
The Immunogenicity of DENV1–4 ED3s Strongly Differ despite Their Almost Identical Three-Dimensional Structures and High Sequence Similarities

 , , and
, , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
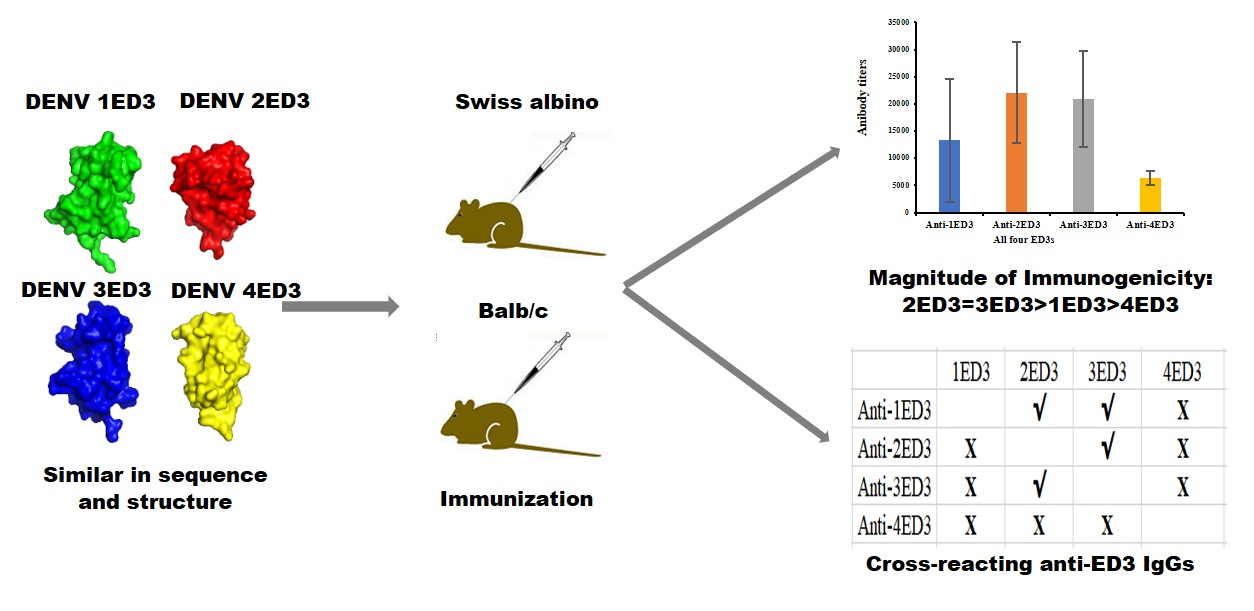
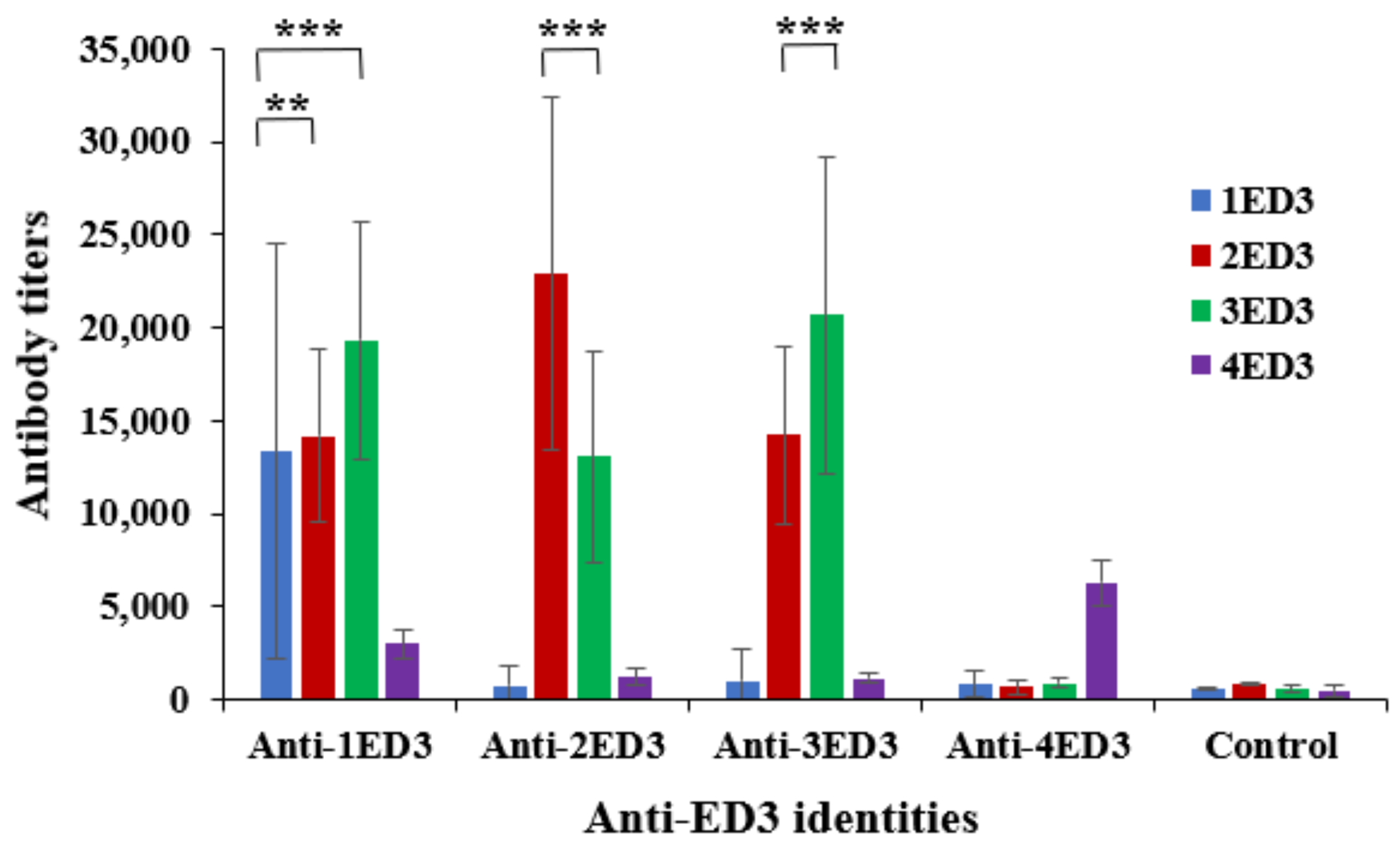
2.1. Serospecificity of Anti-DENV ED3 Sera
2.2. Sero-Cross-Reactivity of Anti-DENV ED3 Sera
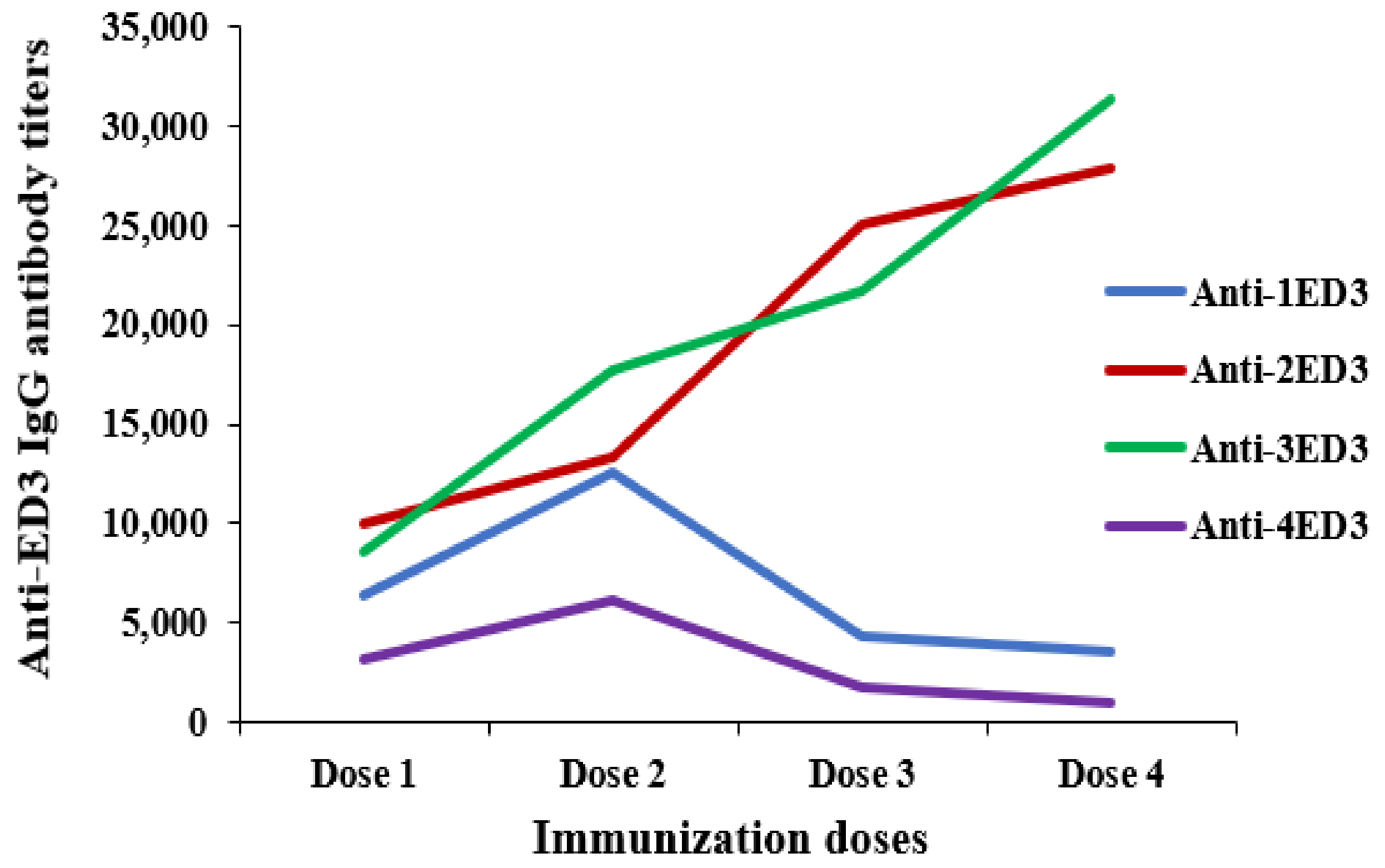
2.3. Long-Term Antibody Response against ED3s
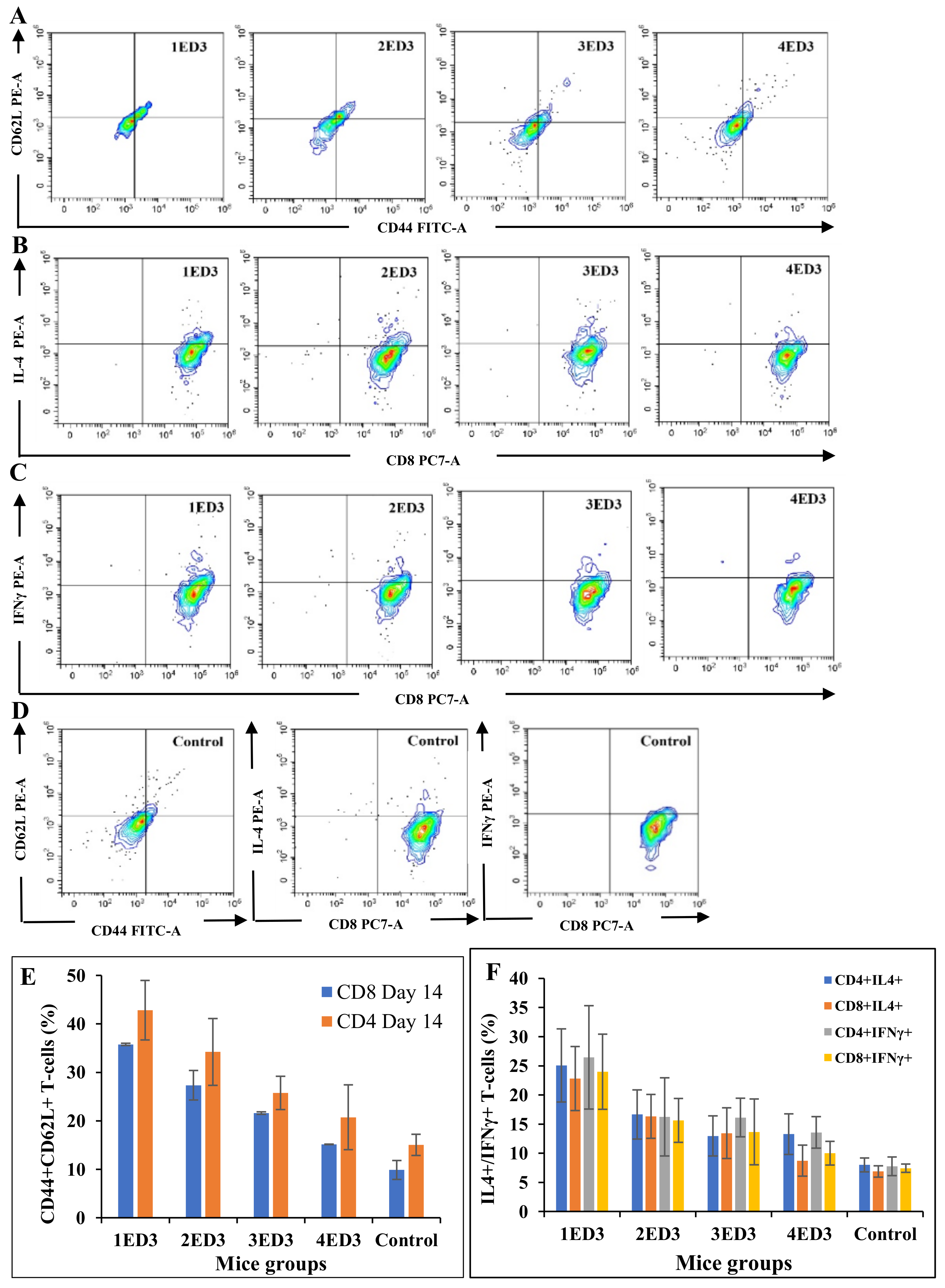
2.4. CD Markers and Cytokines Expression
3. Discussion
4. Materials and Methods
4.1. Mutant Design Protein Expression and Purification
4.2. Analytical Reverse-Phase High-Performance Chromatography (HPLC)
4.3. Matrix-Assisted Laser Desorption/Ionization-Time of Flight Mass Spectroscopy (MALDI-TOF MS)
4.4. Immunization Protocol
4.5. Anti-ED3 IgG Titer by ELISA
4.6. Flow Cytometry Analysis of CD Markers
4.6.1. Cell Surface CD Marker Analysis
4.6.2. Intracellular Cytokine Analysis
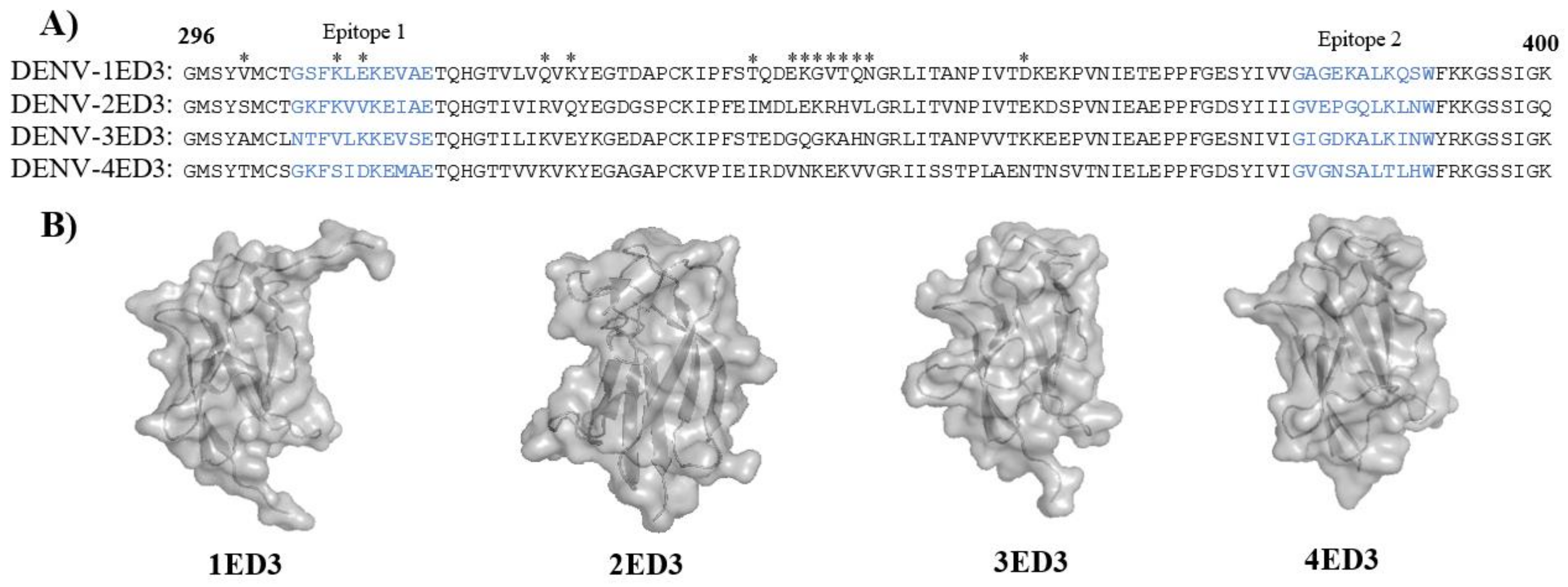
4.7. Structure Modeling of Four Different ED3s
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ashley, E.A. Dengue fever. Trends Anaesth. Crit. 2011, 1, 39–41. [Google Scholar] [CrossRef]
- Farrar, J.; Focks, D.; Gubler, D.; Barrera, R.; Guzman, M.G.; Simmons, C.; Kalayanarooj, S.; Lum, L.; McCall, P.J.; Lloyd, L.; et al. Towards a global dengue research agenda. Trop. Med. Int. Health 2007, 12, 695. [Google Scholar] [CrossRef] [PubMed]
