
Genome-Wide Identification Analysis of the R2R3-MYB Transcription Factor Family in Cymbidium sinense for Insights into Drought Stress Responses

 ,
,
Abstract
:1. Introduction
2. Results
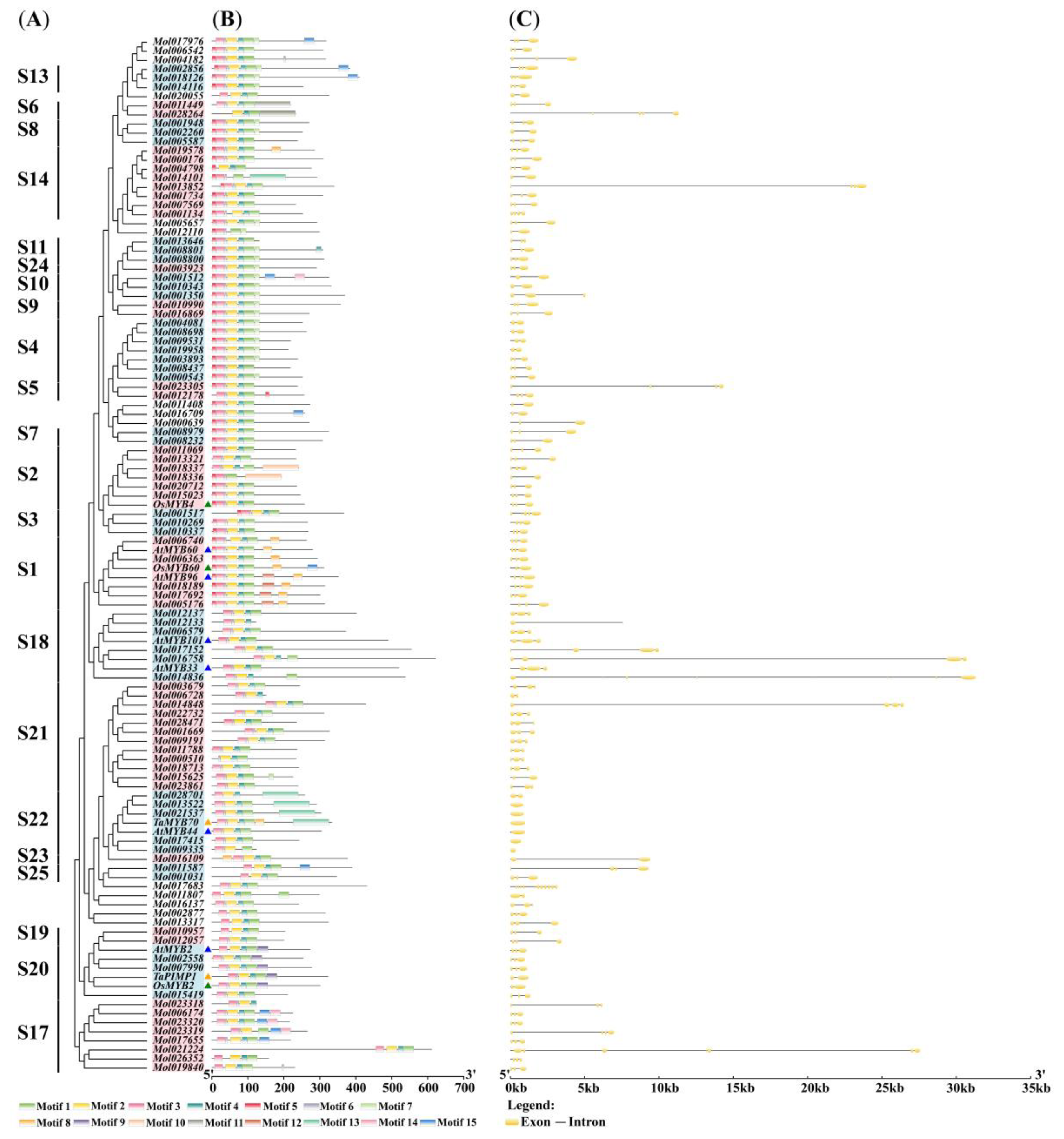
2.1. Identification and Phylogenetic Analysis of the R2R3-MYB Genes in C. sinense
2.2. Gene Structure and Domain Analysis of CsMYBs and DrMYBs
2.3. Collinearity and Purifying Selection Analysis of CsMYB Genes
2.4. Protein Secondary and Tertiary Structures Prediction
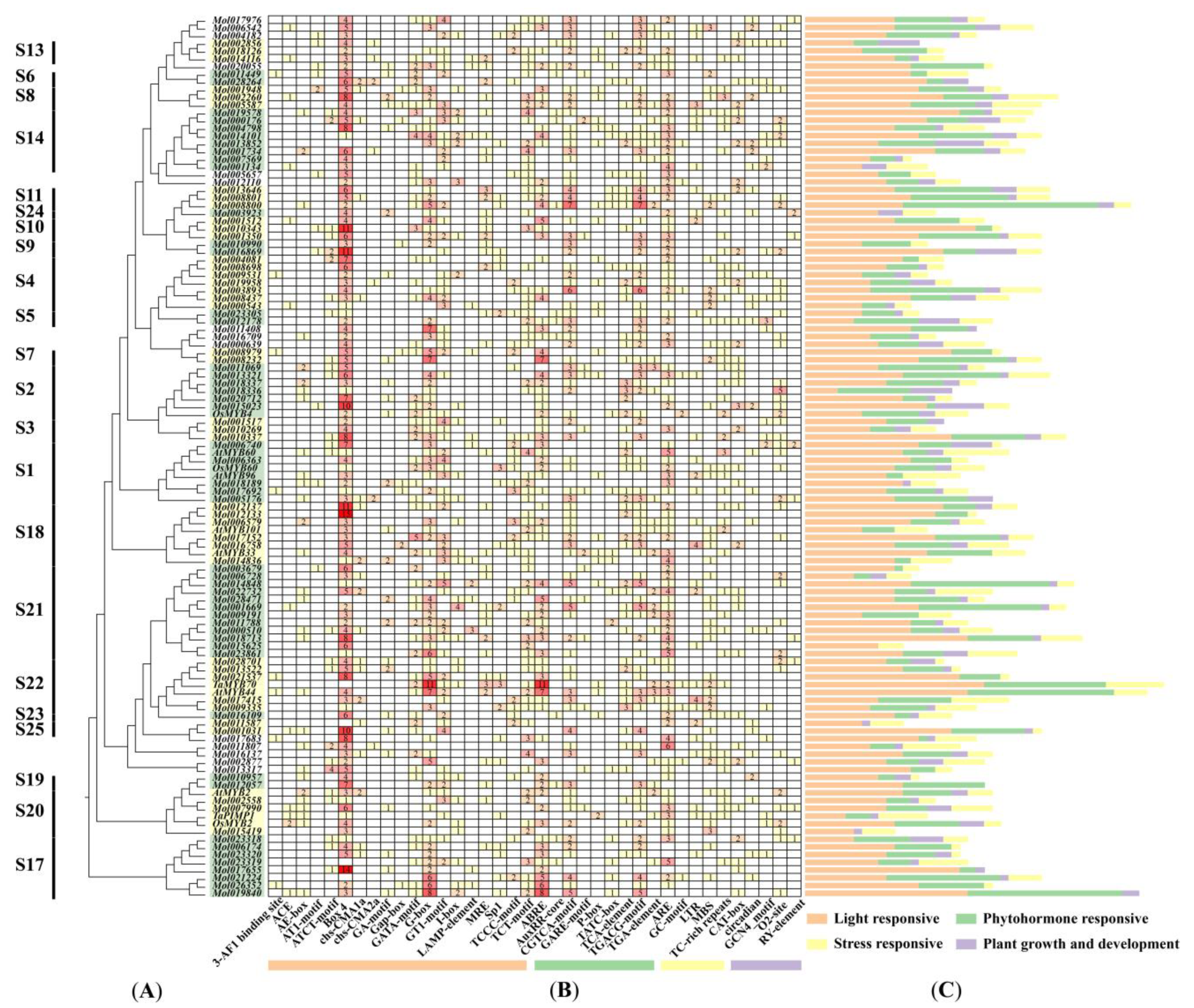
2.5. Cis-Acting Regulatory Elements Analysis in CsMYB and DrMYB Genes
2.6. Expression Patterns Analysis of CsMYB Genes under Drought Stress in Leaves and Roots
3. Discussion
4. Materials and Methods
4.1. Identification and Phylogenetic Analysis of R2R3-MYB Genes in C. sinense
4.2. Intron–Exon Structure and Conserved Domain Analysis of R2R3-MYB Genes
4.3. Collinearity and Selective Pressure
4.4. Protein Secondary and Tertiary Structures Prediction of DrMYBs and CsMYBs
4.5. Cis-Acting Element Analysis
4.6. Expression Analysis of R2R3-MYB Genes under Drought Stress in Different Tissues of C. sinense
4.7. qRT–PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, C.; Zhang, K.X.; Khurshid, M.; Li, J.B.; He, M.; Georgiev, M.I.; Zhang, X.Q.; Zhou, M.L. MYB Transcription Repressors Regulate Plant Secondary Metabolism. Crit. Rev. Plant Sci. 2019, 38, 159–170. [Google Scholar] [CrossRef]
- Baldoni, E.; Genga, A.; Cominelli, E. Plant MYB Transcription Factors: Their Role in Drought Response Mechanisms. Int. J. Mol. Sci. 2015, 16, 15811–15851. [Google Scholar] [CrossRef] [PubMed]
- Ogata, K.; Morikawa, S.; Nakamura, H.; Sekikawa, A.; Inoue, T.; Kanai, H.; Sarai, A.; Ishii, S.; Nishimura, Y. Solution Structure of a Specific DNA Complex of the Myb DNA-Binding Domain with Cooperative Recognition Helices. Cell 1994, 79, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Millard, P.S.; Kragelund, B.B.; Burow, M. R2R3 MYB Transcription Factors–Functions Outside the DNA-Binding Domain. Trends Plant Sci. 2019, 24, 934–946. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB Transcription Factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef]
- Stracke, R.; Werber, M.; Weisshaar, B. The R2R3-MYB Gene Family in Arabidopsis thaliana. Curr. Opin. Plant Biol. 2001, 4, 447–456. [Google Scholar] [CrossRef]
