
Influence of PEG Chain Length of Functionalized Magnetic Nanoparticles on the Cytocompatibility and Immune Competence of Primary Murine Macrophages and Dendritic Cells
 , , and
, , and
Abstract
:1. Introduction
2. Results
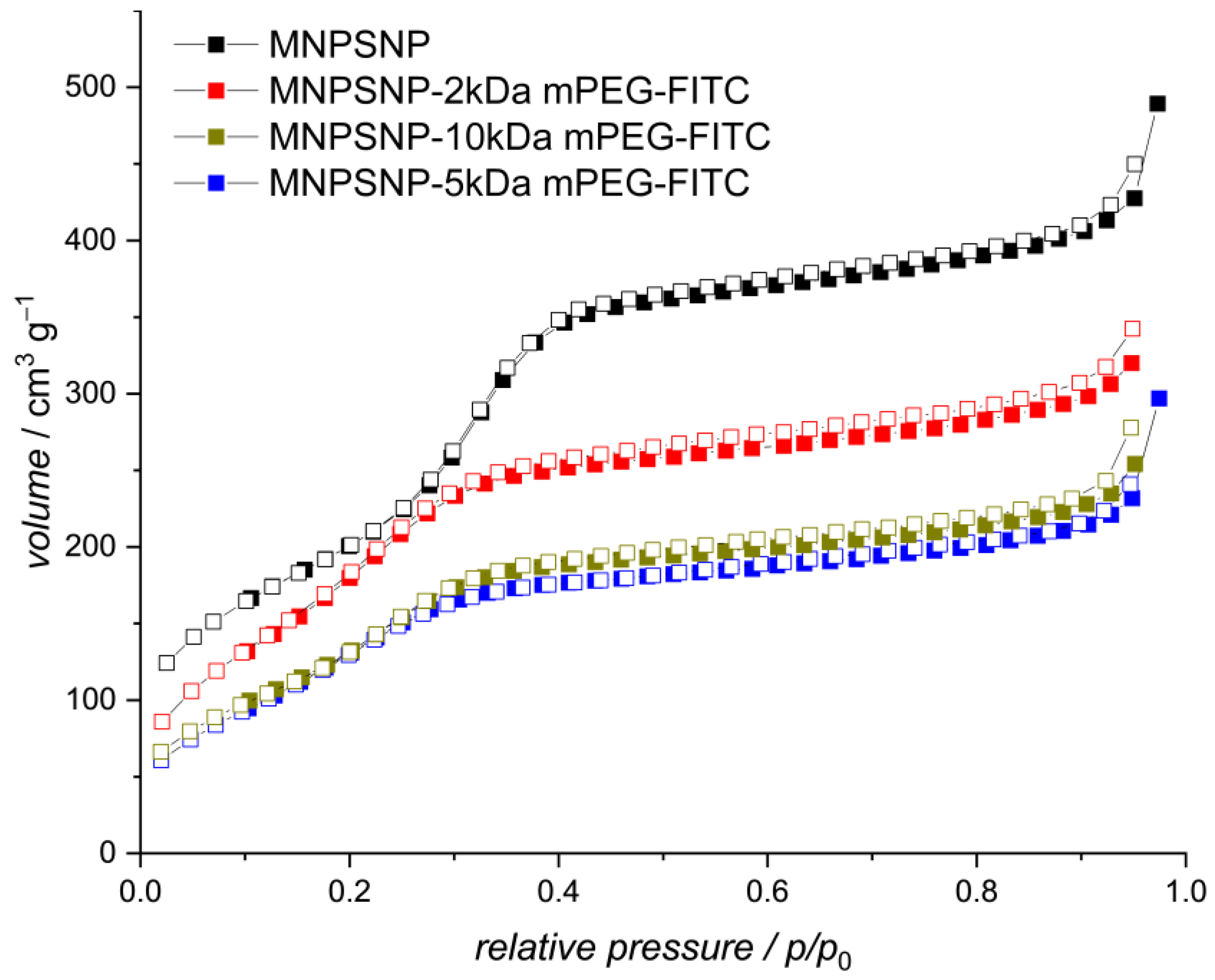
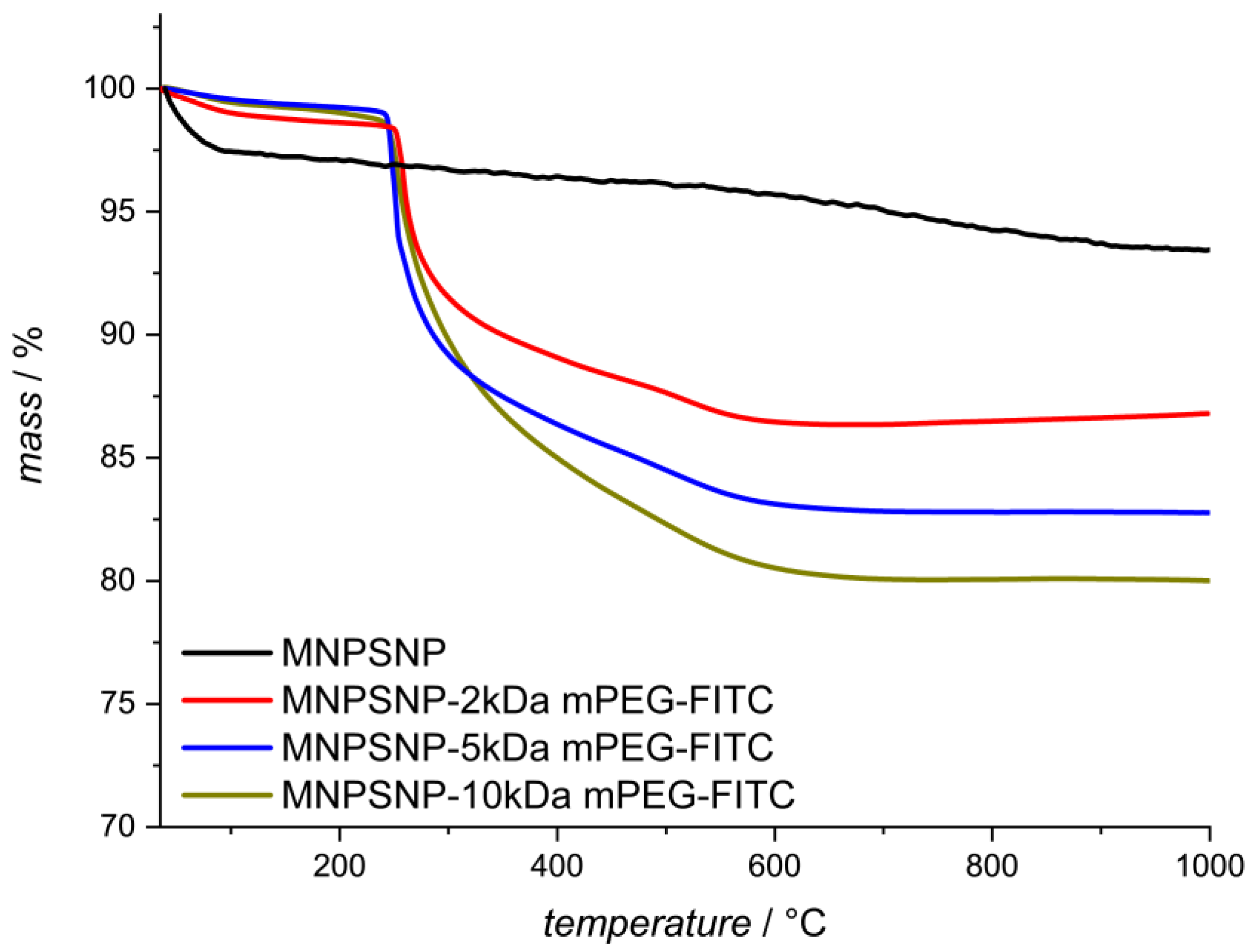
2.1. Characterization of Unfunctionalized and Functionalized MNPSNPs
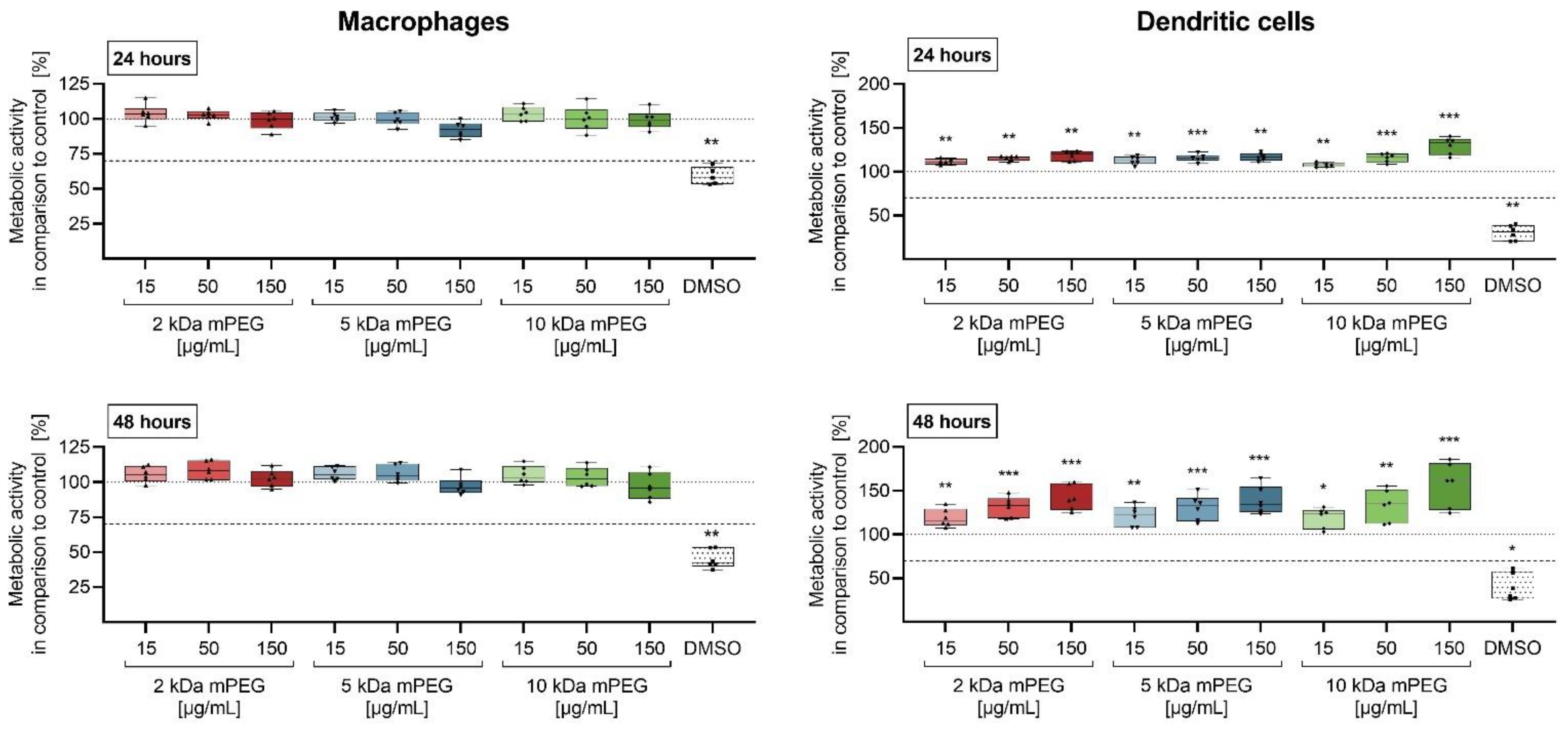
2.2. Cytotoxicity of Functionalized MNPSNPs
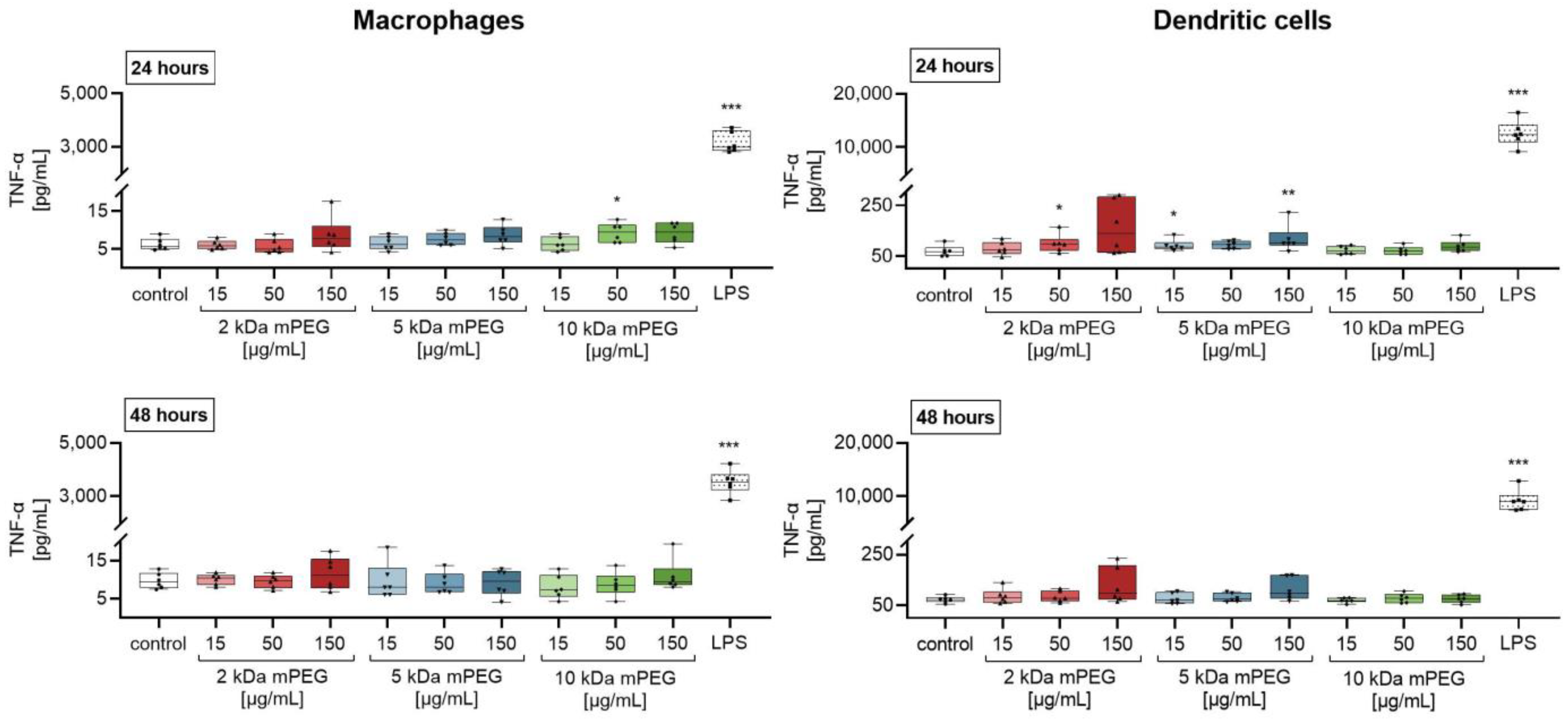
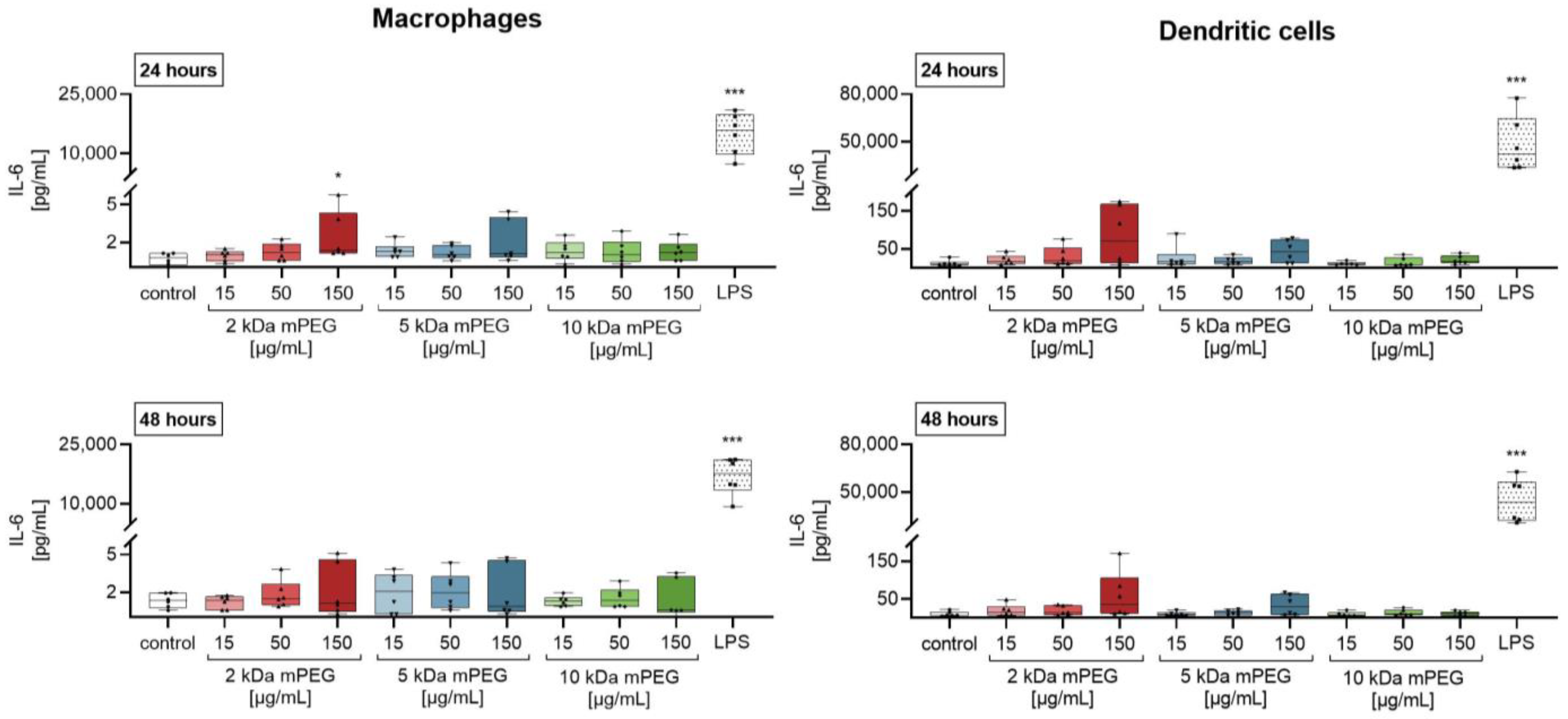
2.3. Effect of Functionalized MNPSNPs on Cytokine Release
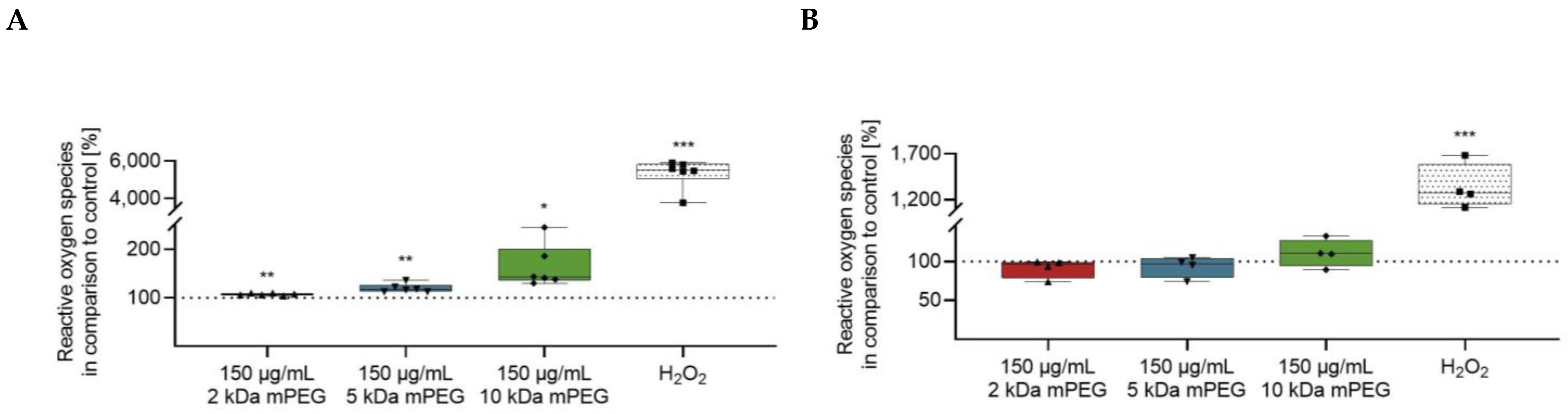
2.4. Effect of Functionalized MNPSNPs on the Formation of Reactive Oxygen Species
2.5. Impact of Functionalized MNPSNPs on the Cell Migration of DCs
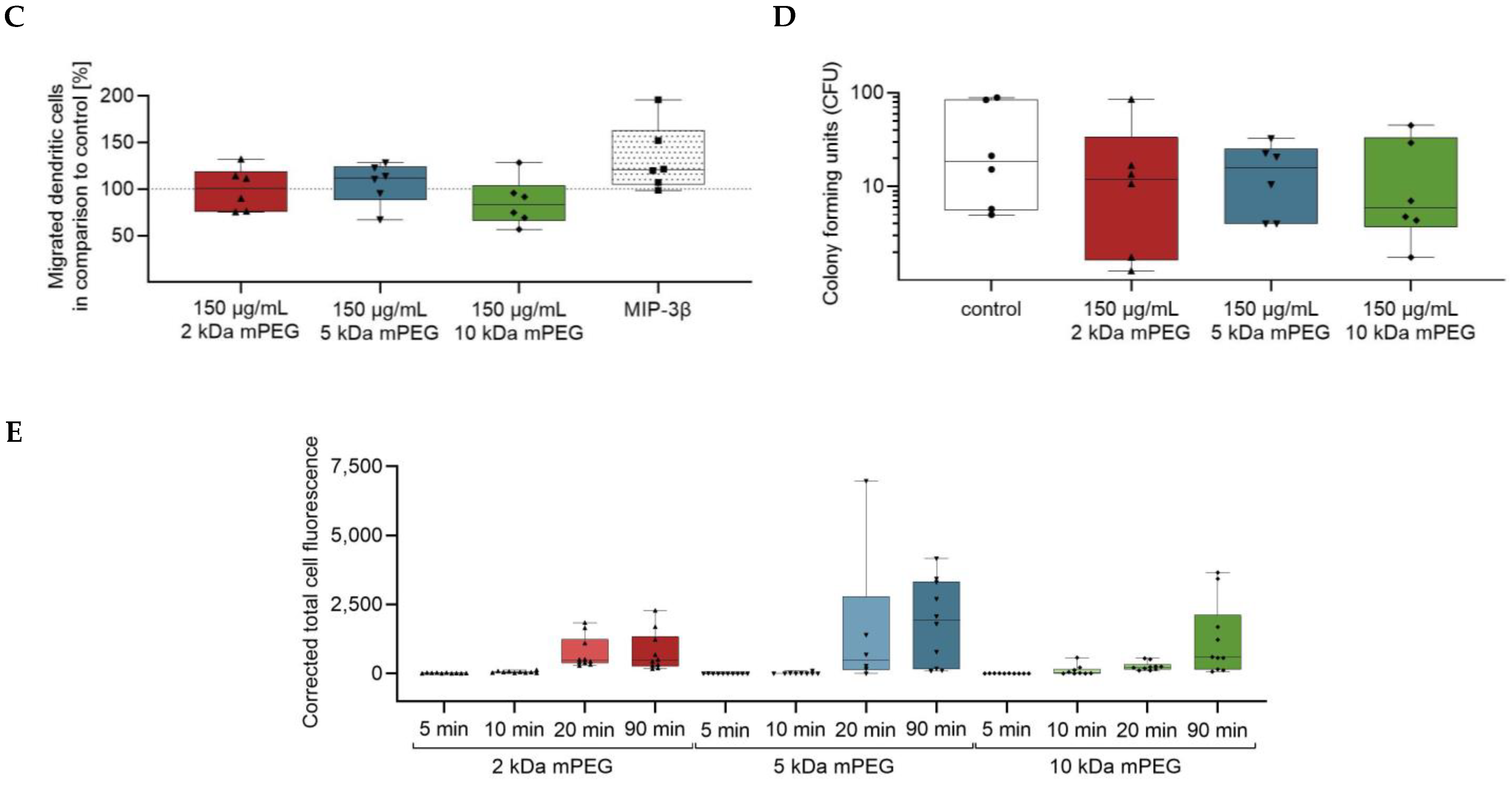
2.6. Impact of Functionalized MNPSNPs on the Immune Competence of Macrophages
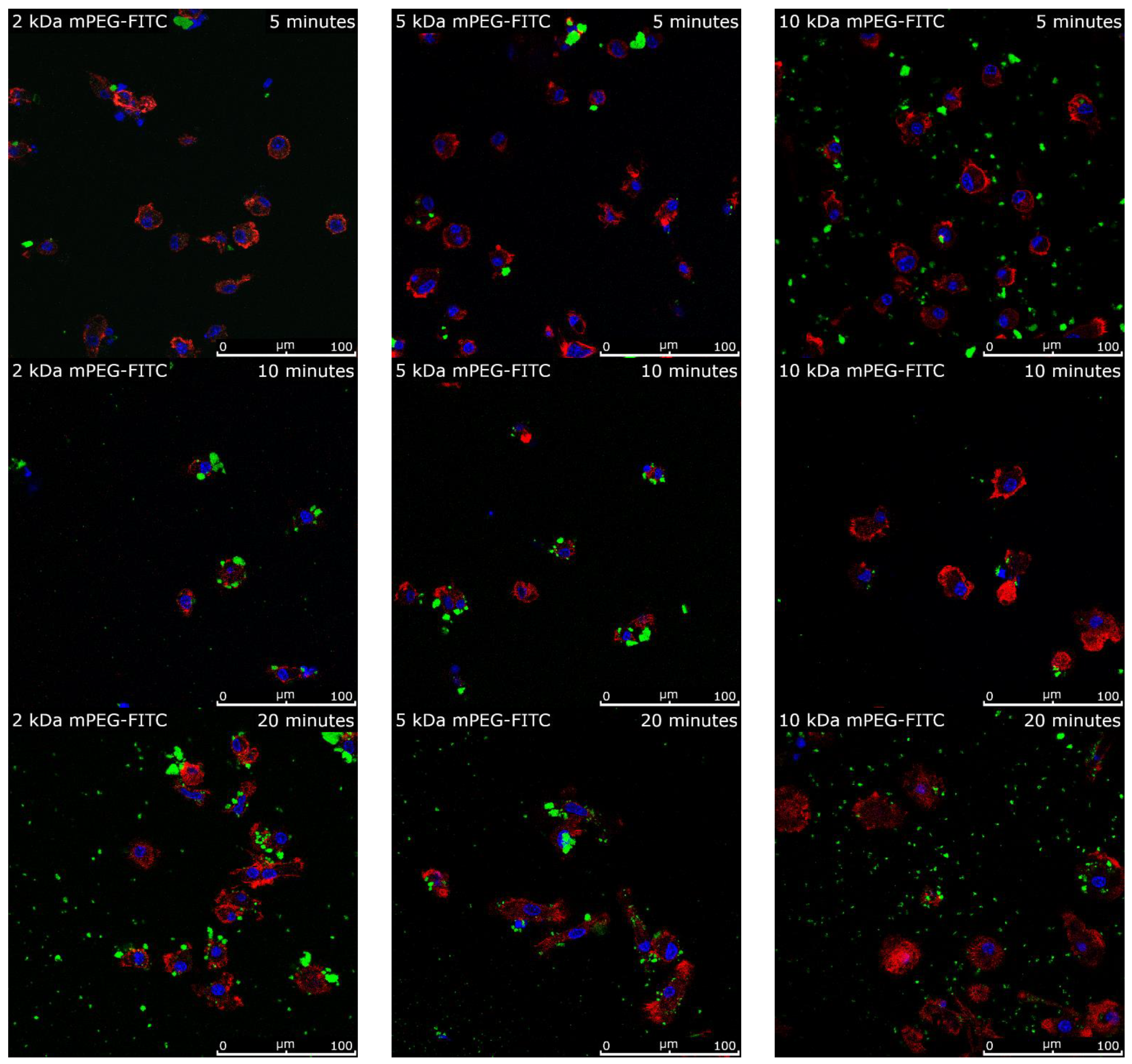
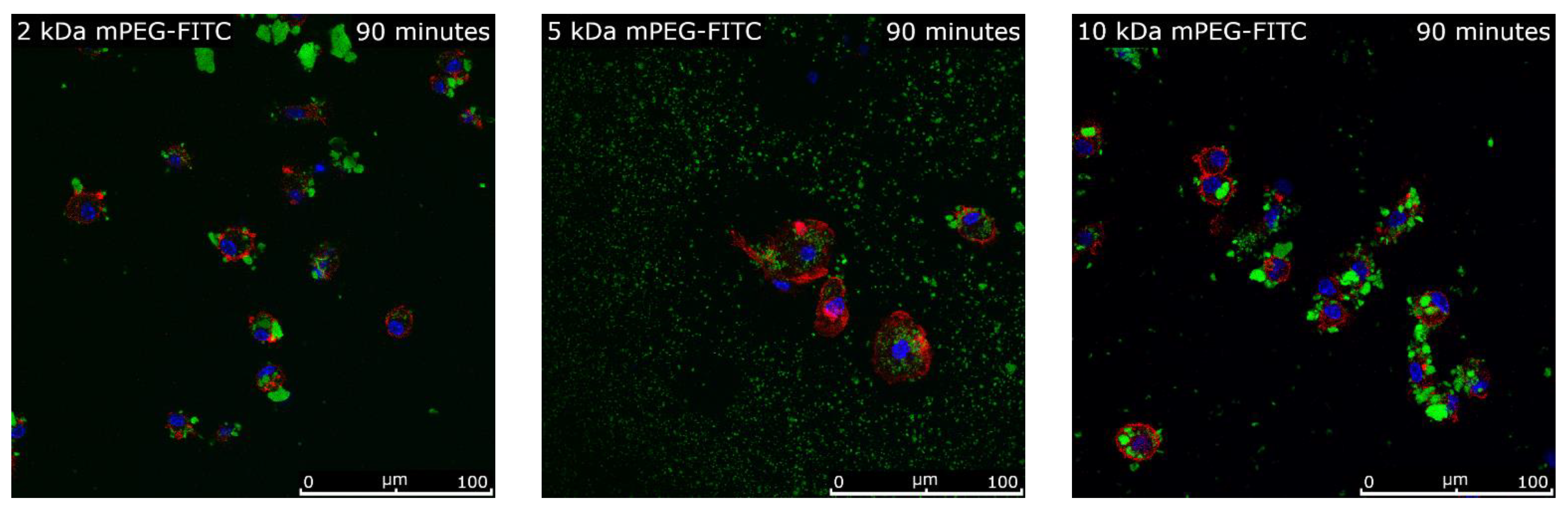
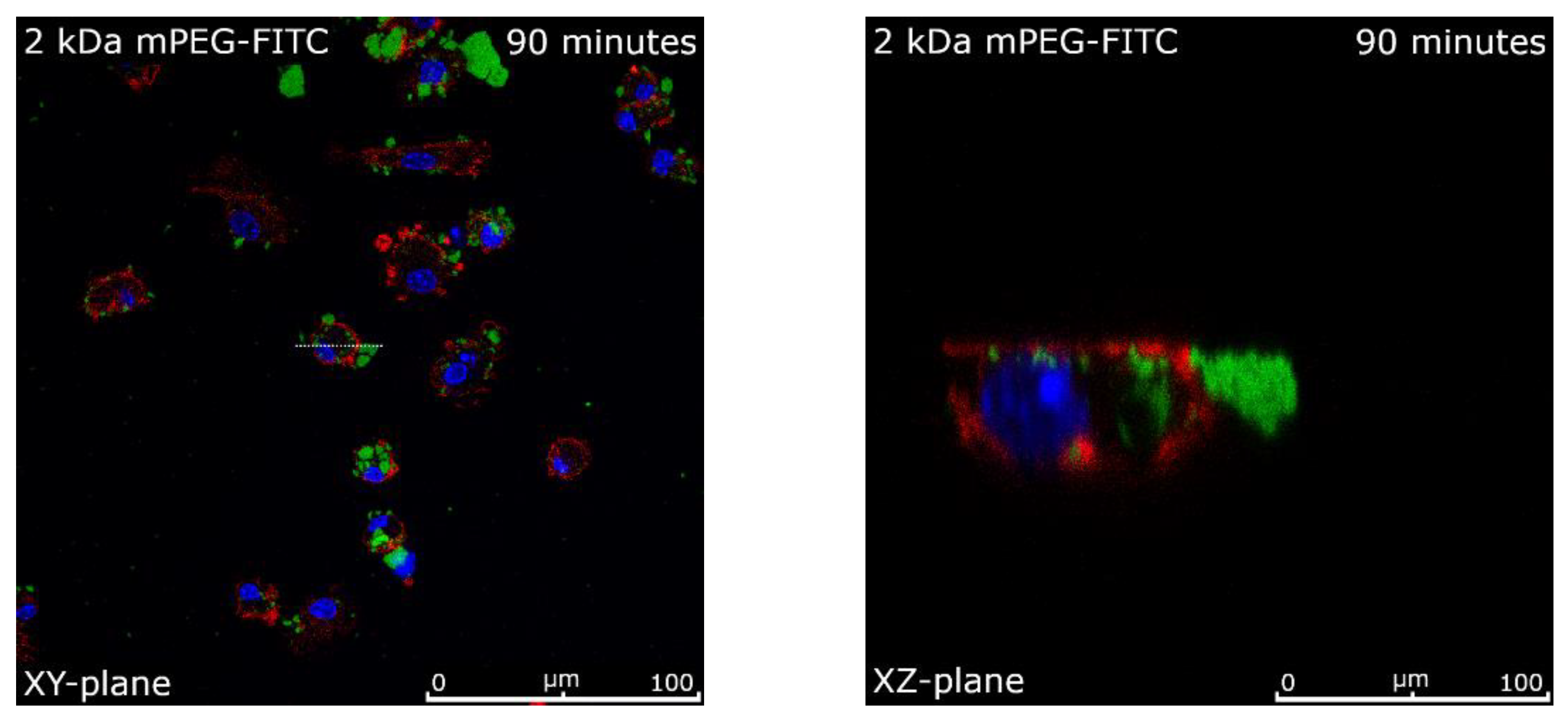
2.7. Intracellular Uptake of Functionalized MNPSNPs
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Synthesis of Magnetic Nanoporous Silica Nanoparticles (MNPSNPs) and Functionalization with mPEG
4.2.1. Synthesis of MNPSNPs
4.2.2. Synthesis of mPEG Silanes with Different Molecular Weights
4.2.3. Functionalization of MNPSNPs with mPEG and FITC
4.3. Characterizations
4.4. Animals
4.5. Generation of Primary Immune Cells
4.6. Cell Viability Assay
4.7. Determination of Cytokine Release
4.8. Measurement of Reactive Oxygen Species
4.9. Migration Assay of Dendritic Cells
4.10. Evaluation of the Immune Competence of Primary Macrophages
4.11. In Vitro Cellular Uptake Assay
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MTS DC 24 h 2 kDa mPEG | 10% aqua bidestillata | 15 µg/mL 2 kDa mPEG | 50 µg/mL 2 kDa mPEG | 150 µg/mL 2 kDa mPEG | 5% DMSO | Background: 150 µg/mL 2 kDa mPEG | Background: MTS | |
|---|---|---|---|---|---|---|---|---|
| Biological replicate | 1 | 0.446 | 0.507 | 0.522 | 0.543 | 0.179 | 0.136 | / |
| 2 | 0.466 | 0.538 | 0.540 | 0.574 | 0.176 | 0.147 | / | |
| 3 | 0.901 | 1.006 | 1.055 | 1.065 | 0.185 | 0.139 | 0.095 | |
| 4 | 0.829 | 0.889 | 0.919 | 0.929 | 0.171 | 0.133 | 0.092 | |
| 5 | 0.517 | 0.562 | 0.587 | 0.575 | 0.175 | 0.122 | 0.069 | |
| 6 | 0.567 | 0.628 | 0.666 | 0.697 | 0.158 | 0.130 | 0.067 | |
| MTS DC 48 h 2 kDa mPEG | 10% aqua bidestillata | 15 µg/mL 2 kDa mPEG | 50 µg/mL 2 kDa mPEG | 150 µg/mL 2 kDa mPEG | 5% DMSO | Background: 150 µg/mL 2 kDa mPEG | Background: MTS | |
|---|---|---|---|---|---|---|---|---|
| Biological replicate | 1 | 0.390 | 0.525 | 0.542 | 0.616 | 0.239 | 0.135 | / |
| 2 | 0.373 | 0.474 | 0.550 | 0.595 | 0.211 | 0.123 | / | |
| 3 | 0.772 | 0.829 | 0.909 | 0.966 | 0.212 | 0.139 | 0.094 | |
| 4 | 0.682 | 0.763 | 0.810 | 0.884 | 0.175 | 0.109 | 0.092 | |