- Aguas, R.; Dorigatti, I.; Coudeville, L.; Luxemburger, C.; Ferguson, N.M. Cross-serotype interactions and disease outcome prediction of dengue infections in Vietnam. Sci. Rep. 2019, 9, 9395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Stanaway, J.D. The global economic burden of dengue: A systematic analysis. Lancet 2016, 16, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Murray, N.E.; Quam, M.B.; Wilder-Smith, A. Epidemiology of dengue: Past, present and future prospects. J. Clin. Epidemiol. 2013, 5, 299. [Google Scholar]
- Rahman, N.; Miura, S.; Okawa, M.; Kibria, M.G.; Islam, M.M.; Kuroda, Y. Solubility Controlling Peptide Tags of Opposite Charges Generate a Bivalent Immune Response Against Dengue ED3 Serotypes 3 and 4. Front. Immunol. 2021, 12, 671590. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [Green Version]
- Warrilow, D.; Northill, J.A.; Pyke, A.; Smith, G.A. Single rapid TaqMan fluorogenic probe-based PCR assay that detects all four dengue serotypes. J. Med. Virol. 2002, 66, 524–528. [Google Scholar] [CrossRef]
- Murphy, B.R.; Whitehead, S.S. Immune response to dengue virus and prospects for a vaccine. Annu. Rev. Immunol. 2011, 29, 587–619. [Google Scholar] [CrossRef]
- Imrie, A.; Meeks, J.; Gurary, A.; Sukhbaatar, M.; Truong, T.T.; Cropp, C.B.; Effler, P. Antibody to dengue 1 detected more than 60 years after infection. Viral Immunol. 2007, 20, 672–675. [Google Scholar] [CrossRef] [Green Version]
- Rothman, A.L. Dengue: Defining protective versus pathologic immunity. J. Clin. Investig. 2004, 113, 946–951. [Google Scholar] [CrossRef]
- Snow, G.E.; Haaland, B.; Ooi, E.E.; Gubler, D.J. Research on dengue during World War II revisited. Am. J. Trop. Med. 2014, 91, 1203. [Google Scholar] [CrossRef] [Green Version]
- Wahala, W.M.; De Silva, A.M. The human antibody response to dengue virus infection. Viruses 2011, 3, 2374–2395. [Google Scholar] [CrossRef] [Green Version]
