
ETC-159, an Upstream Wnt inhibitor, Induces Tumour Necrosis via Modulation of Angiogenesis in Osteosarcoma

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Clinical Osteosarcoma Samples Demonstrate High WLS and β-Catenin Expression
2.2. Effect of ETC-159, a Novel PORCN Inhibitor, on Wnt Signalling in Osteosarcoma
2.3. ETC-159 Decreases Angiogenesis in Osteosarcoma
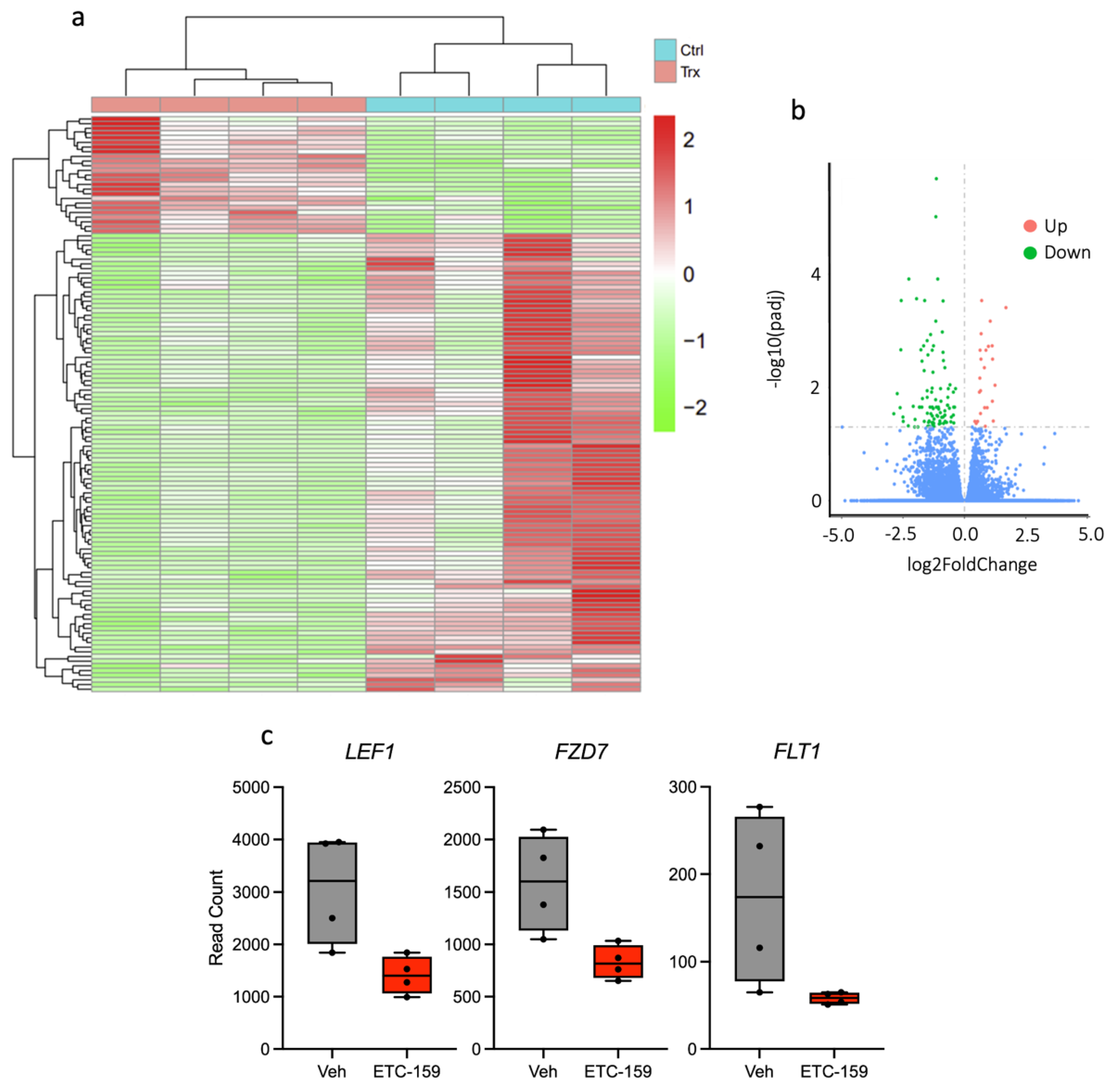
2.4. RNAseq of Osteosarcoma Xenograft Tissue
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. Isolation and Characterization of New Osteosarcoma Cell Lines
4.3. Establishment of Human Tumor Xenograft in Immunodeficient Mice
4.4. Examination of Osteogenic Markers in Patient Samples
4.5. Immunoblot Analysis of Cell Growth Markers
4.6. Chick Chorioallantoic Membrane (CAM) Assay
4.7. RNA Sequencing and Data Analysis
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Isakoff, M.S.; Bielack, S.S.; Meltzer, P.; Gorlick, R. Osteosarcoma: Current Treatment and a Collaborative Pathway to Success. J. Clin. Oncol. 2015, 33, 3029–3035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kager, L.; Tamamyan, G.; Bielack, S. Novel insights and therapeutic interventions for pediatric osteosarcoma. Future Oncol. 2017, 13, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Bielack, S.S.; Kempf-Bielack, B.; Branscheid, D.; Carrle, D.; Friedel, G.; Helmke, K.; Kevric, M.; Jundt, G.; Kühne, T.; Maas, R.; et al. Second and Subsequent Recurrences of Osteosarcoma: Presentation, Treatment, and Outcomes of 249 Consecutive Cooperative Osteosarcoma Study Group Patients. J. Clin. Oncol. 2008, 27, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Kager, L.; Zoubek, A.; Pötschger, U.; Kastner, U.; Flege, S. Primary Metastatic Osteosarcoma: Presentation and Outcome of Patients Treated on Neoadjuvant Cooperative Osteosarcoma Study Group Protocols. J. Clin. Oncol. 2003, 21, 2011–2018. [Google Scholar] [CrossRef] [PubMed]
- Kempf-Bielack, B.; Bielack, S.S.; Jürgens, H.; Branscheid, D.; Berdel, W.E. Exner, G.U.; Göbel, U.; Helmke, K.; Jundt, G.; Kabisch, H; et al. Osteosarcoma Relapse After Combined Modality Therapy: An Analysis of Unselected Patients in the Cooperative Osteosarcoma Study Group (COSS). J. Clin. Oncol. 2004, 23, 559–568. [Google Scholar] [CrossRef]
- Cleton-Jansen, A.-M.; Anninga, J.K.; Brujin, I.H.B.; Romeo, S.; Oosting, J.; Egeler, R.M.; Gelderblom, H.; Taminiau, A.H.M.; Hogendoorn, P.C.W. Profiling of high-grade central osteosarcoma and its putative progenitor cells identifies tumourigenic pathways. Brit. J. Cancer 2009, 101, 1909–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McQueen, P.; Ghaffar, S.; Guo, Y.; Rubin, E.M.; Zi, X.; Hoang, B.H. The Wnt signaling pathway: Implications for therapy in osteosarcoma. Expert Rev. Anticanc. 2014, 11, 1223–1232. [Google Scholar] [CrossRef]
- Yu, J.; Virshup, D.M. Updating the Wnt pathways. Biosci. Rep. 2014, 34, e00142. [Google Scholar] [CrossRef] [Green Version]
