
Role of Damage-Associated Molecular Patterns (DAMPS) in the Postoperative Period after Colorectal Surgery

 , ,
, ,
Abstract
:1. Introduction
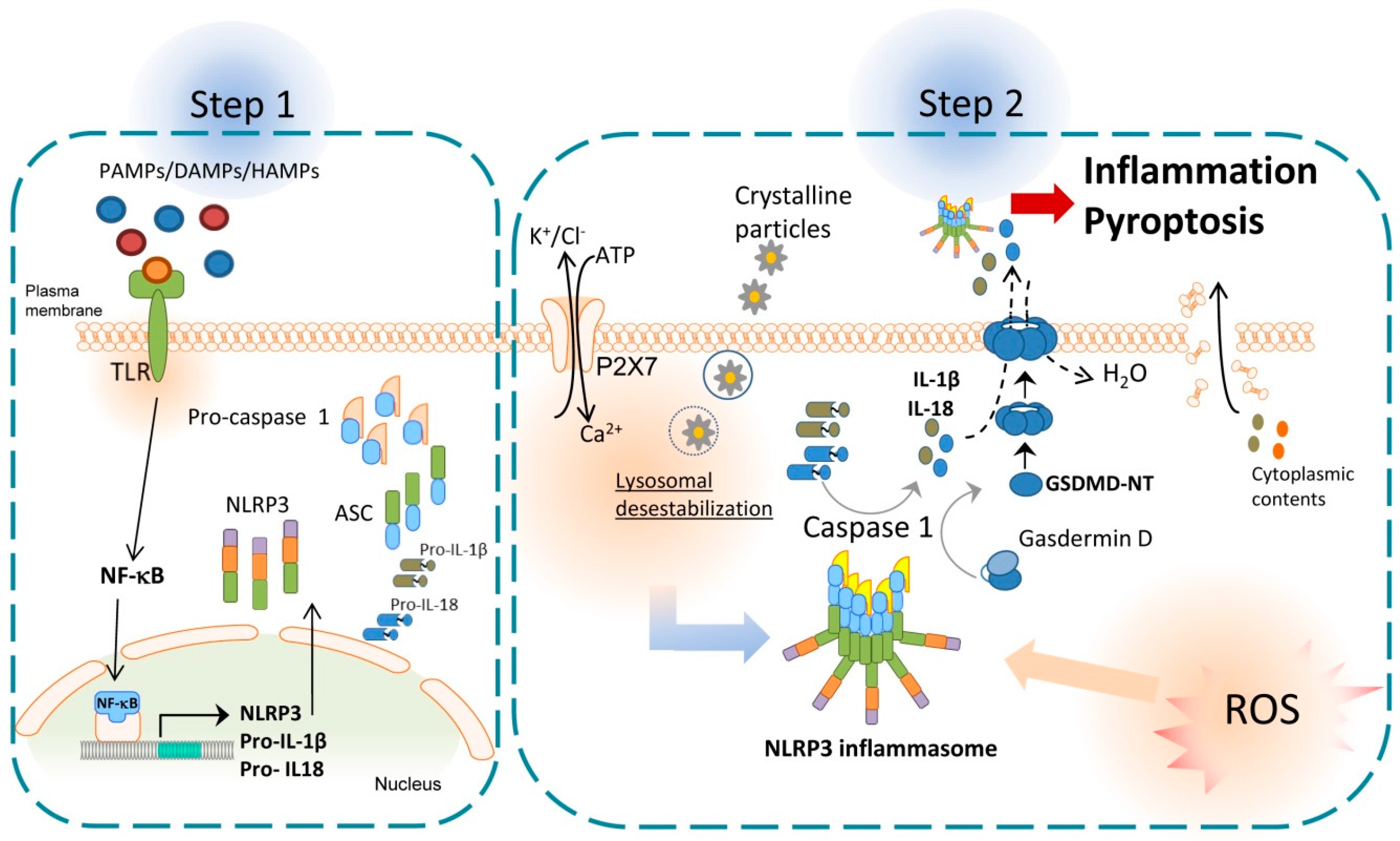
2. NLRP3 Inflammasome and Inflammation
3. Damage-Associated Molecular Patterns (DAMPs)
3.1. Types of DAMPs
3.1.1. Heat Shock Proteins (HSPs)
3.1.2. High-Mobility Group Protein Box 1 (HMGB1)
3.1.3. Adenosine Triphosphate (ATP)
3.1.4. Uric Acid Crystals
3.1.5. Extracellular DNA
3.1.6. Fibronectin
3.1.7. Free Fatty Acids

{kind=link}
{kind=link}
| Name | Origin | Cellular Function | Cellular Target That Activates the Inflammasome |
|---|---|---|---|
| Heat shock proteins (HSPs) | They are proteins that originate in stressful situations for the cell | They maintain an optimal temperature in the cell, participate in cell cycle control, and activate the immune system. | TLR4 [32,33] |
| High-mobility group protein Box 1 (HMGB1) | They are non-histone nuclear proteins that enable transcriptional regulation | Participate in DNA molecule stabilization, gene expression and activate the immune system | Binds to receptors such as RAGE, TLR4, TLR2, TLR9 that produce cytokines and activate the immune system [35] |
| ATP | These molecules are present in all cells and are indispensable for their correct functioning | It releases energy when phosphoanhydride bonds are hydrolyzed | Activates the inflammasome through the P2X4R and P2X7R receptors [32,36,37] |
| Uric Acid Crystals | Compound formed in the body in the catabolism of purines | Decreases nitric oxide, maintains the elasticity of blood vessels and promotes glucose absorption | TLR2 [38] |
| Extracellular DNA | Molecule that contains the information necessary for the development and functioning of the organism | DNA stores the information for building all cellular components | Union of ASC molecules, TLR2 and TLR4 [39,41] |
| Mitochondrial DNA | It is the circular chromosome found within the mitochondria | Encodes 5–10% of mitochondrial proteins | Union of ASC molecules, TLR2 and TLR4 [39,41] |
| Fibronectin | Glycoprotein involved in the protection of cardiac muscle | Enables the formation of fibrils in areas of tissue remodeling | TLR4 and TLR2 [42] |
| Free Fatty Acids | They come from triglycerides, when lipolysis of adipose tissue occurs | They act as a source of energy or precursors of other molecules | TLR4 [43,44] |
| Short fragments of free hyaluronan | It is a compound synthesized by the cell membrane of all cells in the organism and is located in the extracellular matrix | It is a key component for cell motility, adhesion and proliferation | TLR2 [47] |
3.1.8. Hyaluronic Acid
4. Anastomotic Leakage, Inflammation, and Fibrosis
5. Possible Role of DAMPs in Post-Surgery Complications
6. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AL | Anastomotic Leakage |
| DAMPs | Damage-Associated Molecular Patterns |