- Recker, M.; Blyuss, K.B.; Simmons, C.P.; Hien, T.T.; Wills, B.; Farrar, J.; Gupta, S. Immunological serotype interactions and their effect on the epidemiological pattern of dengue. Proc. Royal Soc. B 2009, 276, 2541–2548. [Google Scholar] [CrossRef]
- Halstead, S.B.; Nimmannitya, S.; Cohen, S.N. Observations related to pathogenesis of dengue hemorrhagic fever. IV. Relation of disease severity to antibody response and virus recovered. Yale J. Biol. Med. 1970, 42, 311. [Google Scholar]
- Gebhard, L.G.; Filomatori, C.V.; Gamarnik, A.V. Functional RNA elements in the dengue virus genome. Viruses 2011, 3, 1739–1756. [Google Scholar] [CrossRef]
- Gromowski, G.D.; Barrett, A.D. Characterization of an antigenic site that contains a dominant, type-specific neutralization determinant on the envelope protein domain III (ED3) of dengue 2 virus. Virology 2007, 366, 349–360. [Google Scholar] [CrossRef] [Green Version]
- Slon-Campos, J.L.; Dejnirattisai, W.; Jagger, B.W.; López, C.C.; Wongwiwat, W.; Durnell, L.A.; Winkler, E.S.; Chen, R.E.; Reyes, S.A.; Rey, F.A.; et al. A protective Zika virus E-dimer-based subunit vaccine engineered to abrogate antibody-dependent enhancement of dengue infection. Nat. Immunol. 2019, 20, 1291–1298. [Google Scholar] [CrossRef]
- Rajamanonmani, R.; Nkenfou, C.; Clancy, P.; Yau, Y.H.; Shochat, S.G.; Sukupolvi, P.S.; Schul, W.; Diamond, M.S.; Vasudevan, S.G.; Lescar, J. On a mouse monoclonal antibody that neutralizes all four dengue virus serotypes. J. Gen. Virol. 2009, 90, 799. [Google Scholar] [CrossRef]
