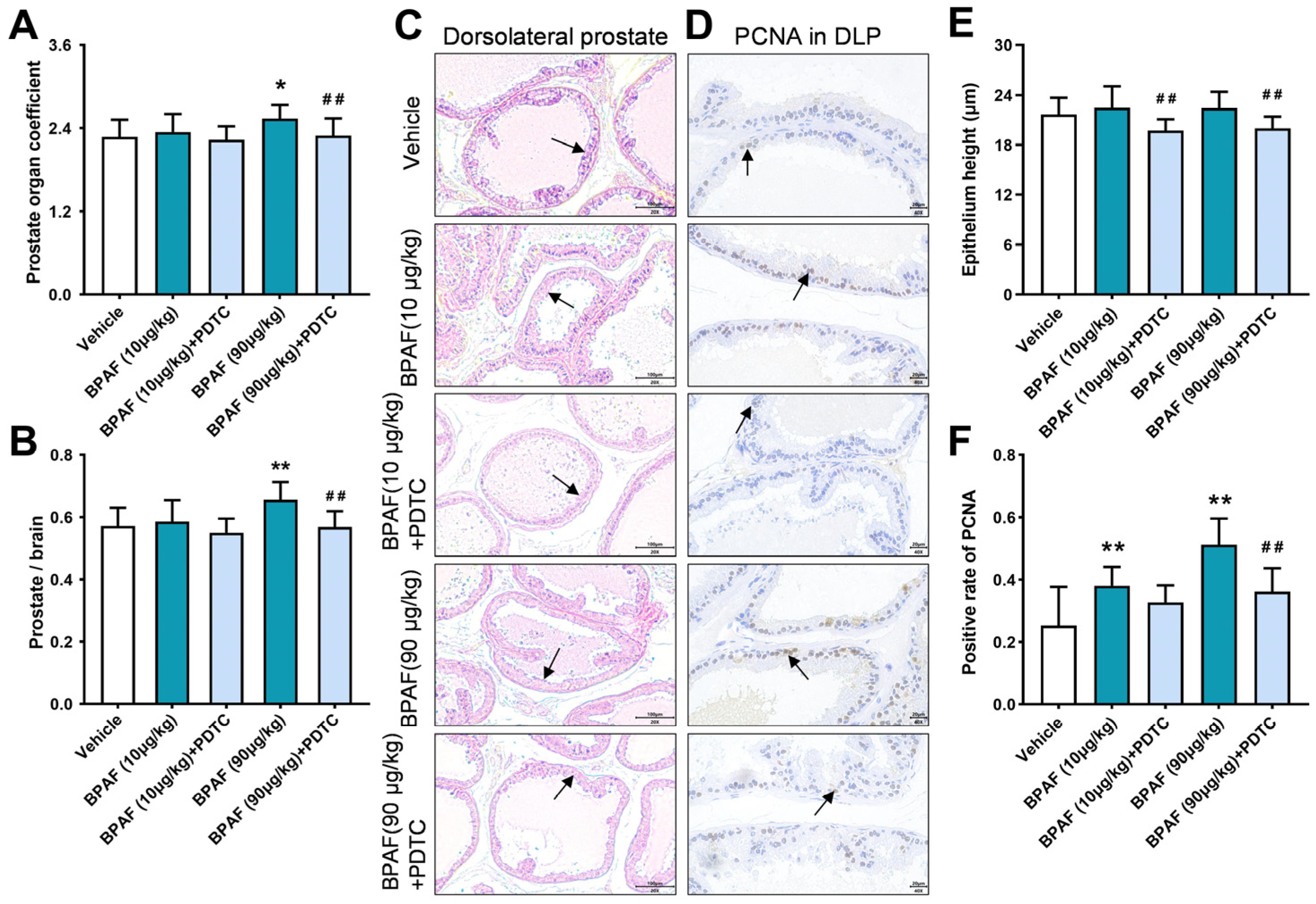
2.1. Anatomical Analysis of Prostates in Rats in Response to BPAF and PDTC
2.2. Histopathological Analysis of Prostates in Rats Induced by BPAF
2.3. Analysis of PCNA Expression in Rat Prostate Tissue Induced by BPAF
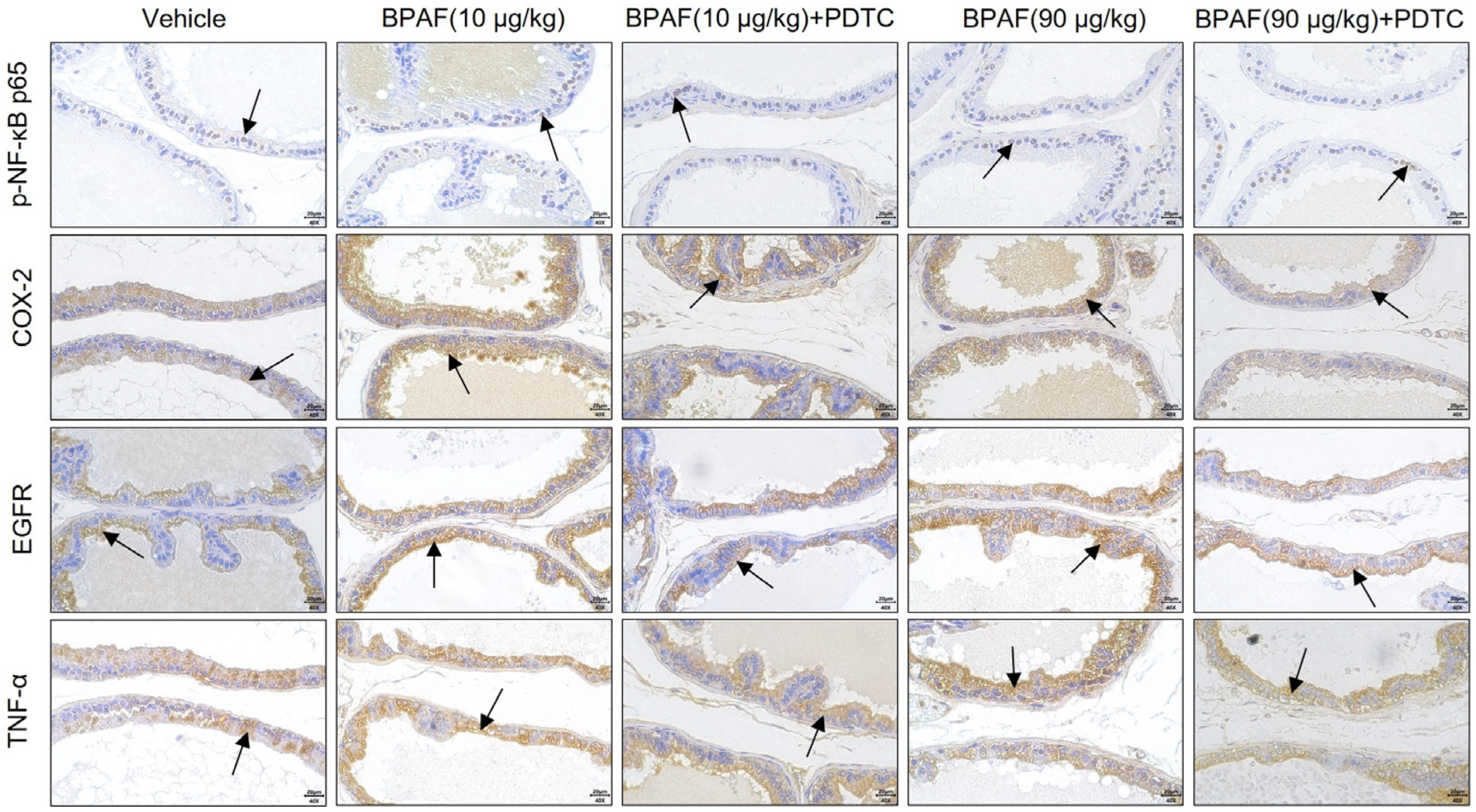
2.4. Localization and Qualification of Key Regulators of NF-κB Signaling Pathway
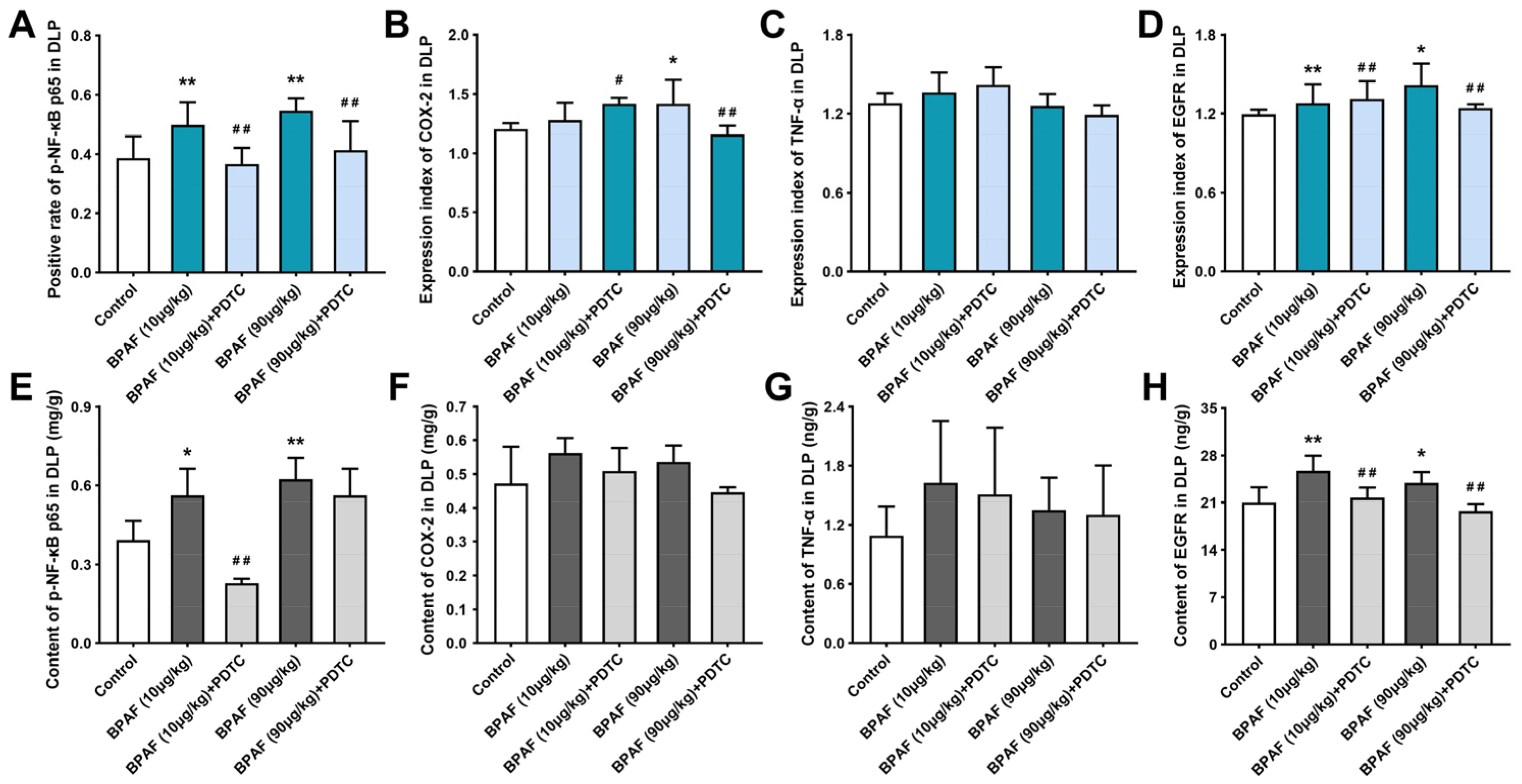
2.5. BPAF Stimulated the Expression of NF-κB p65, COX-2, TNF-α, and EGFR
3. Discussion
4. Materials and Methods
4.1. Animal Treatment
4.2. Histopathological Assessment
4.3. IHC Analysis
4.4. Tissue-Based ELISA
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yilmaz, B.; Terekeci, H.; Sandal, S.; Kelestimur, F. Endocrine disrupting chemicals: Exposure, effects on human health, mechanism of action, models for testing and strategies for prevention. Rev. Endocr. Metab. Disord. 2020, 21, 127–147. [Google Scholar] [CrossRef]
- Zoeller, R.T.; Brown, T.R.; Doan, L.L.; Gore, A.C.; Skakkebaek, N.E.; Soto, A.M.; Woodruff, T.J.; Vom Saal, F.S. Endocrine-disrupting chemicals and public health protection: A statement of principles from The Endocrine Society. Endocrinology 2012, 153, 4097–4110. [Google Scholar] [CrossRef] [PubMed]
- Gore, A.C.; Chappell, V.A.; Fenton, S.E.; Flaws, J.A.; Nadal, A.; Prins, G.S.; Toppari, J.; Zoeller, R.T. EDC-2: The Endocrine Society’s Second Scientific Statement on Endocrine-Disrupting Chemicals. Endocr. Rev. 2015, 36, E1–E150. [Google Scholar] [CrossRef]
- Moosa, A.; Shu, H.; Sarachana, T.; Hu, V.W. Are endocrine disrupting compounds environmental risk factors for autism spectrum disorder? Horm. Behav. 2018, 101, 13–21. [Google Scholar] [CrossRef]
- Rao, A.; Douglas, S.C.; Hall, J.M. Endocrine Disrupting Chemicals, Hormone Receptors, and Acne Vulgaris: A Connecting Hypothesis. Cells 2021, 10, 1439. [Google Scholar] [CrossRef]