| CRP | C Reactive Protein |
| Mg/d | Milligram/day |
| HSPs | Heat Sock Proteins |
| HMGB1 | High-Mobility Group Box 1 |
| ATP | Adenosine triphosphate |
| DNA | Deoxyribonucleic acid |
| IL-1β | Interleukin 1 beta |
| IL-18 | Interleukin 18 |
| NLRs | Nucleotide-binding domain and leucine-rich repeat receptors |
| ALRs | Absent in melanoma 2-like receptors |
| ASC | Apoptosis-associated speck-like protein |
| CARD | Caspase Activation and Recruitment Domain |
| GSDMD | Gasdermin D |
| GSDMD-CT; | C-terminal fragment of Gasdermin D |
| GSDMD-NT; | N-terminal fragment of Gasdermin D |
| HSCs | Hepatic Stellate Cells |
| TLR | Toll-like receptor |
| PAMPs | Pathogen-associated molecular patterns |
| HAMPs | Homeostasis-altering molecular processes |
| NF-κB | Nuclear factor kappa B |
| K+ | Potassium |
| Cl− | Chlorine |
| Ca2+ | Calcium |
| PKR | Protein Kinase R |
| ZBP1 | Z-DNA Binding Protein 1 |
| NETs | Neutrophil Extracellular Traps |
| PRRs | Pattern Recognition Receptors |
| CLR | C-type Lectin Receptor |
| RLR | RIG-I-like receptor |
| NLR | NOD-like receptor |
| TAK1 | Transforming growth factor β-activated kinase 1 |
| TAB1 | TGF-Beta Activated Kinase 1 (MAP3K7) Binding Protein 1 |
| TAB2 | TGF-Beta Activated Kinase 1 (MAP3K7) Binding Protein 2 |
| IKK | IkappaB Kinase |
| IL-1 | Interleukin 1 |
| IL-6 | Interleukin 6 |
| IL-12 | Interleukin 12 |
| TNF-α | Tumour necrosis factor alfa |
| HSP70 | Heat Sock Proteins 70 |
| HSP90 | Heat Sock Proteins 90 |
| ABC | ATP-binding cassette |
| SGT1 | Ubiquitin ligase-associated protein |
| NACHT | Nucleotide-binding oligomerization core domain |
| LRR domain | Leucine-rich repeat |
| STAT | Signal Transducer and Activator of Transcription |
| RAGE | Receptor for Advanced Glycation End-products |
| TLR4 | Toll-like receptor 4 |
| TLR2 | Toll-like receptor 2 |
| TLR9 | Toll-like receptor 9 |
| P2X4R | The purinergic P2X4 receptor |
| P2X7R | The purinergic P2X7 receptor |
| mtDNA | Mitochondrial DNA |
| ROS | Reactive Oxygen Species |
| AIM2 | Absent in melanoma 2 |
| LDL oxidase | Low-density lipoprotein oxidase |
| FASN | Fatty acid synthase enzyme |
| RvD2 | Resolvin D2 |
| XBP1 | X-box binding protein 1 |
| STING | Stimulator of interferon gene |
| TGF-β | Transforming growth factor beta |
| µl | Microlith |
| HIPEC | Hyperthermic intraperitoneal chemotherapy |
| MOF | Multiple Organ Failure |
| SOFA | Sequential Organ Failure Assessment |
References
- Zarnescu, E.C.; Zarnescu, N.O.; Costea, R. Updates of Risk Factors for Anastomotic Leakage after Colorectal Surgery. Diagnostics 2021, 11, 2382. [Google Scholar] [CrossRef] [PubMed]
- Daams, F.; Wu, Z.; Lahaye, M.J.; Jeekel, J.; Lange, J.F. Prediction and diagnosis of colorectal anastomotic leakage: A systematic review of literature. World J. Gastrointest. Surg. 2014, 6, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Fang, A.H.; Chao, W.; Ecker, M. Review of Colonic Anastomotic Leakage and Prevention Methods. J. Clin. Med. 2020, 9, 4061. [Google Scholar] [CrossRef] [PubMed]