- Midgley, C.M.; Bajwa, J.M.; Vasanawathana, S.; Limpitikul, W.; Wills, B.; Flanagan, A.; Waiyaiya, E.; Tran, H.B.; Cowper, A.E.; Chotiyarnwon, P.; et al. An in-depth analysis of original antigenic sin in dengue virus infection. J. Virol. 2011, 85, 410–421. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.M.; Miura, S.; Hasan, M.N.; Rahman, N.; Kuroda, Y. Anti-Dengue ED3 long-term immune response with T-cell memory generated using Solubility Controlling Peptide tags. Front. Immunol. 2020, 11, 333. [Google Scholar] [CrossRef] [PubMed]
- Lazo, L. Dengue virus 4: The ‘black sheep’of the family. Expert Rev. Vaccines 2020, 19, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayak, V.; Dessau, M.; Kucera, K.; Anthony, K.; Ledizet, M.; Modis, Y. Crystal structure of dengue virus type 1 envelope protein in the post fusion conformation and its implications for membrane fusion. J. Virol. 2009, 83, 4338–4344. [Google Scholar] [CrossRef] [Green Version]
- Kikuti, C.; Rouvinski, A.; Guardado, C.P.; Barba, S.P.; Barba, S.G.; Duquerroy, S.; Vaney, M.C.; Rey, F.A. Crystal structure of dengue 2 virus envelope glycoprotein. Nature 2015, 520, 109. [Google Scholar]
- Elahi, M.; Islam, M.M.; Noguchi, K.; Yohda, M.; Kuroda, Y. High resolution crystal structure of dengue-3 envelope protein domain III suggests possible molecular mechanisms for serospecific antibody recognition. Proteins Struct. Funct. Genet. 2013, 81, 1090–1095. [Google Scholar] [CrossRef]
- Elahi, M.; Islam, M.M.; Noguchi, K.; Yohda, M.; Toh, H.; Kuroda, Y. Crystal structure of dengue 4 envelope protein domain III (ED3). Biochim. Biophys. Acta Proteins Proteom. 2014, 1844, 585–592. [Google Scholar] [CrossRef]
