
Differential Transcriptome Responses in Human THP-1 Macrophages Following Exposure to T98G and LN-18 Human Glioblastoma Secretions: A Simplified Bioinformatics Approach to Understanding Patient-Glioma-Specific Effects on Tumor-Associated Macrophages

Abstract
:1. Introduction
2. Results
2.1. THP-1 Cells Treated with PMA Show Differences in Morphology Compared to Untreated THP-1 Cells
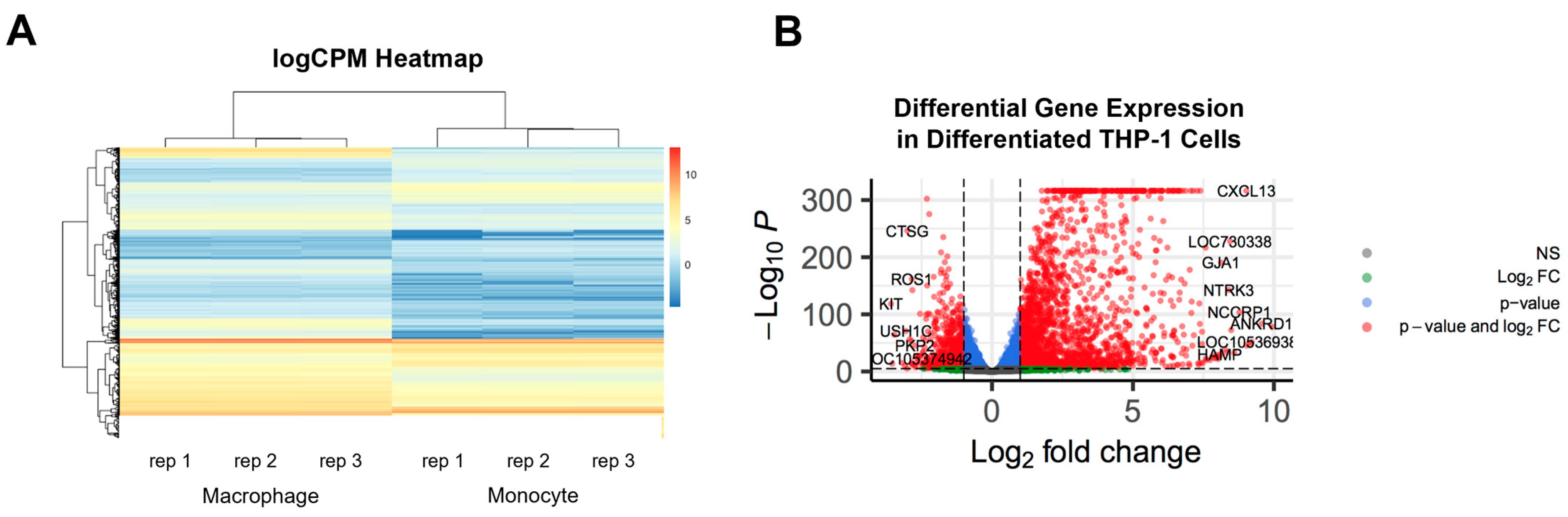
2.2. PMA-Differentiated THP-1 Cells Display Distinct Genotypic Profile Compared to Undifferentiated THP-1 Monocytes and Have Positively Correlated logCPM Values among Biological Replicates
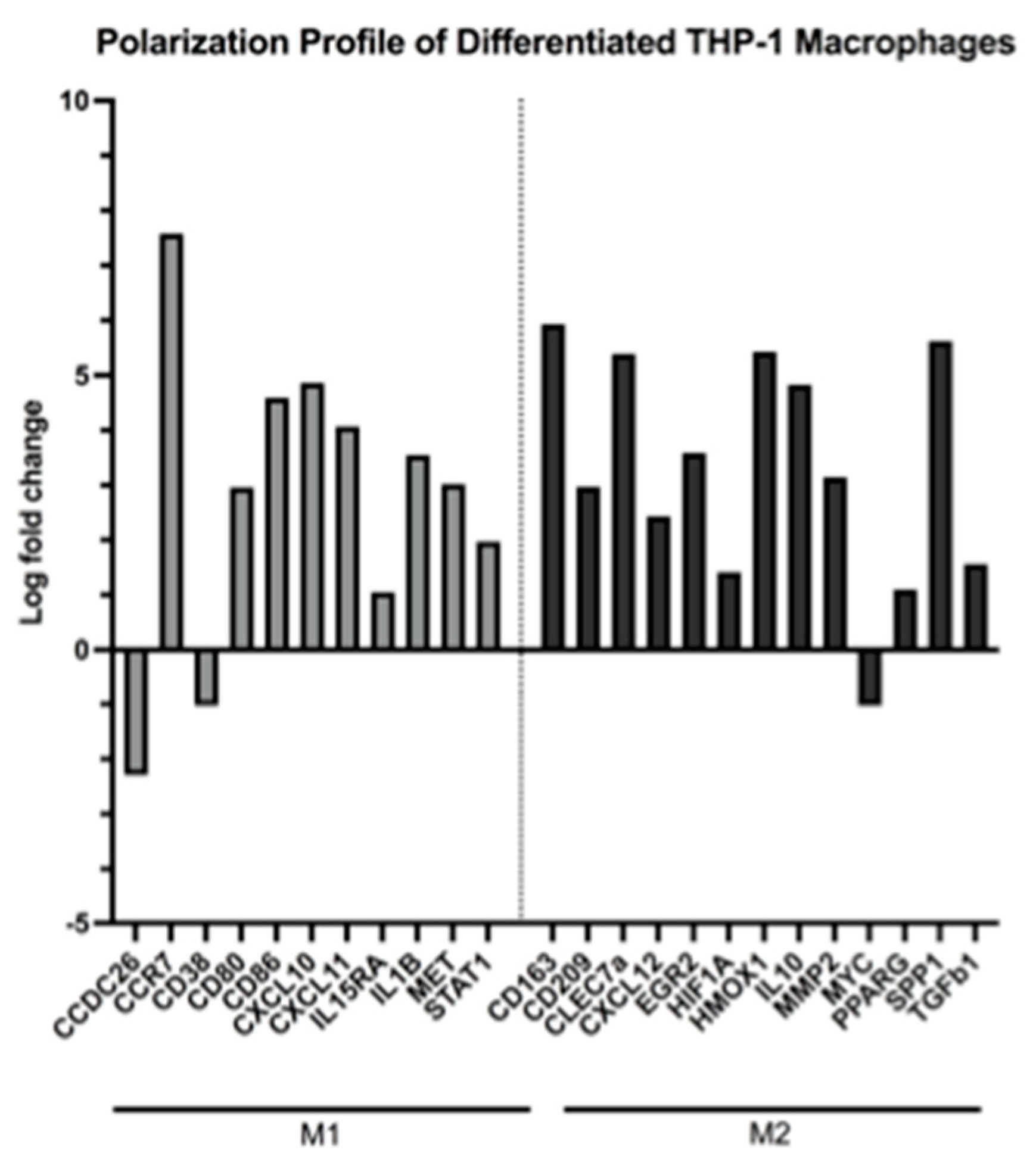
2.3. Key Genes Associated with Macrophage Differentiation and Mixed M1/M2-like Genotypes Are Up-Regulated in PMA-Treated THP-1 Cells
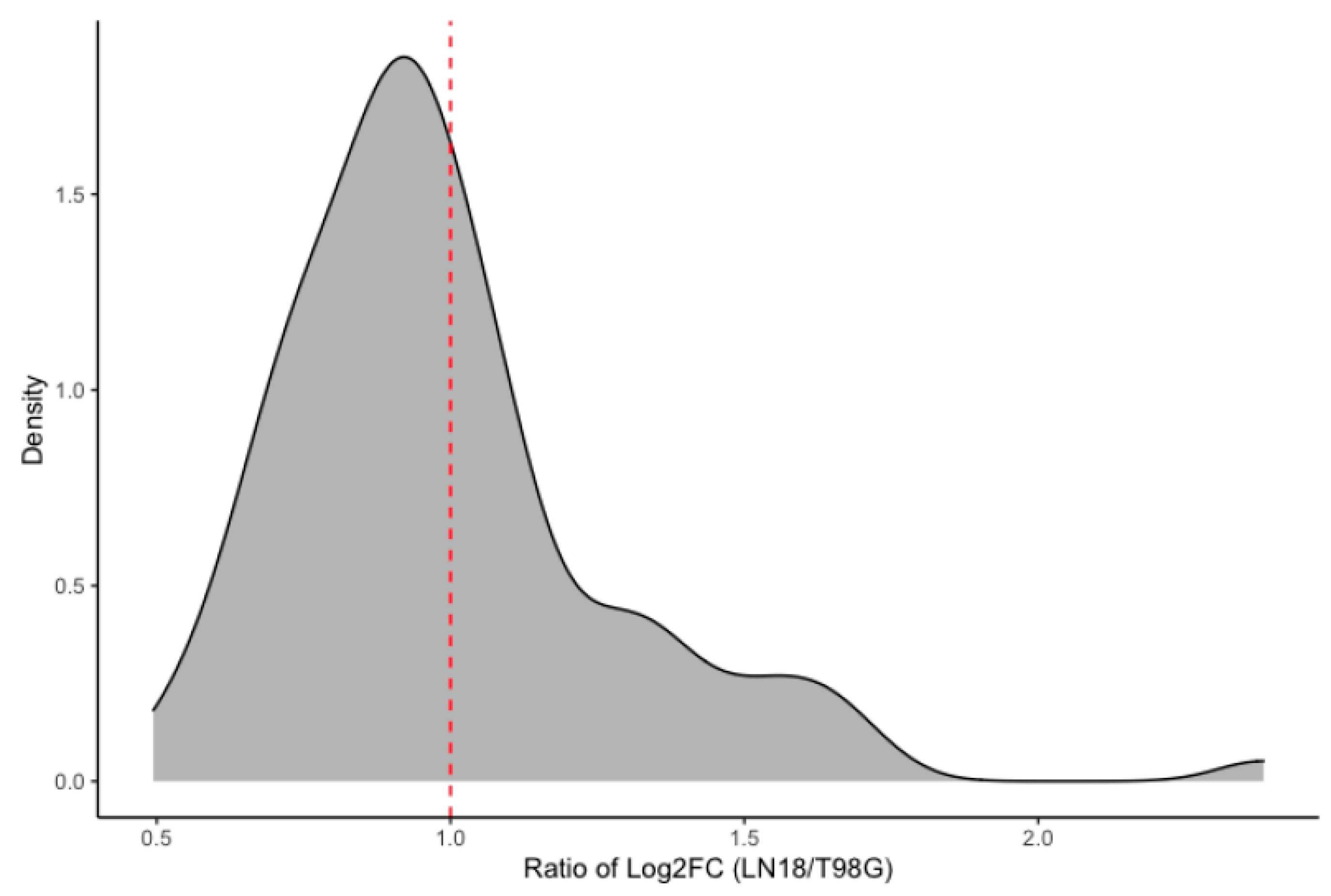
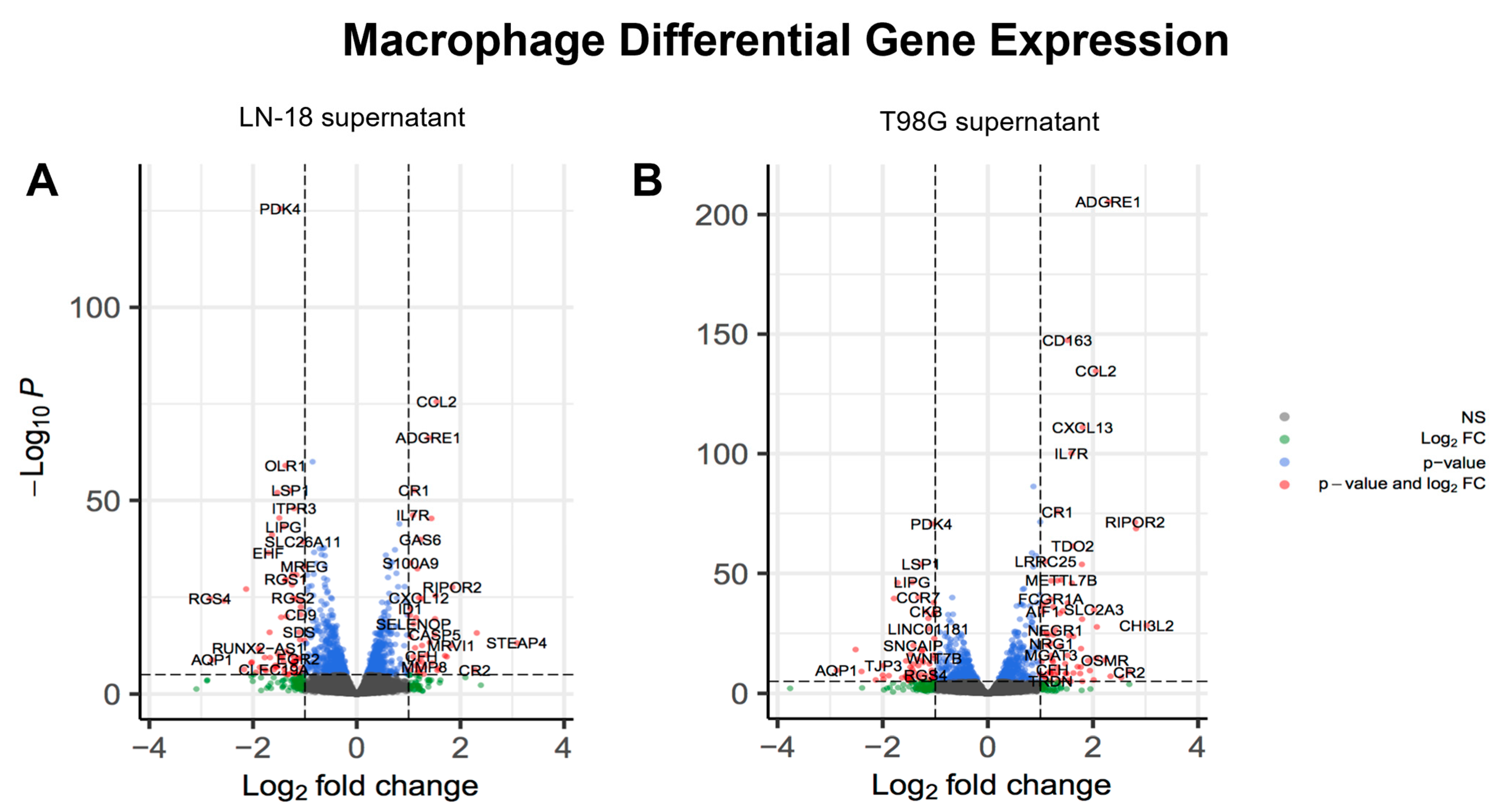