- Bruce, J.; Krukowski, Z.H.; Al-Khairy, G.; Russell, E.M.; Park, K.G. Systematic review of the definition and measurement of anastomotic leak after gastrointestinal surgery. Br. J. Surg. 2001, 88, 1157–1168. [Google Scholar] [CrossRef]
- Foppa, C.; Ng, S.C.; Montorsi, M.; Spinelli, A. Anastomotic leak in colorectal cancer patients: New insights and perspectives. Eur. J. Surg. Oncol. 2020, 46, 943–954. [Google Scholar] [CrossRef]
- Baeza-Murcia, M.; Valero-Navarro, G.; Pellicer-Franco, E.; Soria-Aledo, V.; Mengual-Ballester, M.; Garcia-Marin, J.A.; Betoret-Benavente, L.; Aguayo-Albasini, J.L. Early diagnosis of anastomotic leakage in colorectal surgery: Prospective observational study of the utility of inflammatory markers and determination of pathological levels. Updates Surg. 2021, 73, 2103–2111. [Google Scholar] [CrossRef]
- Bauer, C.; Duewell, P.; Lehr, H.A.; Endres, S.; Schnurr, M. Protective and aggravating effects of Nlrp3 inflammasome activation in IBD models: Influence of genetic and environmental factors. Dig. Dis. 2012, 30 (Suppl. S1), 82–90. [Google Scholar] [CrossRef] [Green Version]
- Matzinger, P. Tolerance, danger, and the extended family. Annu. Rev. Immunol. 1994, 12, 991–1045. [Google Scholar] [CrossRef]
- Pak, H.; Maghsoudi, L.H.; Soltanian, A.; Gholami, F. Surgical complications in colorectal cancer patients. Ann. Med. Surg. 2020, 55, 13–18. [Google Scholar] [CrossRef]
- Amores-Iniesta, J.; Barbera-Cremades, M.; Martinez, C.M.; Pons, J.A.; Revilla-Nuin, B.; Martinez-Alarcon, L.; Di Virgilio, F.; Parrilla, P.; Baroja-Mazo, A.; Pelegrin, P. Extracellular ATP Activates the NLRP3 Inflammasome and Is an Early Danger Signal of Skin Allograft Rejection. Cell Rep. 2017, 21, 3414–3426. [Google Scholar] [CrossRef]
- Patel, S. Danger-Associated Molecular Patterns (DAMPs): The Derivatives and Triggers of Inflammation. Curr. Allergy Asthma Rep. 2018, 18, 63. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, S.; Pelegrin, P. Pyroptosis and Redox Balance in Kidney Diseases. Antioxid. Redox Signal. 2021, 35, 40–60. [Google Scholar] [CrossRef] [PubMed]
- De Miguel, C.; Pelegrin, P.; Baroja-Mazo, A.; Cuevas, S. Emerging Role of the Inflammasome and Pyroptosis in Hypertension. Int. J. Mol. Sci. 2021, 22, 1064. [Google Scholar] [CrossRef]