- Kulkarni, M.R.; Islam, M.M.; Numoto, N.; Elahi, M.; Mahib, M.R.; Ito, N.; Kuroda, Y. Structural and biophysical analysis of sero-specific immune responses using epitope grafted Dengue ED3 mutants. Biochim. Biophys. Acta Proteins Proteom. 2015, 1854, 1438–1443. [Google Scholar] [CrossRef]
- Lovell, S.C.; Word, J.M.; Richardson, J.S.; Richardson, D.C. The penultimate rotamer library. Proteins Struct. Funct. Genet. 2000, 40, 389–408. [Google Scholar] [CrossRef]
- Ji, G.H.; Deng, Y.Q.; Yu, X.J.; Jiang, T.; Wang, H.J.; Shi, X.; Zhang, D.P.; Li, X.F.; Zhu, S.Y.; Zhao, H.; et al. Characterization of a novel dengue serotype 4 virus-specific neutralizing epitope on the envelope protein domain III. PLoS ONE 2015, 10, e0139741. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Cheng, G. Vaccines and immunization strategies for dengue prevention. Emerg. Microbes Infect. 2016, 5, e77. [Google Scholar] [CrossRef] [Green Version]
- Kaech, S.M.; Wherry, E.J.; Ahmed, R. Effector and memory T-cell differentiation: Implications for vaccine development. Nat. Rev. Immunol. 2002, 2, 251–262. [Google Scholar] [CrossRef]
- Bannard, O.; Kraman, M.; Fearon, D. Pathways of memory CD8+ T-cell development. Eur. J. Immunol. 2009, 39, 2083–2087. [Google Scholar] [CrossRef]
- Roberts, A.D.; Ely, K.H.; Woodland, D.L. Differential contributions of central and effector memory T cells to recall responses. J. Exp. Med. 2005, 202, 123–133. [Google Scholar] [CrossRef]
- Faassen, V.H.; Saldanha, M.; Gilbertson, D.; Dudani, R.; Krishnan, L.; Sad, S. Reducing the stimulation of CD8+ T cells during infection with intracellular bacteria promotes differentiation primarily into a central (CD62LhighCD44high) subset. J. Immunol. 2014, 174, 5341–5350. [Google Scholar] [CrossRef] [Green Version]