- Fan, H.; Cui, M.; Li, N.; Li, X.; Liang, Y.; Liu, L.; Cai, Y.; Lin, Y. Genome-Wide Identification and Expression Analyses of R2R3-MYB Transcription Factor Genes from Two Orchid Species. PeerJ 2020, 8. [Google Scholar] [CrossRef]
- Oh, J.E.; Kwon, Y.; Kim, J.H.; Noh, H.; Hong, S.W.; Lee, H. A Dual Role for MYB60 in Stomatal Regulation and Root Growth of Arabidopsis Thaliana under Drought Stress. Plant Mol. Biol. 2011, 77, 91–103. [Google Scholar] [CrossRef]
- Seo, P.J.; Xiang, F.; Qiao, M.; Park, J.-Y.; Lee, Y.N.; Kim, S.-G.; Lee, Y.-H.; Park, W.J.; Park, C.-M. The MYB96 Transcription Factor Mediates Abscisic Acid Signaling during Drought Stress Response in Arabidopsis. Plant Physiol. 2009, 151, 275–289. [Google Scholar] [CrossRef]
- Jung, C.; Jun, S.S.; Sang, W.H.; Yeon, J.K.; Chung, H.K.; Sang, I.S.; Baek, H.N.; Yang, D.C.; Cheong, J.J. Overexpression of AtMYB44 Enhances Stomatal Closure to Confer Abiotic Stress Tolerance in Transgenic Arabidopsis. Plant Physiol. 2008, 146, 623–635. [Google Scholar] [CrossRef] [Green Version]
- Jian, L.; Kang, K.; Choi, Y.; Suh, M.C.; Paek, N. Mutation of OsMYB60 Reduces Rice Resilience to Drought Stress by Attenuating Cuticular Wax Biosynthesis. Plant J. 2022, 112, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Dai, X.; Zhang, W.-H. A R2R3-Type MYB Gene, OsMYB2, Is Involved in Salt, Cold, and Dehydration Tolerance in Rice. J. Exp. Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef] [PubMed]
- Vannini, C.; Iriti, M.; Bracale, M.; Locatelli, F.; Faoro, F.; Croce, P.; Pirona, R.; di Maro, A.; Coraggio, I.; Genga, A. The Ectopic Expression of the Rice Osmyb4 Gene in Arabidopsis Increases Tolerance to Abiotic, Environmental and Biotic Stresses. Physiol. Mol. Plant Pathol. 2006, 69, 26–42. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, X.; Wang, X.; Zhou, M.; Zhou, X.; Ye, X.; Wei, X. An R2R3 MYB Transcription Factor in Wheat, TaPIMP1, Mediates Host Resistance to Bipolaris Sorokiniana and Drought Stresses through Regulation of Defense- and Stress-Related Genes. New Phytol. 2012, 196, 1155–1170. [Google Scholar] [CrossRef]
- Mao, H.; Jian, C.; Cheng, X.; Chen, B.; Mei, F.; Li, F.; Zhang, Y.; Li, S.; Du, L.; Li, T.; et al. The Wheat ABA Receptor Gene TaPYL1-1B Contributes to Drought Tolerance and Grain Yield by Increasing Water-Use Efficiency. Plant Biotechnol. J. 2022, 20, 846–861. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, Y.; Li, J.; Qin, J.; Zhang, W.; Huang, W.; Hu, H. Physiological Diversity of Orchids. Plant Divers. 2018, 40, 196–208. [Google Scholar] [CrossRef]