- Xiao, L.; Magupalli, V.G.; Wu, H. Cryo-EM structures of the active NLRP3 inflammasome disc. Nature 2023, 613, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Tuipulotu, D.E.; Tan, W.H.; Kay, C.; Man, S.M. Emerging Activators and Regulators of Inflammasomes and Pyroptosis. Trends Immunol. 2019, 40, 1035–1052. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Kanneganti, T.D. Converging roles of caspases in inflammasome activation, cell death and innate immunity. Nat. Rev. Immunol. 2016, 16, 7–21. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, F.I.; Lu, A.; Chen, J.W.; Ruan, J.; Tang, C.; Wu, H.; Ploegh, H.L. A single domain antibody fragment that recognizes the adaptor ASC defines the role of ASC domains in inflammasome assembly. J. Exp. Med. 2016, 213, 771–790. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Lucas-Ruiz, F.; Penin-Franch, A.; Pons, J.A.; Ramirez, P.; Pelegrin, P.; Cuevas, S.; Baroja-Mazo, A. Emerging Role of NLRP3 Inflammasome and Pyroptosis in Liver Transplantation. Int. J. Mol. Sci. 2022, 23, 14396. [Google Scholar] [CrossRef]
- McKee, C.M.; Coll, R.C. NLRP3 inflammasome priming: A riddle wrapped in a mystery inside an enigma. J. Leukoc. Biol. 2020, 108, 937–952. [Google Scholar] [CrossRef] [PubMed]
- Paik, S.; Kim, J.K.; Silwal, P.; Sasakawa, C.; Jo, E.K. An update on the regulatory mechanisms of NLRP3 inflammasome activation. Cell. Mol. Immunol. 2021, 18, 1141–1160. [Google Scholar] [CrossRef]
- Coll, R.C.; Schroder, K.; Pelegrin, P. NLRP3 and pyroptosis blockers for treating inflammatory diseases. Trends Pharmacol. Sci. 2022, 43, 653–668. [Google Scholar] [CrossRef]
- Saeedi-Boroujeni, A.; Nashibi, R.; Ghadiri, A.A.; Nakajima, M.; Salmanzadeh, S.; Mahmoudian-Sani, M.R.; Hanafi, M.G.; Sharhani, A.; Khodadadi, A. Tranilast as an Adjunctive Therapy in Hospitalized Patients with Severe COVID- 19: A Randomized Controlled Trial. Arch. Med. Res. 2022, 53, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Faghfouri, A.H.; Baradaran, B.; Khabbazi, A.; Shadbad, M.A.; Papi, S.; Faghfuri, E.; Khajebishak, Y.; Rahmani, S.; Hayat, P.T.; Alipour, B. Regulation of NLRP3 inflammasome by zinc supplementation in Behcet’s disease patients: A double-blind, randomized placebo-controlled clinical trial. Int. Immunopharmacol. 2022, 109, 108825. [Google Scholar] [CrossRef]
- Murao, A.; Aziz, M.; Wang, H.; Brenner, M.; Wang, P. Release mechanisms of major DAMPs. Apoptosis 2021, 26, 152–162. [Google Scholar] [CrossRef]
- Torres Pedraza, S.; Betancur, J.G.; Urcuqui-inchima, S. Viral recognition by the innate immune system: The role of pattern recognition receptors. Colombino Med. 2010, 41, 11. [Google Scholar] [CrossRef]
- Sundaram, B.; Kanneganti, T.D. Advances in Understanding Activation and Function of the NLRC4 Inflammasome. Int. J. Mol. Sci. 2021, 22, 1048. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Olarreaga, I.O.; Alberdi, L.G.; Navalon, J.A.A. El sistema inmune y su función en el sistema nervioso central. Revista de la Fac. De Cienc. De La Salud De La Univ. Alfonso X El Sabio 2014, 11, 1–21. [Google Scholar]
- De Maio, A.; Vazquez, D. Extracellular heat shock proteins: A new location, a new function. Shock 2013, 40, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Suarez, R.; Buelvas, N. Inflammasome: Activation mechanisms. Invest. Clin. 2015, 56, 74–99. [Google Scholar] [PubMed]