- Baaten, B.J.; Tinoco, R.; Chen, A.T.; Bradley, L.M. Regulation of antigen-experienced T cells: Lessons from the quintessential memory marker CD44. Front. Immunol. 2012, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Budd, R.C.; Cerottini, J.C.; Horvath, C.; Bron, C.; Pedrazzini, T.; Howe, R.C.; MacDonald, H.R. Distinction of virgin and memory T lymphocytes. Stable acquisition of the Pgp-1 glycoprotein concomitant with antigenic stimulation. J. Immunol. 1987, 138, 3120–3129. [Google Scholar] [CrossRef]
- Liang, X.; Li, X.; Duan, J.; Chen, Y.; Wang, X.; Pang, L.; Kong, D.; Song, B.; Li, C.; Yang, J. Nanoparticles with CD44 targeting and ROS triggering properties as effective in vivo antigen delivery system. Mol. Pharm. 2018, 15, 508–518. [Google Scholar] [CrossRef]
- Feng, C.G.; Bean, A.G.; Hooi, H.; Briscoe, H.; Britton, W.J. Increase in gamma interferon-secreting CD8+, as well as CD4+, T cells in lungs following aerosol infection with Mycobacterium tuberculosis. Infect. Immun. 1999, 67, 3242–3247. [Google Scholar] [CrossRef] [Green Version]
- Whitmire, J.K.; Asano, M.S.; Murali, K.K.; Suresh, M.; Ahmed, R. Long-term CD4 Th1 and Th2 memory following acute lymphocytic choriomeningitis virus infection. J. Virol. 1998, 72, 8281–8288. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, M.; Bryson, C.J.; Cloake, E.A.; Welch, K.; Filipe, V.; Romeijn, S.; Hawe, A.; Jiskoot, W.; Baker, M.P.; Fogg, M.H. Small amounts of sub-visible aggregates enhance the immunogenic potential of monoclonal antibody therapeutics. Pharm. Res. 2015, 32, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Maki, K.; Ebina, T.; Kuwajima, K.; Soda, K.; Kuroda, Y. Mutational analysis of protein solubility enhancement using short peptide tags. Biopolymers 2007, 85, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Kibria, M.G.; Akazawa, O.Y.; Rahman, N.; Hagihara, Y.; Kuroda, Y. The immunogenicity of an anti-EGFR single domain antibody (VHH) is enhanced by misfolded amorphous aggregation but not by heat-induced aggregation. Eur. J. Pharm. 2020, 152, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Frei, J.C.; Wirchnianski, A.S.; Govero, J.; Vergnolle, O.; Dowd, K.A.; Pierson, T.C.; Kielian, M.; Girvin, M.E.; Diamond, M.S.; Lai, J.R. Engineered dengue virus domain III proteins elicit cross-neutralizing antibody responses in mice. J. Virol. 2018, 92, e01023-18. [Google Scholar] [CrossRef] [Green Version]
- Brindha, S.; Kuroda, Y. A Multi-Disulfide Receptor-Binding Domain (RBD) of the SARS-CoV-2 Spike Protein Expressed in E. coli Using a SEP-Tag Produces Antisera Interacting with the Mammalian Cell Expressed Spike (S1) Protein. Int. J. Mol. Sci. 2022, 23, 1703. [Google Scholar] [CrossRef]
- Saotome, T.; Yamazaki, T.; Kuroda, Y. Misfolding of a single disulfide bonded globular protein into a low-solubility species conformationally and biophysically distinct from the native one. Biomolecules 2019, 9, 250. [Google Scholar] [CrossRef] [Green Version]
- Babu, J.P.; Pattnaik, P.; Gupta, N.; Shrivastava, A.; Khan, M.; Rao, P.L. Immunogenicity of a recombinant envelope domain III protein of dengue virus type-4 with various adjuvants in mice. Vaccine 2008, 26, 4655–4663. [Google Scholar] [CrossRef]