- Alok, A.; Lei, Z.; Jagannathan, N.S.; Kaur, S.; Harmston, N.; Rozen, S.G.; Tucker-Kellog, L.; Virshup, D.M. Wnt proteins synergize to activate β-catenin signaling. J. Cell Sci. 2017, 130, 1532–1544. [Google Scholar] [CrossRef]
- Hu, J.; Hu, W.-X. Targeting signaling pathways in multiple myeloma: Pathogenesis and implication for treatments. Cancer Lett. 2018, 414, 214–221. [Google Scholar] [CrossRef]
- Huang, F.; Chen, J.; Wang, Z.; Lan, R.; Fu, L.; Zhang, L. δ-Catenin promotes tumorigenesis and metastasis of lung adenocarcinoma. Oncol. Rep. 2018, 39, 809–817. [Google Scholar] [CrossRef]
- Rahmani, F.; Avan, A.; Hashemy, S.I.; Hassanian, S.M. Role of Wnt/β-catenin signaling regulatory microRNAs in the pathogenesis of colorectal cancer. J. Cell Physiol. 2018, 233, 811–817. [Google Scholar] [CrossRef]
- Jin, H.; Luo, S.; Wang, Y.; Liu, C.; Piao, Z.; Xu, M.; Guan, W.; Li, Q.; Zou, H.; Tan, Q.; et al. miR-135b Stimulates Osteosarcoma Recurrence and Lung Metastasis via Notch and Wnt/β-Catenin Signaling. Mol. Ther.—Nucleic Acids 2017, 8, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Martins-Neves, S.R.; Paiva-Oliveria, D.I.; Fontes-Ribeiro, C.; Bovée, J.V.M.G.; Cleton-Jansen, A. IWR-1, a tankyrase inhibitor, attenuates Wnt/β-catenin signaling in cancer stem-like cells and inhibits in vivo the growth of a subcutaneous human osteosarcoma xenograft. Cancer Lett. 2018, 414, 1–15. [Google Scholar] [CrossRef]
- Vega, O.A.; Lucero, C.M.J.; Araya, H.F.; Jerez, S.; Tapia, J.C.; Antonelli, M.; Salazar-Onfray, F.; Heras, F.L.; Thaler, R.; Riester, S.M.; et al. Wnt/β-Catenin Signaling Activates Expression of the Bone-Related Transcription Factor RUNX2 in Select Human Osteosarcoma Cell Types. J. Cell Biochem. 2017, 118, 3662–3674. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, D.; Han, S.; Gao, P.; Liu, C.; Li, J.; Pan, X. Fibulin-3 promotes osteosarcoma invasion and metastasis by inducing epithelial to mesenchymal transition and activating the Wnt/β-catenin signaling pathway. Sci. Rep. 2017, 7, 6215. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Zhu, J.; Liu, F.; Liu, J.; Li, M. Downregulation of 14-3-3β inhibits proliferation and migration in osteosarcoma cells. Mol. Med. Rep. 2018, 17, 2493–2500. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, A.M.; D’Amore, P.A. Wnt signaling in the vasculature. Angiogenesis 2002, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, A.M.; Sullivan, K.M.; D’Amore, P.A. Cultured endothelial cells display endogenous activation of the canonical Wnt signaling pathway and express multiple ligands, receptors, and secreted modulators of Wnt signaling. Dev. Dynam. 2006, 235, 3110–3120. [Google Scholar] [CrossRef] [PubMed]