| 5 | 0.471 | 0.560 | 0.636 | 0.653 | 0.180 | 0.115 | 0.102 | |
| 6 | 0.603 | 0.679 | 0.790 | 0.847 | 0.179 | 0.120 | 0.099 | |
| MTS DC 24 h 5 kDa mPEG | 10% aqua bidestillata | 15 µg/mL 5 kDa mPEG | 50 µg/mL 5 kDa mPEG | 150 µg/mL 5 kDa mPEG | 5% DMSO | Background: 150 µg/mL 5 kDa mPEG | Background: MTS | |
|---|---|---|---|---|---|---|---|---|
| Biological replicate | 1 | 0.446 | 0.492 | 0.509 | 0.507 | 0.179 | 0.125 | / |
| 2 | 0.466 | 0.535 | 0.569 | 0.573 | 0.176 | 0.129 | / | |
| 3 | 0.901 | 0.995 | 1.030 | 1.082 | 0.185 | 0.126 | 0.095 | |
| 4 | 0.829 | 0.876 | 0.905 | 0.923 | 0.171 | 0.114 | 0.092 | |
| 5 | 0.517 | 0.613 | 0.599 | 0.597 | 0.175 | 0.114 | 0.069 | |
| 6 | 0.567 | 0.659 | 0.664 | 0.664 | 0.158 | 0.124 | 0.067 | |
| MTS DC 48 h 5 kDa mPEG | 10% aqua bidestillata | 15 µg/mL 5 kDa mPEG | 50 µg/mL 5 kDa mPEG | 150 µg/mL 5 kDa mPEG | 5% DMSO | Background: 150 µg/mL 5 kDa mPEG | Background: MTS | |
|---|---|---|---|---|---|---|---|---|
| Biological replicate | 1 | 0.390 | 0.506 | 0.540 | 0.591 | 0.239 | 0.125 | / |
| 2 | 0.373 | 0.509 | 0.563 | 0.613 | 0.211 | 0.124 | / | |
| 3 | 0.772 | 0.834 | 0.868 | 0.955 | 0.212 | 0.126 | 0.094 | |
| 4 | 0.682 | 0.736 | 0.792 | 0.864 | 0.175 | 0.104 | 0.092 | |
| 5 | 0.471 | 0.592 | 0.640 | 0.641 | 0.180 | 0.113 | 0.102 | |
| 6 | 0.603 | 0.725 | 0.778 | 0.800 | 0.179 | 0.119 | 0.099 | |
| MTS DC 24 h 10 kDa mPEG-FITC | 10% aqua bidestillata | 15 µg/mL 10 kDa mPEG-FITC | 50 µg/mL 10 kDa mPEG-FITC | 150 µg/mL 10 kDa mPEG-FITC | 5% DMSO | Background: 150 µg/mL 10 kDa mPEG-FITC | Background: MTS | |
|---|---|---|---|---|---|---|---|---|
| Biological replicate | 1 | 0.446 | 0.484 | 0.538 | 0.626 | 0.179 | 0.201 | / |
| 2 | 0.466 | 0.492 | 0.541 | 0.633 | 0.176 | 0.200 | / | |
| 3 | 0.901 | 0.968 | 1.007 | 1.084 | 0.185 | 0.211 | 0.095 | |
| 4 | 0.829 | 0.870 | 0.898 | 0.963 | 0.171 | 0.184 | 0.092 | |
| 5 | 0.517 | 0.575 | 0.610 | 0.671 | 0.175 | 0.206 | 0.069 | |
| 6 | 0.567 | 0.614 | 0.681 | 0.769 | 0.158 | 0.199 | 0.067 | |
| MTS DC 48 h 10 kDa mPEG-FITC | 10% aqua bidestillata | 15 µg/mL 10 kDa mPEG-FITC | 50 µg/mL 10 kDa mPEG-FITC | 150 µg/mL 10 kDa mPEG-FITC | 5% DMSO | Background: 150 µg/mL 10 kDa mPEG-FITC | Background: MTS | |
|---|---|---|---|---|---|---|---|---|
| Biological replicate | 1 | 0.390 | 0.493 | 0.584 | 0.703 | 0.239 | 0.222 | / |
| 2 | 0.373 | 0.490 | 0.579 | 0.692 | 0.211 | 0.175 | / | |
| 3 | 0.772 | 0.795 | 0.860 | 0.962 | 0.212 | 0.211 | 0.094 | |
| 4 | 0.682 | 0.729 | 0.770 | 0.884 | 0.175 | 0.177 | 0.092 | |
| 5 | 0.471 | 0.580 | 0.640 | 0.759 | 0.180 | 0.206 | 0.102 | |
| 6 | 0.603 | 0.752 | 0.808 | 0.975 | 0.179 | 0.191 | 0.099 | |
References