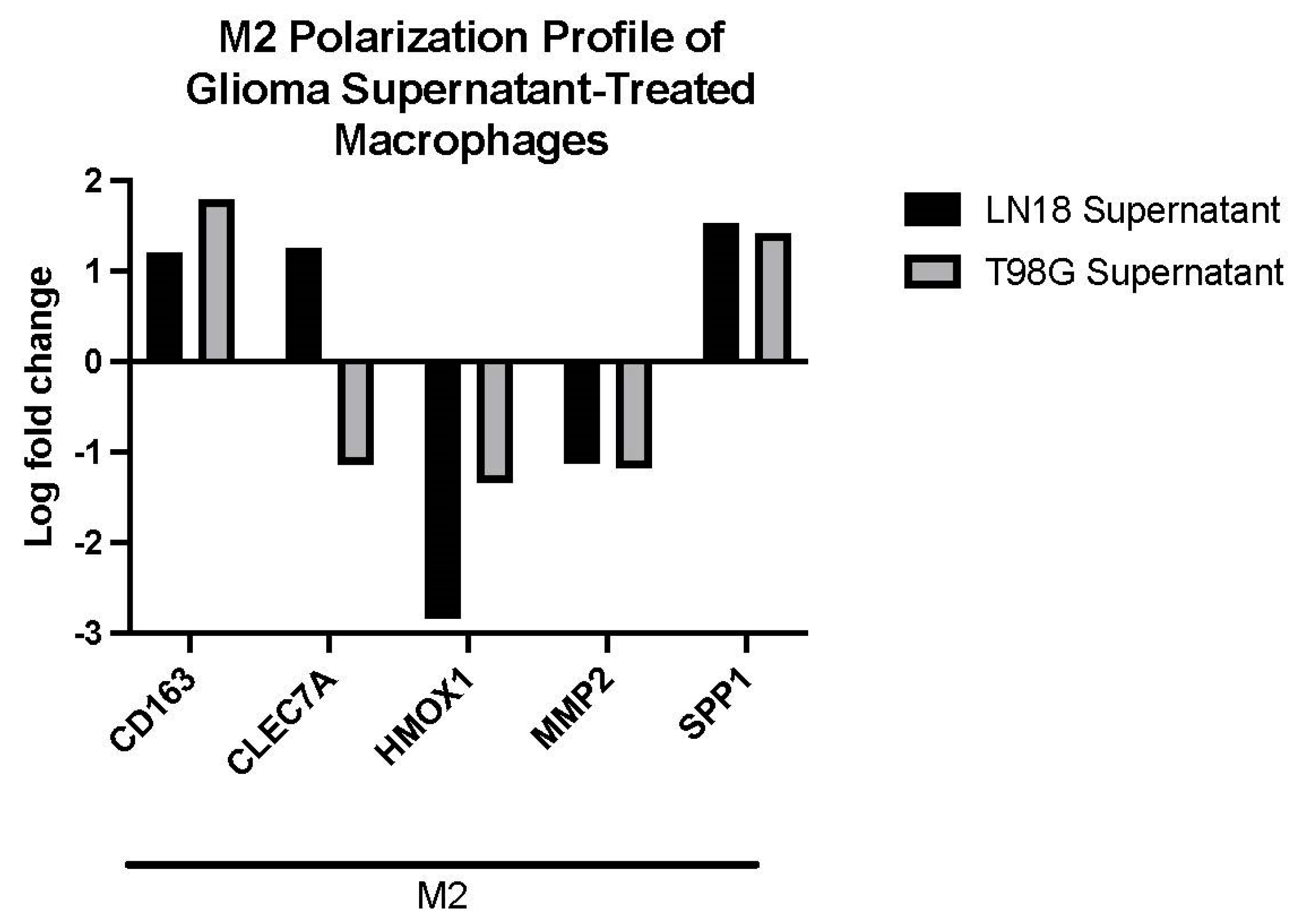
2.4. Glioma LN-18 or T98G Secretions Show Distinct Impact on Gene Expression Profile of THP-1 Macrophages
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Treatments
4.3. Sample Preparation and mRNA Sequencing
4.4. Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Low, J.T.; Ostrom, Q.T.; Cioffi, G.; Neff, C.; Waite, K.A.; Kruchko, C.; Barnholtz-Sloan, J.S. Primary brain and other central nervous system tumors in the United States (2014–2018): A summary of the CBTRUS statistical report for clinicians. Neuro-Oncol. Pract. 2022, 9, 165–182. [Google Scholar] [CrossRef]
- Holland, E.C. Glioblastoma multiforme: The terminator. Proc. Natl. Acad. Sci. USA 2000, 97, 6242–6244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, H.; Verhaak, R.G.W.; Canoll, P. The cellular origin for malignant glioma and prospects for clinical advancements. Expert Rev. Mol. Diagn. 