- Sifakis, S.; Androutsopoulos, V.P.; Tsatsakis, A.M.; Spandidos, D.A. Human exposure to endocrine disrupting chemicals: Effects on the male and female reproductive systems. Environ. Toxicol. Pharmacol. 2017, 51, 56–70. [Google Scholar] [CrossRef]
- Di Donato, M.; Cernera, G.; Giovannelli, P.; Galasso, G.; Bilancio, A.; Migliaccio, A.; Castoria, G. Recent advances on bisphenol-A and endocrine disruptor effects on human prostate cancer. Mol. Cell Endocrinol. 2017, 457, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Cannarella, R.; Condorelli, R.A.; Barbagallo, F.; La Vignera, S.; Calogero, A.E. Endocrinology of the Aging Prostate: Current Concepts. Front. Endocrinol. 2021, 12, 554078. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Huang, C.J.; Jiao, X.F.; Ding, Z.M.; Zhang, S.X.; Miao, Y.L.; Huo, L.J. Bisphenol AF compromises blood-testis barrier integrity and sperm quality in mice. Chemosphere 2019, 237, 124410. [Google Scholar] [CrossRef] [PubMed]
- Corona, G.; Vignozzi, L.; Rastrelli, G.; Lotti, F.; Cipriani, S.; Maggi, M. Benign prostatic hyperplasia: A new metabolic disease of the aging male and its correlation with sexual dysfunctions. Int. J. Endocrinol. 2014, 2014, 329456. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Huang, D.; Zhou, P.; Su, X.; Yang, R.; Shao, C.; Ma, A.; Wu, J. Individual and Combined Effect of Bisphenol A and Bisphenol AF on Prostate Cell Proliferation through NF-kappaB Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 12283. [Google Scholar] [CrossRef]
- Song, S.; Ruan, T.; Wang, T.; Liu, R.; Jiang, G. Distribution and preliminary exposure assessment of bisphenol AF (BPAF) in various environmental matrices around a manufacturing plant in China. Environ. Sci. Technol. 2012, 46, 13136–13143. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Xin, X.; Ma, F.; Li, X.; Wang, Y.; Zhu, Q.; Chen, H.; Li, H.; Ge, R.S. Bisphenol AF blocks Leydig cell regeneration from stem cells in male rats. Environ. Pollut. 2022, 298, 118825. [Google Scholar] [CrossRef]