- Neumann, A.; Christel, A.; Kasper, C.; Behrens, P. BMP2-loaded nanoporous silica nanoparticles promote osteogenic differentiation of human mesenchymal stem cells. RSC Adv. 2013, 3, 24222–24230. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Haynes, C.L. Synthesis and Characterization of Biocompatible and Size-Tunable Multifunctional Porous Silica Nanoparticles. Chem. Mater. 2009, 21, 3979–3986. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef]
- Wei, H.; Hu, Y.; Wang, J.; Gao, X.; Qian, X.; Tang, M. Superparamagnetic Iron Oxide Nanoparticles: Cytotoxicity, Metabolism, and Cellular Behavior in Biomedicine Applications. Int. J. Nanomed. 2021, 16, 6097–6113. [Google Scholar] [CrossRef]
- Al-Jamal, K.T.; Bai, J.; Wang, J.T.-W.; Protti, A.; Southern, P.; Bogart, L.; Heidari, H.; Li, X.; Cakebread, A.; Asker, D.; et al. Magnetic Drug Targeting: Preclinical in Vivo Studies, Mathematical Modeling, and Extrapolation to Humans. Nano Lett. 2016, 16, 5652–5660. [Google Scholar] [CrossRef] [Green Version]
- Morishige, K.; Kacher, D.F.; Libby, P.; Josephson, L.; Ganz, P.; Weissleder, R.; Aikawa, M. High-resolution magnetic resonance imaging enhanced with superparamagnetic nanoparticles measures macrophage burden in atherosclerosis. Circulation 2010, 122, 1707–1715. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.-F.; Chang, Y.; Xia, N. Recent Development of Nanomaterials-Based Cytosensors for the Detection of Circulating Tumor Cells. Biosensors 2021, 11, 281. [Google Scholar] [CrossRef]
- Lee, H.; Sun, E.; Ham, D.; Weissleder, R. Chip-NMR biosensor for detection and molecular analysis of cells. Nat. Med. 2008, 14, 869–874. [Google Scholar] [CrossRef] [Green Version]
- Maier-Hauff, K.; Ulrich, F.; Nestler, D.; Niehoff, H.; Wust, P.; Thiesen, B.; Orawa, H.; Budach, V.; Jordan, A. Efficacy and safety of intratumoral thermotherapy using magnetic iron-oxide nanoparticles combined with external beam radiotherapy on patients with recurrent glioblastoma multiforme. J. Neurooncol. 2011, 103, 317–324. [Google Scholar] [CrossRef] [Green Version]
- Grauer, O.; Jaber, M.; Hess, K.; Weckesser, M.; Schwindt, W.; Maring, S.; Wölfer, J.; Stummer, W. Combined intracavitary thermotherapy with iron oxide nanoparticles and radiotherapy as local treatment modality in recurrent glioblastoma patients. J. Neurooncol. 2019, 141, 83–94. [Google Scholar] [CrossRef]
- Chen, W.; Schilperoort, M.; Cao, Y.; Shi, J.; Tabas, I.; Tao, W. Macrophage-targeted nanomedicine for the diagnosis and treatment of atherosclerosis. Nat. Rev. Cardiol. 2022, 19, 228–249. [Google Scholar] [CrossRef]
- Obermeier, A.; Kuchler, S.; Matl, F.D.; Pirzer, T.; Stemberger, A.; Mykhaylyk, O.; Friess, W.; Burgkart, R. Magnetic Drug Targeting as New Therapeutic Option for the Treatment of Biomaterial Infections. J. Biomater. Sci. Polym. Ed. 2012, 23, 2321–2336. [Google Scholar] [CrossRef]
- Lübbe, A.S.; Bergemann, C.; Riess, H.; Schriever, F.; Reichardt, P.; Possinger, K.; Matthias, M.; Dörken, B.; Herrmann, F.; Gürtler, R.; et al. Clinical experiences with magnetic drug targeting: A phase I study with 4’-epidoxorubicin in 14 patients with advanced solid tumors. Cancer Res. 1996, 56, 4686–4693. [Google Scholar]