- Chen, X.Q.; Liu, Z.J.; Zhu, G.H.; Lang, K.Y.; Tsi, Z.H.; Luo, Y.B.; Jin, X.H.; Cribb, P.J.; Wood, J.J.; Gale, S.W. Flora of China, Vol. 25; Science Press and Missouri Botanical Garden Press: St. Louis, MO, USA, 2009; pp. 260–280. [Google Scholar]
- Liu, Z.J.; Chen, L.J.; Liu, K.W.; Li, L.Q.; Zhang, Y.T.; Huang, L.Q. Climate Warming Brings about Extinction Tendency in Wild Population of Cymbidium Sinense. Acta Ecol. Sin. 2009, 29, 3443–3455. [Google Scholar]
- Wei, Y.L.; Jin, J.P.; Liang, D.; Gao, J.; Li, J.; Xie, Q.; Lu, C.Q.; Yang, F.X.; Zhu, G.F. Genome-Wide Identification of Cymbidium Sinense WRKY Gene Family and the Importance of Its Group III Members in Response to Abiotic Stress. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Zheng, X.N.; Wen, Z.Q.; Pan, R.C.; Hew, C.S. Response of Cymbidium sinense to Drought Stress. J. Hortic. Sci. 1992, 67, 295–299. [Google Scholar] [CrossRef]
- Yang, F.-X.; Gao, J.; Wei, Y.-L.; Ren, R.; Zhang, G.-Q.; Lu, C.-Q.; Jin, J.-P.; Ai, Y.; Wang, Y.-Q.; Chen, L.-J.; et al. The Genome of Cymbidium sinense Revealed the Evolution of Orchid Traits. Plant Biotechnol. J. 2021, 19, 2501–2516. [Google Scholar] [CrossRef]
- Yujie, K.E.; Mingkun, C.; Shanhu, M.A.; Yue, O.U.; Yi, W.; Qingdong, Z.; Zhongjian, L.I.U.; Ye, A.I. Research Progress of MYB Transcription Factors in Orchidaceae. Acta Hortic. Sin. 2021, 48, 2311. [Google Scholar]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and Analyzing DNA and Protein Sequence Motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Wilkins, O.; Nahal, H.; Foong, J.; Provart, N.J.; Campbell, M.M. Expansion and Diversification of the Populus R2R3-MYB Family of Transcription Factors. Plant Physiol. 2009, 149, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Bartley, L.E. Comparative Genomic Analysis of the R2R3 MYB Secondary Cell Wall Regulators of Arabidopsis, Poplar, Rice, Maize, and Switchgrass. BMC Plant Biol. 2014, 14. [Google Scholar] [CrossRef]
- Wang, X.; Liang, L.; Li, L.; Wang, T. Genome-wide analysis of R2R3-MYB transcription factors in Phalaenopsis equestris. For. Res. 2018, 31, 104–113. (In Chinese) [Google Scholar]