- Feng, Y.; Yin, J.; Jiao, Z.; Shi, J.; Li, M.; Shao, B. Bisphenol AF may cause testosterone reduction by directly affecting testis function in adult male rats. Toxicol. Lett. 2012, 211, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Moreman, J.; Lee, O.; Trznadel, M.; David, A.; Kudoh, T.; Tyler, C.R. Acute Toxicity, Teratogenic, and Estrogenic Effects of Bisphenol A and Its Alternative Replacements Bisphenol S, Bisphenol F, and Bisphenol AF in Zebrafish Embryo-Larvae. Environ. Sci. Technol. 2017, 51, 12796–12805. [Google Scholar] [CrossRef]
- Li, Y.; Burns, K.A.; Arao, Y.; Luh, C.J.; Korach, K.S. Differential estrogenic actions of endocrine-disrupting chemicals bisphenol A, bisphenol AF, and zearalenone through estrogen receptor alpha and beta in vitro. Environ. Health Perspect. 2012, 120, 1029–1035. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-kappaB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef]
- Lalle, G.; Twardowski, J.; Grinberg-Bleyer, Y. NF-kappaB in Cancer Immunity: Friend or Foe? Cells 2021, 10, 355. [Google Scholar] [CrossRef]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-kappaB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, S.; Saghari, S.; Bassiri, F.; Raesi, R.; Zarrabi, A.; Hushmandi, K.; Sethi, G.; Tergaonkar, V. NF-kappaB as a regulator of cancer metastasis and therapy response: A focus on epithelial-mesenchymal transition. J. Cell Physiol. 2022, 237, 2770–2795. [Google Scholar] [CrossRef]
- Pande, V.; Sharma, R.K.; Inoue, J.; Otsuka, M.; Ramos, M.J. A molecular modeling study of inhibitors of nuclear factor kappa-B (p50)—DNA binding. J. Comput. Aided Mol. Des. 2003, 17, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Li, R.; Chen, X.; Xue, Y.; Liu, D. Neougonin A Inhibits Lipopolysaccharide-Induced Inflammatory Responses via Downregulation of the NF-kB