- Alexiou, C.; Jurgons, R.; Schmid, R.; Hilpert, A.; Bergemann, C.; Parak, F.; Iro, H. In vitro and in vivo investigations of targeted chemotherapy with magnetic nanoparticles. J. Magn. Magn. Mater. 2005, 293, 389–393. [Google Scholar] [CrossRef]
- Aghanejad, A.; Babamiri, H.; Adibkia, K.; Barar, J.; Omidi, Y. Mucin-1 aptamer-armed superparamagnetic iron oxide nanoparticles for targeted delivery of doxorubicin to breast cancer cells. Bioimpacts 2018, 8, 117–127. [Google Scholar] [CrossRef]
- Janssen, H.C.; Angrisani, N.; Kalies, S.; Hansmann, F.; Kietzmann, M.; Warwas, D.P.; Behrens, P.; Reifenrath, J. Biodistribution, biocompatibility and targeted accumulation of magnetic nanoporous silica nanoparticles as drug carrier in orthopedics. J. Nanobiotechnol. 2020, 18, 14. [Google Scholar] [CrossRef] [Green Version]
- Fam, S.Y.; Chee, C.F.; Yong, C.Y.; Ho, K.L.; Mariatulqabtiah, A.R.; Tan, W.S. Stealth Coating of Nanoparticles in Drug-Delivery Systems. Nanomaterials 2020, 10, 787. [Google Scholar] [CrossRef] [Green Version]
- Arvizo, R.R.; Miranda, O.R.; Moyano, D.F.; Walden, C.A.; Giri, K.; Bhattacharya, R.; Robertson, J.D.; Rotello, V.M.; Reid, J.M.; Mukherjee, P. Modulating pharmacokinetics, tumor uptake and biodistribution by engineered nanoparticles. PLoS ONE 2011, 6, e24374. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Nie, H.; Wang, K.; Tan, W.; Wu, X.; Zhang, P. In vivo study of biodistribution and urinary excretion of surface-modified silica nanoparticles. Anal. Chem. 2008, 80, 9597–9603. [Google Scholar] [CrossRef]
- Perrault, S.D.; Walkey, C.; Jennings, T.; Fischer, H.C.; Chan, W.C.W. Mediating Tumor Targeting Efficiency of Nanoparticles Through Design. Nano Lett. 2009, 9, 1909–1915. [Google Scholar] [CrossRef]
- Kayal, S.; Ramanujan, R.V. Doxorubicin loaded PVA coated iron oxide nanoparticles for targeted drug delivery. Mater. Sci. Eng. C 2010, 30, 484–490. [Google Scholar] [CrossRef]
- Mojica Pisciotti, M.L.; Lima, E., Jr.; Vasquez Mansilla, M.; Tognoli, V.E.; Troiani, H.E.; Pasa, A.A.; Creczynski-Pasa, T.B.; Silva, A.H.; Gurman, P.; Colombo, L.; et al. In vitro and in vivo experiments with iron oxide nanoparticles functionalized with DEXTRAN or polyethylene glycol for medical applications: Magnetic targeting. J. Biomed. Mater. Res. B Appl. Biomater. 2014, 102, 860–868. [Google Scholar] [CrossRef]
- Lipka, J.; Semmler-Behnke, M.; Sperling, R.A.; Wenk, A.; Takenaka, S.; Schleh, C.; Kissel, T.; Parak, W.J.; Kreyling, W.G. Biodistribution of PEG-modified gold nanoparticles following intratracheal instillation and intravenous injection. Biomaterials 2010, 31, 6574–6581. [Google Scholar] [CrossRef]