- Pate, K.T.; Stringari, C.; Sprowl-Tanio, S.; Wang, K.; TeSlaa, T.; Hoverter, N.P.; McQuade, M.M.; Garner, C.; Digman, M.A.; Teitell, M.A.; et al. Wnt signaling directs a metabolic program of glycolysis and angiogenesis in colon cancer. EMBO J. 2014, 33, 1454–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Easwaran, V.; Lee, S.H.; Inge, L.; Guo, L.; Goldbeck, C.; Garrett, E.; Wiesmann, M.; Garcia, P.D.; Fuller, J.H.; Chan, V.; et al. β-Catenin Regulates Vascular Endothelial Growth Factor Expression in Colon Cancer. Cancer Res. 2003, 63, 3145–3153. [Google Scholar] [PubMed]
- Lammli, J.; Fan, M.; Rosenthal, H.G.; Patni, M.; Rinehart, E.; Vergara, G.; Ablah, E.; Wooley, P.H.; Lucas, G.; Yang, S.-Y. Expression of Vascular Endothelial Growth Factor correlates with the advance of clinical osteosarcoma. Int. Orthop. 2012, 36, 2307–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, N.; Yano, H.; Nishida, T.; Kamura, T.; Kojiro, M. Angiogenesis in Cancer. Vasc. Health Risk Manag. 2006, 2, 213–219. [Google Scholar] [CrossRef]
- Folkman, J. Angiogenesis. Medicine 2006, 57, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; VanCleave, A.; Helmuth, R.; Torres, H.; Rickel, K.; Wollenzien, H.; Sun, H.; Zeng, E.; Zhao, J.; Tao, J. Targeting the Wnt/β-catenin pathway in human osteosarcoma cells. Oncotarget 2018, 9, 36780–36792. [Google Scholar] [CrossRef] [Green Version]
- Ottaviano, L.; Schaefer, K.; Gajewski, M.; Huckenbeck, W.; Baldus, S.; Rogel, U.; Mackintosh, C.; de Alava, E.; Myklebost, O.; Kresse, S.H.; et al. Molecular characterization of commonly used cell lines for bone tumor research: A trans-European EuroBoNet effort. Genes Chromosom. Cancer 2010, 49, 40–51. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, J.; Sun, M.; Zuo, D.; Wang, H.; Shen, J.; Jiang, W.; Mu, H.; Ma, X.; Yin, F.; et al. Multi-omics analysis identifies osteosarcoma subtypes with distinct prognosis indicating stratified treatment. Nat. Commun. 2022, 13, 7207. [Google Scholar] [CrossRef] [PubMed]
- Coombs, G.S.; Yu, J.; Canning, C.A.; Veltri, C.A.; Covey, T.M.; Cheong, J.K.; Utomo, V.; Banerjee, N.; Zhang, Z.H.; Jadulco, R.C.; et al. WLS-dependent secretion of WNT3A requires Ser209 acylation and vacuolar acidification. J. Cell Sci. 2010, 123, 3357–3367. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Chia, J.; Canning, C.A.; Jones, C.M.; Bard, F.A.; Virshup, D.M. WLS Retrograde Transport to the Endoplasmic Reticulum during Wnt Secretion. Dev. Cell 2014, 29, 277–291. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhao, M.; Tian, A.; Zhang, X.; Yao, Z.; Ma, X. Aberrant activation of Wnt/β-catenin signaling drives proliferation of bone sarcoma cells. Oncotarget 2015, 6, 17570–17583. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Mohseny, A.B.; Karperien, M.; Hogendoorn, P.C.; Zhou, G.; Cleton-Jansen, A. Inactive Wnt/β-catenin pathway in conventional high-grade osteosarcoma. J. Pathol. 2010, 220, 24–33. [Google Scholar] [CrossRef]
- Bänziger, C.; Soldini, D.; Schütt, C.; Zipperlen, P.; Hausmann, G.; Basler, K. Wntless, a Conserved Membrane Protein Dedicated to the Secretion of Wnt Proteins from Signaling Cells. Cell 2006, 125, 509–522. [Google Scholar] [CrossRef] [Green Version]