Signaling Pathway in RAW 264.7 Macrophages. Inflammation 2016, 39, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Charalambous, M.P.; Lightfoot, T.; Speirs, V.; Horgan, K.; Gooderham, N.J. Expression of COX-2, NF-kappaB-p65, NF-kappaB-p50 and IKKalpha in malignant and adjacent normal human colorectal tissue. Br. J. Cancer 2009, 101, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Matusan-Ilijas, K.; Damante, G.; Fabbro, D.; Dordevic, G.; Hadzisejdic, I.; Grahovac, M.; Avirovic, M.; Grahovac, B.; Jonjic, N.; Lucin, K. EGFR expression is linked to osteopontin and Nf-kappaB signaling in clear cell renal cell carcinoma. Clin. Transl. Oncol. 2013, 15, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Mustieles, V.; D’Cruz, S.C.; Couderq, S.; Rodriguez-Carrillo, A.; Fini, J.B.; Hofer, T.; Steffensen, I.L.; Dirven, H.; Barouki, R.; Olea, N.; et al. Bisphenol A and its analogues: A comprehensive review to identify and prioritize effect biomarkers for human biomonitoring. Environ. Int. 2020, 144, 105811. [Google Scholar] [CrossRef]
- Abrantes-Soares, F.; Lorigo, M.; Cairrao, E. Effects of BPA substitutes on the prenatal and cardiovascular systems. Crit. Rev. Toxicol. 2022, 52, 469–498. [Google Scholar] [CrossRef]
- Zeng, J.Y.; Chen, P.P.; Liu, C.; Deng, Y.L.; Miao, Y.; Zhang, M.; Cui, F.P.; Lu, T.T.; Shi, T.; Yang, K.D.; et al. Bisphenol A analogues in associations with serum hormone levels among reproductive-aged Chinese men. Environ. Int. 2022, 167, 107446. [Google Scholar] [CrossRef]
- Nguyen, J.L.; Ricke, E.A.; Liu, T.T.; Gerona, R.; MacGillivray, L.; Wang, Z.; Timms, B.G.; Bjorling, D.E.; Vom Saal, F.S.; Ricke, W.A. Bisphenol-A analogs induce lower urinary tract dysfunction in male mice. Biochem. Pharmacol. 2022, 197, 114889. [Google Scholar] [CrossRef]
- Cho, Y.J.; Park, S.B.; Park, J.W.; Oh, S.R.; Han, M. Bisphenol A modulates inflammation and proliferation pathway in human endometrial stromal cells by inducing oxidative stress. Reprod. Toxicol. 2018, 81, 41–49. [Google Scholar] [CrossRef]