- Galliussi, G. Mecanismos antiinflamatorios de los nitroalquenos: Regulación del inflamasoma NLRP3. Tesina Grado Univ. República 2015, 74, 1. [Google Scholar]
- Chen, R.; Kang, R.; Tang, D. The mechanism of HMGB1 secretion and release. Exp. Mol. Med. 2022, 54, 91–102. [Google Scholar] [CrossRef]
- Rivera-Pérez, R.J.; Sevilla-González, M.d.l.L.; Flores-Estrada, J.J. Estrategias para inhibir a HMGB1 liberado durante la sepsis. Rev. Hosp. Jua. Mex. 2016, 83, 6. [Google Scholar]
- Sandall, C.F.; Ziehr, B.K.; MacDonald, J.A. ATP-Binding and Hydrolysis in Inflammasome Activation. Molecules 2020, 25, 4572. [Google Scholar] [CrossRef]
- Silverman, W.R.; de Rivero Vaccari, J.P.; Locovei, S.; Qiu, F.; Carlsson, S.K.; Scemes, E.; Keane, R.W.; Dahl, G. The pannexin 1 channel activates the inflammasome in neurons and astrocytes. J. Biol. Chem. 2009, 284, 18143–18151. [Google Scholar] [CrossRef] [Green Version]
- Ghaemi-Oskouie, F.; Shi, Y. The role of uric acid as an endogenous danger signal in immunity and inflammation. Curr. Rheumatol. Rep. 2011, 13, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Shimada, K.; Crother, T.R.; Karlin, J.; Dagvadorj, J.; Chiba, N.; Chen, S.; Ramanujan, V.K.; Wolf, A.J.; Vergnes, L.; Ojcius, D.M.; et al. Oxidized mitochondrial DNA activates the NLRP3 inflammasome during apoptosis. Immunity 2012, 36, 401–414. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Liang, S.; Sanchez-Lopez, E.; He, F.; Shalapour, S.; Lin, X.J.; Wong, J.; Ding, S.; Seki, E.; Schnabl, B.; et al. New mitochondrial DNA synthesis enables NLRP3 inflammasome activation. Nature 2018, 560, 198–203. [Google Scholar] [CrossRef]
- Lupfer, C.R.; Rodriguez, A.; Kanneganti, T.D. Inflammasome activation by nucleic acids and nucleosomes in sterile inflammation... or is it sterile? FEBS J. 2017, 284, 2363–2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, C.; Zhang, X.; Hu, M.; Teng, T.; Yuan, Y.P.; Song, P.; Kong, C.Y.; Xu, S.C.; Ma, Z.G.; Tang, Q.Z. Fibronectin type III domain-containing 5 improves aging-related cardiac dysfunction in mice. Aging Cell 2022, 21, e13556. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.S.; Lee, S.; Park, M.A.; Siempos, I.I.; Haslip, M.; Lee, P.J.; Yun, M.; Kim, C.K.; Howrylak, J.; Ryter, S.W.; et al. UCP2-induced fatty acid synthase promotes NLRP3 inflammasome activation during sepsis. J. Clin. Invest. 2015, 125, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, T.; Takahashi, M. Saturated fatty acid-crystals activate NLRP3 inflammasome. Aging (Albany NY) 2019, 11, 1613–1614. [Google Scholar] [CrossRef]
- Wang, Y.H.; Li, Y.; Wang, J.N.; Zhao, Q.X.; Jin, J.; Wen, S.; Wang, S.C.; Sun, T. Maresin 1 Attenuates Radicular Pain Through the Inhibition of NLRP3 Inflammasome-Induced Pyroptosis via NF-kappaB Signaling. Front. Neurosci. 2020, 14, 831. [Google Scholar] [CrossRef]