- Ke, Y.J.; Zheng, Q.D.; Yao, Y.H.; Ou, Y.; Chen, J.Y.; Wang, M.J.; Lai, H.P.; Yan, L.; Liu, Z.J.; Ai, Y. Genome-Wide Identification of the Myb Gene Family in Cymbidium ensifolium and Its Expression Analysis in Different Flower Colors. Int. J. Mol. Sci. 2021, 22, 13245. [Google Scholar] [CrossRef]
- Chen, J.; Bi, Y.-Y.; Wang, Q.-Q.; Liu, D.-K.; Zhang, D.; Ding, X.; Liu, Z.-J.; Chen, S.-P. Genome-Wide Identification and Analysis of Anthocyanin Synthesis-Related R2R3-MYB Genes in Cymbidium goeringii. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef]
- Lixiang, J.U.; Xin, L.E.I.; Chengzhi, Z.; Huangying, S.H.U.; Zhiwei, W.; Shanhan, C. Identification of MYB Family Genes and Its Relationship with Pungency of Pepper. Acta Hortic. Sin. 2020, 47, 875. [Google Scholar]
- Bowers, J.E.; Chapman, B.A.; Rong, J.; Paterson, A.H. Unravelling Angiosperm Genome Evolution by Phylogenetic Analysis of Chromosomal Duplication Events. Nature 2003, 422, 433–438. [Google Scholar] [CrossRef]
- Ai, Y.; Li, Z.; Sun, W.-H.; Chen, J.; Zhang, D.; Ma, L.; Zhang, Q.-H.; Chen, M.-K.; Zheng, Q.-D.; Liu, J.-F.; et al. The Cymbidium Genome Reveals the Evolution of Unique Morphological Traits. Hortic. Res. 2021, 8, 255. [Google Scholar] [CrossRef] [PubMed]
- Bi, H.; Luang, S.; Li, Y.; Bazanova, N.; Morran, S.; Song, Z.; Perera, M.A.; Hrmova, M.; Borisjuk, N.; Lopato, S. Identification and Characterization of Wheat Drought-Responsive MYB Transcription Factors Involved in the Regulation of Cuticle Biosynthesis. J. Exp. Bot. 2016, 67, 5363–5380. [Google Scholar] [CrossRef] [PubMed]
- Butt, H.I.; Yang, Z.; Gong, Q.; Chen, E.; Wang, X.; Zhao, G.; Ge, X.; Zhang, X.; Li, F. GaMYB85, an R2R3 MYB Gene, in Transgenic Arabidopsis Plays an Important Role in Drought Tolerance. BMC Plant Biol. 2017, 17, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Li, X.; Li, Y.; Yang, G.; Liu, W.; Shao, B.; Zhong, J.; Huang, P.; Han, D. Overexpression of a Malus baccata MYB Transcription Factor Gene MbMYB4 Increases Cold and Drought Tolerance in Arabidopsis thaliana. Int. J. Mol. Sci. 2022, 23, 1794. [Google Scholar] [CrossRef]
- Wu, Y.; Li, T.; Cheng, Z.; Zhao, D.; Tao, J.; Arasimowicz-Jelonek, M.; Kosmala, A.; Sobieszczuk-Nowicka, E.; Aroca, R. R2R3-MYB Transcription Factor PlMYB108 Confers Drought Tolerance in Herbaceous peony (Paeonia Lactiflora Pall.). Int. J. Mol. Sci. 2021, 22, 11884. [Google Scholar] [CrossRef]