- Song, H.; Park, J.; Bui, P.T.C.; Choi, K.; Gye, M.C.; Hong, Y.C.; Kim, J.H.; Lee, Y.J. Bisphenol A induces COX-2 through the mitogen-activated protein kinase pathway and is associated with levels of inflammation-related markers in elderly populations. Environ. Res. 2017, 158, 490–498. [Google Scholar] [CrossRef]
- Leonel, E.C.R.; Campos, S.G.P.; Bedolo, C.M.B.; Guerra, L.H.A.; Vilamaior, P.S.L.; Calmon, M.F.; Rahal, P.; Amorim, C.A.; Taboga, S.R. Perinatal exposure to bisphenol A impacts in the mammary gland morphology of adult Mongolian gerbils. Exp. Mol. Pathol. 2020, 113, 104374. [Google Scholar] [CrossRef]
- Xiong, S.; Wang, Y.; Li, H.; Zhang, X. Low Dose of Bisphenol A Activates NF-kappaB/IL-6 Signals to Increase Malignancy of Neuroblastoma Cells. Cell. Mol. Neurobiol. 2017, 37, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.F.; Zhang, J.; Shuai, H.L.; Guan, B.Z.; Luo, X.; Yan, R.L. IKKbeta/NF-kappaB mediated the low doses of bisphenol A induced migration of cervical cancer cells. Arch. Biochem. Biophys. 2015, 573, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, U.; Samanta, A.; Biswas, S.; Das, S.; Ghosh, S.; Mandal, D.K.; Maitra, S. Bisphenol A-induced oxidative stress, hepatotoxicity and altered estrogen receptor expression in Labeo bata: Impact on metabolic homeostasis and inflammatory response. Ecotoxicol. Environ. Saf. 2020, 202, 110944. [Google Scholar] [CrossRef] [PubMed]
- Zang, L.; Tian, F.; Yao, Y.; Chen, Y.; Shen, Y.; Han, M.; Meng, Z.; Fan, S.; Zhang, X.; Cai, T.; et al. Qianliexin capsule exerts anti-inflammatory activity in chronic non-bacterial prostatitis and benign prostatic hyperplasia via NF-kappaB and inflammasome. J. Cell. Mol. Med. 2021, 25, 5753–5768. [Google Scholar] [CrossRef]