- Fang, C.; Shi, B.; Pei, Y.-Y.; Hong, M.-H.; Wu, J.; Chen, H.-Z. In vivo tumor targeting of tumor necrosis factor-α-loaded stealth nanoparticles: Effect of MePEG molecular weight and particle size. Eur. J. Pharm. Sci. 2006, 27, 27–36. [Google Scholar] [CrossRef]
- Harris, J.M.; Chess, R.B. Effect of pegylation on pharmaceuticals. Nat. Rev. Drug Discov. 2003, 2, 214–221. [Google Scholar] [CrossRef]
- Ganson, N.J.; Povsic, T.J.; Sullenger, B.A.; Alexander, J.H.; Zelenkofske, S.L.; Sailstad, J.M.; Rusconi, C.P.; Hershfield, M.S. Pre-existing anti–polyethylene glycol antibody linked to first-exposure allergic reactions to pegnivacogin, a PEGylated RNA aptamer. J. Allergy Clin. Immunol. 2016, 137, 1610–1613.e1617. [Google Scholar] [CrossRef] [Green Version]
- McSweeney, M.D.; Wessler, T.; Price, L.S.L.; Ciociola, E.C.; Herity, L.B.; Piscitelli, J.A.; Zamboni, W.C.; Forest, M.G.; Cao, Y.; Lai, S.K. A minimal physiologically based pharmacokinetic model that predicts anti-PEG IgG-mediated clearance of PEGylated drugs in human and mouse. J. Control. Release Off. J. Control. Release Soc. 2018, 284, 171–178. [Google Scholar] [CrossRef]
- Yang, Q.; Jacobs, T.M.; McCallen, J.D.; Moore, D.T.; Huckaby, J.T.; Edelstein, J.N.; Lai, S.K. Analysis of Pre-existing IgG and IgM Antibodies against Polyethylene Glycol (PEG) in the General Population. Anal. Chem. 2016, 88, 11804–11812. [Google Scholar] [CrossRef]
- International Organization for Standardization. ISO 10993-5:2009-06: Biological Evaluation of Medical Devices-Part 5: Tests for In Vitro Cytotoxicity; International Organization for Standardization: Geneva, Switzerland, 2009. [Google Scholar] [CrossRef]
- Janssen, H.C.; Warwas, D.P.; Dahlhaus, D.; Meissner, J.; Taptimthong, P.; Kietzmann, M.; Behrens, P.; Reifenrath, J.; Angrisani, N. In vitro and in vivo accumulation of magnetic nanoporous silica nanoparticles on implant materials with different magnetic properties. J. Nanobiotechnol. 2018, 16, 96. [Google Scholar] [CrossRef] [Green Version]
- Au, C.; Mutkus, L.; Dobson, A.; Riffle, J.; Lalli, J.; Aschner, M. Effects of Nanoparticles on the Adhesion and Cell Viability on Astrocytes. Biol. Trace Elem. Res. 2007, 120, 248–256. [Google Scholar] [CrossRef]
- Demine, S.; Renard, P.; Arnould, T. Mitochondrial Uncoupling: A Key Controller of Biological Processes in Physiology and Diseases. Cells 2019, 8, 795. [Google Scholar] [CrossRef] [Green Version]
- Mahmoudi, M.; Hofmann, H.; Rothen-Rutishauser, B.; Petri-Fink, A. Assessing the In Vitro and In Vivo Toxicity of Superparamagnetic Iron Oxide Nanoparticles. Chem. Rev. 2012, 112, 2323–2338. [Google Scholar] [CrossRef] [Green Version]
- Patil, R.M.; Thorat, N.D.; Shete, P.B.; Bedge, P.A.; Gavde, S.; Joshi, M.G.; Tofail, S.A.M.; Bohara, R.A. Comprehensive cytotoxicity studies of superparamagnetic iron oxide nanoparticles. Biochem. Biophys. Rep. 2018, 13, 63–72. [Google Scholar] [CrossRef]