- Cao, Z.H.; Zhang, S.Z.; Wang, R.K.; Zhang, R.F.; Hao, Y.J. Genome Wide Analysis of the Apple MYB Transcription Factor Family Allows the Identification of MdoMYB121 Gene Confering Abiotic Stress Tolerance in Plants. PLoS One 2013, 8, e69955. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Rozewicki, J.; Li, S.; Amada, K.M.; Standley, D.M.; Katoh, K. MAFFT-DASH: Integrated Protein Sequence and Structural Alignment. Nucleic Acids Res. 2019, 47, W5–W10. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES Science Gateway: A Community Resource for Phylogenetic Analyses. In Proceedings of the Proceedings of the TeraGrid 2011 Conference: Extreme Digital Discovery, TG’11, Salt Lake City, UT, USA, 18–21 July 2011. [Google Scholar] [CrossRef]
- He, Z.; Zhang, H.; Gao, S.; Lercher, M.J.; Chen, W.-H.; Hu, S. Evolview v2: An Online Visualization and Management Tool for Customized and Annotated Phylogenetic Trees. Nucleic Acids Res. 2016, 44, W236–W241. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Liu, B.B.; Wang, L.J.; Zhang, L.; Hu, J.J.; Chen, J.; Zheng, H.Q.; Lu, M.Z. Characterization of the Populus Rab Family Genes and the Function of PtRabE1b in Salt Tolerance. BMC Plant Biol. 2018, 18. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, J.; Zhao, X.-Q.; Wang, J.; Wong, G.K.-S.; Yu, J. KaKs_calculator: Calculating Ka and Ks through Model Selection and Model Averaging. Genom. Proteom. Bioinform. 2006, 4, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Q.; Zhu, M.J.; Yu, X.; Bi, Y.Y.; Zhou, Z.; Chen, M.K.; Chen, J.; Zhang, D.; Ai, Y.; Liu, Z.J.; et al. Genome-Wide Identification and Expression Analysis of Terpene Synthase Genes in Cymbidium faberi. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Geourjon, C.; Deleage, G. SOPMA: Significant Improvements in Protein Secondary Structure Prediction by Consensus Prediction from Multiple Alignments. Bioinformatics 1995, 11, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Schwede, T.; Kopp, J.; Guex, N.; Peitsch, M.C. SWISS-MODEL: An Automated Protein Homology-Modeling Server. Nucleic Acids Res. 2003, 31, 3381–3385. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Edwards, P.M. COMPUTER SOFTWARE REVIEWS Origin 7.0: Scientific Graphing and Data Analysis Software. Available online: https://0-pubs-acs-org.brum.beds.ac.uk/doi/pdf/10.1021/ci0255432 (accessed on 25 December 2022).