- Hoang, B.H.; Kubo, T.; Healey, J.H.; Yang, R.; Nathan, S.S.; Kolb, E.A.; Mazza, B.; Meyers, P.A.; Gorlick, R. Dickkopf 3 Inhibits Invasion and Motility of Saos-2 Osteosarcoma Cells by Modulating the Wnt-β-Catenin Pathway. Cancer Res. 2004, 64, 2734–2739. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Fallen, S.; Abaan, H.Ö.; Hayran, M.; Gonzalez, C.; Wodajo, F.; MacDonald, T.; Toretsky, J.A.; Üren, A. Wnt10b induces chemotaxis of osteosarcoma and correlates with reduced survival. Pediatr. Blood Cancer 2008, 51, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Madan, B.; Ke, Z.; Harmston, N.; Ho, S.Y.; Frois, A.O.; Alam, J.; Jeyaraj, D.A.; Pendharkar, V.; Ghosh, K.; Virshup, I.H.; et al. Wnt addiction of genetically defined cancers reversed by PORCN inhibition. Oncogene 2016, 35, 2197–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komekado, H.; Yamamoto, H.; Chiba, T.; Kikuchi, A. Glycosylation and palmitoylation of Wnt-3a are coupled to produce an active form of Wnt-3a. Genes Cells 2007, 12, 521–534. [Google Scholar] [CrossRef] [Green Version]
- Galli, L.M.; Barnes, T.L.; Secrest, S.S.; Kadowaki, T.; Burrus, L.W. Porcupine-mediated lipid-modification regulates the activity and distribution of Wnt proteins in the chick neural tube. Development 2007, 134, 3339–3348. [Google Scholar] [CrossRef] [Green Version]
- Takada, R.; Satomi, Y.; Kurata, T.; Ueno, N.; Norioka, S.; Kondoh, H.; Takao, T.; Takada, S. Monounsaturated Fatty Acid Modification of Wnt Protein: Its Role in Wnt Secretion. Dev. Cell 2006, 11, 791–801. [Google Scholar] [CrossRef] [Green Version]
- Schiavone, K.; Garnier, D.; Heymann, M.-F.; Heymann, D. Stem Cells Heterogeneity in Cancer. Adv. Exp. Med. Biol. 2019, 1139, 187–200. [Google Scholar] [PubMed]
- Aberle, H.; Bauer, A.; Stappert, J.; Kispert, A.; Kemler, R. β-catenin is a target for the ubiquitin–proteasome pathway. EMBO J. 1997, 16, 3797–3804. [Google Scholar] [CrossRef] [Green Version]
- Stamos, J.L.; Weis, W.I. The β-Catenin Destruction Complex. Cold Spring Harb. Perspect. Biol. 2013, 5, a007898. [Google Scholar] [CrossRef]
- Toné, S.; Sugimoto, K.; Tanda, K.; Suda, T.; Uehira, K.; Kanouchi, H.; Samejima, K.; Minatogawa, Y.; Earnshaw, W.C. Three distinct stages of apoptotic nuclear condensation revealed by time-lapse imaging, biochemical and electron microscopy analysis of cell-free apoptosis. Exp. Cell Res. 2007, 313, 3635–3644. [Google Scholar] [CrossRef] [Green Version]
- Gown, A.M.; Willingham, M.C. Improved Detection of Apoptotic Cells in Archival Paraffin Sections: Immunohistochemistry Using Antibodies to Cleaved Caspase 3. J. Histochem. Cytochem. 2001, 50, 449–454. [Google Scholar] [CrossRef] [Green Version]
- Crowley, L.C.; Waterhouse, N.J. Detecting Cleaved Caspase-3 in Apoptotic Cells by Flow Cytometry. Cold Spring Harb. Perspect. Biol. 2016, 2016, pdb.prot087312. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Wang, D.; Guo, P.; Zhang, Z.; Feng, J. Apatinib Functioned as Tumor Suppressor of Synovial Sarcoma through Regulating miR-34a-5p/HOXA13 Axis. Comput. Math. Method Med. 2022, 2022, 7214904. [Google Scholar] [CrossRef]