- Kunzmann, A.; Andersson, B.; Vogt, C.; Feliu, N.; Ye, F.; Gabrielsson, S.; Toprak, M.S.; Buerki-Thurnherr, T.; Laurent, S.; Vahter, M.; et al. Efficient internalization of silica-coated iron oxide nanoparticles of different sizes by primary human macrophages and dendritic cells. Toxicol. Appl. Pharmacol. 2011, 253, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.; Lim, J.-S. Induction of functional changes of dendritic cells by silica nanoparticles. Immune Netw. 2012, 12, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Jin, R.; Feng, T.; Liu, L.; Yang, L.; Tao, Y.; Anderson, J.M.; Ai, H.; Li, H. Iron oxide nanoparticles and induced autophagy in human monocytes. Int. J. Nanomed. 2017, 12, 3993–4005. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Liu, Y.; Huang, J.; Chen, K.; Huang, J.; Xiao, K. Uptake, distribution, clearance, and toxicity of iron oxide nanoparticles with different sizes and coatings. Sci. Rep. 2018, 8, 2082. [Google Scholar] [CrossRef] [Green Version]
- Calero, M.; Gutiérrez, L.; Salas, G.; Luengo, Y.; Lázaro, A.; Acedo, P.; Morales, M.P.; Miranda, R.; Villanueva, A. Efficient and safe internalization of magnetic iron oxide nanoparticles: Two fundamental requirements for biomedical applications. Nanomed. Nanotechnol. Biol. Med. 2014, 10, 733–743. [Google Scholar] [CrossRef]
- Díaz, B.; Sánchez-Espinel, C.; Arruebo, M.; Faro, J.; de Miguel, E.; Magadán, S.; Yagüe, C.; Fernández-Pacheco, R.; Ibarra, M.R.; Santamaría, J.; et al. Assessing Methods for Blood Cell Cytotoxic Responses to Inorganic Nanoparticles and Nanoparticle Aggregates. Small 2008, 4, 2025–2034. [Google Scholar] [CrossRef]
- Hanini, A.; Schmitt, A.; Kacem, K.; Chau, F.; Ammar, S.; Gavard, J. Evaluation of iron oxide nanoparticle biocompatibility. Int. J. Nanomed. 2011, 6, 787–794. [Google Scholar] [CrossRef]
- Malvindi, M.A.; De Matteis, V.; Galeone, A.; Brunetti, V.; Anyfantis, G.C.; Athanassiou, A.; Cingolani, R.; Pompa, P.P. Toxicity Assessment of Silica Coated Iron Oxide Nanoparticles and Biocompatibility Improvement by Surface Engineering. PLoS ONE 2014, 9, e85835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesárošová, M.; Kozics, K.; Bábelová, A.; Regendová, E.; Pastorek, M.; Vnuková, D.; Buliaková, B.; Rázga, F.; Gábelová, A. The role of reactive oxygen species in the genotoxicity of surface-modified magnetite nanoparticles. Toxicol. Lett. 2014, 226, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Lehman, S.E.; Morris, A.S.; Mueller, P.S.; Salem, A.K.; Grassian, V.H.; Larsen, S.C. Silica nanoparticle-generated ROS as a predictor of cellular toxicity: Mechanistic insights and safety by design. Environ. Sci. Nano 2016, 3, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Witasp, E.; Shvedova, A.A.; Kagan, V.E.; Fadeel, B. Single-walled carbon nanotubes impair human macrophage engulfment of apoptotic cell corpses. Inhal. Toxicol. 2009, 21, 131–136. [Google Scholar] [CrossRef]