- Kibria, M.G.; Akazawa-Ogawa, Y.; Hagihara, Y.; Kuroda, Y. Immune response with long-term memory triggered by amorphous aggregates of misfolded anti-EGFR VHH-7D12 is directed against the native VHH-7D12 as well as the framework of the analogous VHH-9G8. Eur. J. Pharm. 2021, 165, 13–21. [Google Scholar]
- Rahman, N.; Islam, M.M.; Kibria, M.G.; Unzai, S.; Kuroda, Y. A systematic mutational analysis identifies a 5-residue proline tag that enhances the in vivo immunogenicity of a non-immunogenic model protein. FEBS Open Bio 2020, 10, 1947–1956. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.D.; Sharmin, T.; Tipo, I.H.; Saha, A.; Yesmin, S.; Roy, M.G.; Brindha, S.; Kuroda, Y.; Islam, M.M. The Immunogenicity of DENV1–4 ED3s Strongly Differ despite Their Almost Identical Three-Dimensional Structures and High Sequence Similarities. Int. J. Mol. Sci. 2023, 24, 2393. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032393
Islam MD, Sharmin T, Tipo IH, Saha A, Yesmin S, Roy MG, Brindha S, Kuroda Y, Islam MM. The Immunogenicity of DENV1–4 ED3s Strongly Differ despite Their Almost Identical Three-Dimensional Structures and High Sequence Similarities. International Journal of Molecular Sciences. 2023; 24(3):2393. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032393
Chicago/Turabian StyleIslam, Md. Din, Tahmina Sharmin, Imrul Hasan Tipo, Antara Saha, Sanjida Yesmin, Moushumi Ghosh Roy, Subbaian Brindha, Yutaka Kuroda, and M. Monirul Islam. 2023. "The Immunogenicity of DENV1–4 ED3s Strongly Differ despite Their Almost Identical Three-Dimensional Structures and High Sequence Similarities" International Journal of Molecular Sciences 24, no. 3: 2393. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032393