- Sherwood, V. WNT Signaling: An Emerging Mediator of Cancer Cell Metabolism? Mol. Cell Biol. 2015, 35, 2–10. [Google Scholar] [CrossRef] [Green Version]
- Gately, S. The Contributions of Cyclooxygenase-2 to Tumor Angiogenesis. Cancer Metastasis Rev. 2000, 19, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, N. Role of vascular endothelial growth factor in regulation of physiological angiogenesis. J. Physiol. -Cell Physiol. 2001, 280, C1358–C1366. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Dong, A.; Fernandez-Ruiz, V.; Shan, J.; Kawa, M.; Martínez-Ansó, E.; Prieto, J.; Qian, C. Blockade of Wnt Signaling Inhibits Angiogenesis and Tumor Growth in Hepatocellular Carcinoma. Cancer Res. 2009, 69, 6951–6959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.; Ng, M.; Subbiah, V.; Messersmith, W.; Teneggi, V.; Diermayr, V.; Ethirajulu, K.; Yeo, P.; Gan, B.H.; Lee, L.H.; et al. 71O Phase I extension study of ETC-159 an oral PORCN inhibitor administered with bone protective treatment, in patients with advanced solid tumours. Ann. Oncol. 2018, 29, ix23–ix24. [Google Scholar]
- Madan, B.; McDonald, M.J.; Foxa, G.E.; Diegel, C.R.; Williams, B.O.; Virshup, D.M. Bone loss from Wnt inhibition mitigated by concurrent alendronate therapy. Bone Res. 2018, 6, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, B.P.; Zhou, Y.; Gupta, A.; Leong, D.T.; Aung, K.Z.; Ling, L.; Pho, R.W.H.; Galindo, M.; Salto-Tellez, M.; Stein, G.S.; et al. Runx2, p53, and pRB status as diagnostic parameters for deregulation of osteoblast growth and differentiation in a new pre-chemotherapeutic osteosarcoma cell line (OS1). J. Cell. Physiol. 2009, 221, 778–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chua, K.; Sim, A.Y.L.; Yeo, E.Y.M.; Bin Masroni, M.S.; Naw, W.W.; Leong, S.M.; Lee, K.W.; Lim, H.J.; Virshup, D.M.; Lee, V.K.M. ETC-159, an Upstream Wnt inhibitor, Induces Tumour Necrosis via Modulation of Angiogenesis in Osteosarcoma. Int. J. Mol. Sci. 2023, 24, 4759. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24054759
Chua K, Sim AYL, Yeo EYM, Bin Masroni MS, Naw WW, Leong SM, Lee KW, Lim HJ, Virshup DM, Lee VKM. ETC-159, an Upstream Wnt inhibitor, Induces Tumour Necrosis via Modulation of Angiogenesis in Osteosarcoma. International Journal of Molecular Sciences. 2023; 24(5):4759. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24054759
Chicago/Turabian StyleChua, Kenon, Arthur Yi Loong Sim, Eric Yew Meng Yeo, Muhammad Sufyan Bin Masroni, Wah Wah Naw, Sai Mun Leong, Kee Wah Lee, Huey Jin Lim, David M. Virshup, and Victor Kwan Min Lee. 2023. "ETC-159, an Upstream Wnt inhibitor, Induces Tumour Necrosis via Modulation of Angiogenesis in Osteosarcoma" International Journal of Molecular Sciences 24, no. 5: 4759. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24054759