- Charalambous, M.P.; Maihofner, C.; Bhambra, U.; Lightfoot, T.; Gooderham, N.J.; Colorectal Cancer Study, G. Upregulation of cyclooxygenase-2 is accompanied by increased expression of nuclear factor-kappa B and I kappa B kinase-alpha in human colorectal cancer epithelial cells. Br. J. Cancer 2003, 88, 1598–1604. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, Y.; Li, W.; Zhang, K.; Yuan, Z.; Cai, Y.; Xu, K.; Zhou, J.; Du, Z. Curcuma oil ameliorates benign prostatic hyperplasia through suppression of the nuclear factor-kappa B signaling pathway in rats. J. Ethnopharmacol. 2021, 279, 113703. [Google Scholar] [CrossRef]
- Xie, C.; Sun, X.; Chen, J.; Ng, C.F.; Lau, K.M.; Cai, Z.; Jiang, X.; Chan, H.C. Down-regulated CFTR During Aging Contributes to Benign Prostatic Hyperplasia. J. Cell Physiol. 2015, 230, 1906–1915. [Google Scholar] [CrossRef]
- Chopra, D.P.; Menard, R.E.; Januszewski, J.; Mattingly, R.R. TNF-alpha-mediated apoptosis in normal human prostate epithelial cells and tumor cell lines. Cancer Lett. 2004, 203, 145–154. [Google Scholar] [CrossRef]
- Abdou, H.M.; Abd Elkader, H.A.E.; El-Gendy, A.H.; Eweda, S.M. Neurotoxicity and neuroinflammatory effects of bisphenol A in male rats: The neuroprotective role of grape seed proanthocyanidins. Environ. Sci. Pollut. Res. Int. 2022, 29, 9257–9268. [Google Scholar] [CrossRef]
- Shi, F.; Deng, Z.; Zhou, Z.; Jiang, B.; Jiang, C.Y.; Zhao, R.Z.; Sun, F.; Cui, D.; Sun, M.H.; Sun, Q.; et al. Heat injured stromal cells-derived exosomal EGFR enhances prostatic wound healing after thulium laser resection through EMT and NF-kappaB signaling. Prostate 2019, 79, 1238–1255. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Zhang, Y.; Huang, M.; Deng, Z.; Zhang, A.; Pei, L.; Wang, L.; Zhao, W.; Ma, L.; Zhang, Q.; et al. Urinary exosomes-based Engineered Nanovectors for Homologously Targeted Chemo-Chemodynamic Prostate Cancer Therapy via abrogating EGFR/AKT/NF-kB/IkB signaling. Biomaterials 2021, 275, 120946. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, D.; Su, X.; Yan, H.; Wu, J.; Sun, Z. Oral exposure to low-dose bisphenol A induces hyperplasia of dorsolateral prostate and upregulates EGFR expression in adult Sprague-Dawley rats. Toxicol. Ind. Health 2019, 35, 647–659. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Group | Anatomical | Histological | IHC | ELISA |
|---|---|---|---|---|
| BPAF (10 μg/kg) | - | - | PCNA ↑, NF-κB p65 ↑, EGFR ↑ | NF-κB p65 ↑, EGFR ↑ |
| BPAF (10 μg/kg) + PDTC | - | Epithelial height of DLP ↓ | NF-κB p65 ↓, COX-2 ↓, EGFR ↓ | NF-κB p65 ↓, EGFR ↓ |
| BPAF (90 μg/kg) | Prostate organ coefficient ↑a, prostate–brain coefficient ↑ | - | PCNA ↑, NF-κB p65 ↑, COX-2 ↑, EGFR ↑ | NF-κB p65 ↑, EGFR ↑ |
| BPAF (90 μg/kg) + PDTC | Prostate organ coefficient ↓b, prostate–brain coefficient ↓ | Epithelial height of DLP ↓ | PCNA ↓, NF-κB p65 ↓, COX-2 ↓, EGFR ↓ | EGFR ↓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.; Wang, K.; Huang, D.; Su, X.; Yang, R.; Shao, C.; Jiang, J.; Wu, J. Bisphenol AF Induces Prostatic Dorsal Lobe Hyperplasia in Rats through Activation of the NF-κB Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 16221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216221
Huang S, Wang K, Huang D, Su X, Yang R, Shao C, Jiang J, Wu J. Bisphenol AF Induces Prostatic Dorsal Lobe Hyperplasia in Rats through Activation of the NF-κB Signaling Pathway. International Journal of Molecular Sciences. 2023; 24(22):16221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216221
Chicago/Turabian StyleHuang, Sisi, Kaiyue Wang, Dongyan Huang, Xin Su, Rongfu Yang, Congcong Shao, Juan Jiang, and Jianhui Wu. 2023. "Bisphenol AF Induces Prostatic Dorsal Lobe Hyperplasia in Rats through Activation of the NF-κB Signaling Pathway" International Journal of Molecular Sciences 24, no. 22: 16221. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms242216221