- Bäumer, W.; Tschernig, T.; Sülzle, B.; Seegers, U.; Lührmann, A.; Kietzmann, M. Effects of cilomilast on dendritic cell function in contact sensitivity and dendritic cell migration through skin. Eur. J. Pharmacol. 2003, 481, 271–279. [Google Scholar] [CrossRef]
- Lutz, M.B.; Kukutsch, N.; Ogilvie, A.L.J.; Rößner, S.; Koch, F.; Romani, N.; Schuler, G. An advanced culture method for generating large quantities of highly pure dendritic cells from mouse bone marrow. J. Immunol. Methods 1999, 223, 77–92. [Google Scholar] [CrossRef]
- Vanbervliet, B.; Homey, B.; Durand, I.; Massacrier, C.; Aït-Yahia, S.; De Bouteiller, O.; Vicari, A.; Caux, C. Sequential involvement of CCR2 and CCR6 ligands for immature dendritic cell recruitment: Possible role at inflamed epithelial surfaces. Eur. J. Immunol. 2002, 32, 231–242. [Google Scholar] [CrossRef]
- Gudkov, S.V.; Burmistrov, D.E.; Serov, D.A.; Rebezov, M.B.; Semenova, A.A.; Lisitsyn, A.B. Do Iron Oxide Nanoparticles Have Significant Antibacterial Properties? Antibiotics 2021, 10, 884. [Google Scholar] [CrossRef] [PubMed]
- Foth, F. Grundlegende Untersuchungen zum Implant-Directed-Magnetic-Drug-Targeting und zu Seinem Möglichen, Zukünftigen Einsatz bei der Behandlung von Implantat-Assoziierten-Infektionen in der Orthopädischen Chirurgie. Ph.D. Thesis, University of Veterinary Medicine Hannover, Hannover, Germany, 2014. [Google Scholar]
- Pelaz, B.; del Pino, P.; Maffre, P.; Hartmann, R.; Gallego, M.; Rivera-Fernández, S.; de la Fuente, J.M.; Nienhaus, G.U.; Parak, W.J. Surface Functionalization of Nanoparticles with Polyethylene Glycol: Effects on Protein Adsorption and Cellular Uptake. ACS Nano 2015, 9, 6996–7008. [Google Scholar] [CrossRef]
- Jenkins, S.I.; Weinberg, D.; al-Shakli, A.F.; Fernandes, A.R.; Yiu, H.H.P.; Telling, N.D.; Roach, P.; Chari, D.M. ‘Stealth’ nanoparticles evade neural immune cells but also evade major brain cell populations: Implications for PEG-based neurotherapeutics. J. Control. Release 2016, 224, 136–145. [Google Scholar] [CrossRef]
- Graf, C.; Gao, Q.; Schütz, I.; Noufele, C.N.; Ruan, W.; Posselt, U.; Korotianskiy, E.; Nordmeyer, D.; Rancan, F.; Hadam, S.; et al. Surface Functionalization of Silica Nanoparticles Supports Colloidal Stability in Physiological Media and Facilitates Internalization in Cells. Langmuir 2012, 28, 7598–7613. [Google Scholar] [CrossRef] [PubMed]
- Halamoda-Kenzaoui, B.; Ceridono, M.; Urbán, P.; Bogni, A.; Ponti, J.; Gioria, S.; Kinsner-Ovaskainen, A. The agglomeration state of nanoparticles can influence the mechanism of their cellular internalisation. J. Nanobiotechnol. 2017, 15, 48. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.L.; Harada, T.; Christian, D.A.; Pantano, D.A.; Tsai, R.K.; Discher, D.E. Minimal “Self” peptides that inhibit phagocytic clearance and enhance delivery of nanoparticles. Science 2013, 339, 971–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuan, M.; Shao, J.; Dai, L.; He, Q.; Li, J. Macrophage Cell Membrane Camouflaged Mesoporous Silica Nanocapsules for In Vivo Cancer Therapy. Adv. Healthc. Mater. 2015, 4, 1645–1652. [Google Scholar] [CrossRef] [PubMed]