- Cao, L.; Wang, Y.; Wang, Y.; Lv, F.; Liu, L.; Li, Z. Resolvin D2 suppresses NLRP3 inflammasome by promoting autophagy in macrophages. Exp. Ther. Med. 2021, 22, 1222. [Google Scholar] [CrossRef]
- Feng, F.; Li, Z.; Potts-Kant, E.N.; Wu, Y.; Foster, W.M.; Williams, K.L.; Hollingsworth, J.W. Hyaluronan activation of the Nlrp3 inflammasome contributes to the development of airway hyperresponsiveness. Environ. Health Perspect. 2012, 120, 1692–1698. [Google Scholar] [CrossRef]
- Leijte, G.P.; Custers, H.; Gerretsen, J.; Heijne, A.; Roth, J.; Vogl, T.; Scheffer, G.J.; Pickkers, P.; Kox, M. Increased Plasma Levels of Danger-Associated Molecular Patterns Are Associated With Immune Suppression and Postoperative Infections in Patients Undergoing Cytoreductive Surgery and Hyperthermic Intraperitoneal Chemotherapy. Front. Immunol. 2018, 9, 663. [Google Scholar] [CrossRef]
- El Zaher, H.A.; Ghareeb, W.M.; Fouad, A.M.; Madbouly, K.; Fathy, H.; Vedin, T.; Edelhamre, M.; Emile, S.H.; Faisal, M. Role of the triad of procalcitonin, C-reactive protein, and white blood cell count in the prediction of anastomotic leak following colorectal resections. World J. Surg. Oncol. 2022, 20, 33. [Google Scholar] [CrossRef]
- Lee, S.B.; Kalluri, R. Mechanistic connection between inflammation and fibrosis. Kidney International. Suppl. 2010, 78, S22–S26. [Google Scholar] [CrossRef] [Green Version]
- Lara-de-la-Fuente, R.; Alanis-Cruces, J.M. Postoperative fibrosis after lumbar surgery. Acta Ortop. Mex. 2009, 23, 90–93. [Google Scholar] [PubMed]
- Kersey, J.P.; Vivian, A.J. Mitomycin and amniotic membrane: A new method of reducing adhesions and fibrosis in strabismus surgery. Strabismus 2008, 16, 116–118. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhou, H.F.; Tong, M.; Chen, L.; Ren, K.; Zhao, G.J. Colchicine inhibits endothelial inflammation via NLRP3/CRP pathway. Int. J. Cardiol. 2019, 294, 55. [Google Scholar] [CrossRef] [PubMed]
- Straatman, J.; Cuesta, M.A.; Tuynman, J.B.; Veenhof, A.; Bemelman, W.A.; van der Peet, D.L. C-reactive protein in predicting major postoperative complications are there differences in open and minimally invasive colorectal surgery? Substudy from a randomized clinical trial. Surg. Endosc. 2018, 32, 2877–2885. [Google Scholar] [CrossRef] [Green Version]
- Straatman, J.; Harmsen, A.M.; Cuesta, M.A.; Berkhof, J.; Jansma, E.P.; van der Peet, D.L. Predictive Value of C-Reactive Protein for Major Complications after Major Abdominal Surgery: A Systematic Review and Pooled-Analysis. PLoS ONE 2015, 10, e0132995. [Google Scholar] [CrossRef] [Green Version]
- Zajonz, T.; Koch, C.; Schwiddessen, J.; Markmann, M.; Hecker, M.; Edinger, F.; Schmidt, G.; Boening, A.; Sander, M.; Schneck, E. Minimized Extracorporeal Circulation Is Associated with Reduced Plasma Levels of Free-Circulating Mitochondrial DNA Compared to Conventional Cardiopulmonary Bypass: A Secondary Analysis of an Exploratory, Prospective, Interventional Study. J. Clin. Med. 2022, 11, 2994. [Google Scholar] [CrossRef]