- Chen, Y.; Chen, Y.; Shi, C.; Huang, Z.; Zhang, Y.; Li, S.; Li, Y.; Ye, J.; Yu, C.; Li, Z. SOAPnuke: A MapReduce Acceleration-Supported Software for Integrated Quality Control and Preprocessing of High-Throughput Sequencing Data. Gigascience 2018, 7, gix120. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinform. 2011, 12, 1–16. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- He, C.; Teixeira da Silva, J.A.; Wang, H.; Si, C.; Zhang, M.; Zhang, X.; Li, M.; Tan, J.; Duan, J. Mining MYB Transcription Factors from the Genomes of Orchids (Phalaenopsis and Dendrobium) and Characterization of an Orchid R2R3-MYB Gene Involved in Water-Soluble Polysaccharide Biosynthesis. Sci. Rep. 2019, 9, 13818. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | MYB Groups | ||||
|---|---|---|---|---|---|
| MYB-related | R2R3-MYB | 3R-MYB | Atypical MYB | ||
| Eudicot | A. thaliana [5,6] | 64 | 126 | 5 | 2 |
| P. trichocarpa [25,26] | - | 192 | 5 | - | |
| O. sativa [26] | 70 | 125 | 5 | 1 | |
| Monocot | P. equestri [27] | 27 | 96 | 2 | - |
| P. aphrodite [7] | - | 99 | - | - | |
| D. officinale [7] | - | 101 | - | - | |
| C. ensifolium [28] | 27 | 102 | 2 | 2 | |
| C. goeringii [29] | - | 104 | - | - | |
| C. sinense 1 | 55 | 100 | 4 | 1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, M.; Wang, Q.; Tu, S.; Ke, S.; Bi, Y.; Ahmad, S.; Zhang, D.; Liu, D.; Lan, S. Genome-Wide Identification Analysis of the R2R3-MYB Transcription Factor Family in Cymbidium sinense for Insights into Drought Stress Responses. Int. J. Mol. Sci. 2023, 24, 3235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043235
Zhu M, Wang Q, Tu S, Ke S, Bi Y, Ahmad S, Zhang D, Liu D, Lan S. Genome-Wide Identification Analysis of the R2R3-MYB Transcription Factor Family in Cymbidium sinense for Insights into Drought Stress Responses. International Journal of Molecular Sciences. 2023; 24(4):3235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043235
Chicago/Turabian StyleZhu, Mengjia, Qianqian Wang, Song Tu, Shijie Ke, Yuanyang Bi, Sagheer Ahmad, Diyang Zhang, Dingkun Liu, and Siren Lan. 2023. "Genome-Wide Identification Analysis of the R2R3-MYB Transcription Factor Family in Cymbidium sinense for Insights into Drought Stress Responses" International Journal of Molecular Sciences 24, no. 4: 3235. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043235