- Cauda, V.; Argyo, C.; Bein, T. Impact of different PEGylation patterns on the long-term bio-stability of colloidal mesoporous silica nanoparticles. J. Mater. Chem. 2010, 20, 8693–8699. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Tsai, C.-P.; Huang, H.-Y.; Kuo, C.-T.; Hung, Y.; Huang, D.-M.; Chen, Y.-C.; Mou, C.-Y. Well-Ordered Mesoporous Silica Nanoparticles as Cell Markers. Chem. Mater. 2005, 17, 4570–4573. [Google Scholar] [CrossRef]
- Bäumer, W.; Wendorff, S.; Gutzmer, R.; Werfel, T.; Dijkstra, D.; Chazot, P.; Stark, H.; Kietzmann, M. Histamine H4 receptors modulate dendritic cell migration through skin–immunomodulatory role of histamine. Allergy 2008, 63, 1387–1394. [Google Scholar] [CrossRef] [PubMed]
- Roth, I.; Schumacher, S.; Basler, T.; Baumert, K.; Seitz, J.M.; Evertz, F.; Muller, P.P.; Baumer, W.; Kietzmann, M. Magnesium corrosion particles do not interfere with the immune function of primary human and murine macrophages. Prog. Biomater. 2015, 4, 21–30. [Google Scholar] [CrossRef] [PubMed]








| Functionalization | Blank MNPSNP | 2 kDa mPEG-FITC | 5 kDa mPEG-FITC | 10 kDa mPEG-FITC |
|---|---|---|---|---|
| BET [m2·g−1] | 910 | 730 | 610 | 640 |
| d [nm] | 3.6 | 3.4 | 3.0 | 3.4 |
| V [cm3 g−1] | 0.7 | 0.5 | 0.3 | 0.4 |
| ZP [mV] | −30 | −38 | −35 | −36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Storjohann, R.; Gericke, B.; Reifenrath, J.; Herrmann, T.; Behrens, P.; Oltmanns, H.; Meißner, J. Influence of PEG Chain Length of Functionalized Magnetic Nanoparticles on the Cytocompatibility and Immune Competence of Primary Murine Macrophages and Dendritic Cells. Int. J. Mol. Sci. 2023, 24, 2565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032565
Storjohann R, Gericke B, Reifenrath J, Herrmann T, Behrens P, Oltmanns H, Meißner J. Influence of PEG Chain Length of Functionalized Magnetic Nanoparticles on the Cytocompatibility and Immune Competence of Primary Murine Macrophages and Dendritic Cells. International Journal of Molecular Sciences. 2023; 24(3):2565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032565
Chicago/Turabian StyleStorjohann, Ronja, Birthe Gericke, Janin Reifenrath, Timo Herrmann, Peter Behrens, Hilke Oltmanns, and Jessica Meißner. 2023. "Influence of PEG Chain Length of Functionalized Magnetic Nanoparticles on the Cytocompatibility and Immune Competence of Primary Murine Macrophages and Dendritic Cells" International Journal of Molecular Sciences 24, no. 3: 2565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24032565