- Pencovich, N.; Nevo, N.; Weiser, R.; Bonder, E.; Bogoch, Y.; Nachmany, I. Postoperative Rise of Circulating Mitochondrial DNA Is Associated with Inflammatory Response in Patients following Pancreaticoduodenectomy. Eur. Surg. Research. Eur. Chir. Forschung. Rech. Chir. Eur. 2021, 62, 18–24. [Google Scholar] [CrossRef]
- Grigoriev, E.; Ponasenko, A.V.; Sinitskaya, A.V.; Ivkin, A.A.; Kornelyuk, R.A. Mitochondrial DNA as a Candidate Marker of Multiple Organ Failure after Cardiac Surgery. Int. J. Mol. Sci. 2022, 23, 14748. [Google Scholar] [CrossRef]
- Yoshino, O.; Wong, B.K.L.; Cox, D.R.A.; Lee, E.; Hepworth, G.; Christophi, C.; Jones, R.; Dobrovic, A.; Muralidharan, V.; Perini, M.V. Elevated levels of circulating mitochondrial DNA predict early allograft dysfunction in patients following liver transplantation. J. Gastroenterol. Hepatol. 2021, 36, 3500–3507. [Google Scholar] [CrossRef]
- Doukas, P.; Hellfritsch, G.; Krabbe, H.; Frankort, J.; Jacobs, M.J.; Gombert, A.; Simon, F. Onset of Adverse Abdominal Events Due to Intestinal Ischemia-Reperfusion Injury after Aortic Cross-Clamping Is Associated with Elevated HSP70 Serum Levels in the Early Postoperative Phase. Int. J. Mol. Sci. 2022, 23, 15063. [Google Scholar] [CrossRef]
- Xiao, L.; Zhang, Z.; Liu, J.; Zheng, Z.; Xiong, Y.; Li, C.; Feng, Y.; Yin, S. HMGB1 accumulation in cytoplasm mediates noise-induced cochlear damage. Cell Tissue Res. 2022, 391, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Sun, P.; Chen, Y.; Jiang, X.; Zhang, C.; Qu, B.; Meng, X. NLRP3 inflammasome activation contributes to the cognitive decline after cardiac surgery. Front. Surg. 2022, 9, 992769. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caballero-Herrero, M.J.; Jumilla, E.; Buitrago-Ruiz, M.; Valero-Navarro, G.; Cuevas, S. Role of Damage-Associated Molecular Patterns (DAMPS) in the Postoperative Period after Colorectal Surgery. Int. J. Mol. Sci. 2023, 24, 3862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043862
Caballero-Herrero MJ, Jumilla E, Buitrago-Ruiz M, Valero-Navarro G, Cuevas S. Role of Damage-Associated Molecular Patterns (DAMPS) in the Postoperative Period after Colorectal Surgery. International Journal of Molecular Sciences. 2023; 24(4):3862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043862
Chicago/Turabian StyleCaballero-Herrero, María José, Esther Jumilla, Manuel Buitrago-Ruiz, Graciela Valero-Navarro, and Santiago Cuevas. 2023. "Role of Damage-Associated Molecular Patterns (DAMPS) in the Postoperative Period after Colorectal Surgery" International Journal of Molecular Sciences 24, no. 4: 3862. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24043862