2012, 12, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Zong, H.; Parada, L.F.; Baker, S.J. Cell of origin for malignant gliomas and its implication in therapeutic development. Cold Spring Harb. Perspect. Biol. 2015, 7, a020610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcantara Llaguno, S.R.; Parada, L.F. Cell of origin of glioma: Biological and clinical implications. Br. J. Cancer 2016, 115, 1445–1450. [Google Scholar] [CrossRef] [PubMed]
- Pesenti, C.; Navone, S.E.; Guarnaccia, L.; Terrasi, A.; Costanza, J.; Silipigni, R.; Guarneri, S.; Fusco, N.; Fontana, L.; Locatelli, M.; et al. The Genetic Landscape of Human Glioblastoma and Matched Primary Cancer Stem Cells Reveals Intratumour Similarity and Intertumour Heterogeneity. Stem Cells Int. 2019, 2019, 2617030. [Google Scholar] [CrossRef]
- Pine, A.R.; Cirigliano, S.M.; Nicholson, J.G.; Hu, Y.; Linkous, A.; Miyaguchi, K.; Edwards, L.; Singhania, R.; Schwartz, T.H.; Ramakrishna, R.; et al. Tumor Microenvironment Is Critical for the Maintenance of Cellular States Found in Primary Glioblastomas. Cancer Discov. 2020, 10, 964–979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, M.; Hassiotou, F.; Nowak, A. Glioblastoma stem-like cells: At the root of tumor recurrence and a therapeutic target. Carcinogenesis 2014, 36, 177–185. [Google Scholar] [CrossRef]
- Xie, Q.; Mittal, S.; Berens, M.E. Targeting adaptive glioblastoma: An overview of proliferation and invasion. Neuro-Oncology 2014, 16, 1575–1584. [Google Scholar] [CrossRef] [Green Version]
- Ostrom, Q.T.; Gittleman, H.; Truitt, G.; Boscia, A.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2011–2015. Neuro-Oncology 2018, 20, iv1–iv86. [Google Scholar] [CrossRef] [Green Version]
- Alifieris, C.; Trafalis, D.T. Glioblastoma multiforme: Pathogenesis and treatment. Pharmacol. Ther. 2015, 152, 63–82. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stupp, R.; Hegi, M.E.; Mason, W.P.; van den Bent, M.J.; Taphoorn, M.J.B.; Janzer, R.C.; Ludwin, S.K.; Allgeier, A.; Fisher, B.; Belanger, K.; et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009, 10, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Chinot, O.L.; Wick, W.; Mason, W.; Henriksson, R.; Saran, F.; Nishikawa, R.; Carpentier, A.F.; Hoang-Xuan, K.; Kavan, P.; Cernea, D.; et al. Bevacizumab plus Radiotherapy–Temozolomide for Newly Diagnosed Glioblastoma. N. Engl. J. Med. 2014, 370, 709–722. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, M.R.; Dignam, J.J.; Armstrong, T.S.; Wefel, J.S.; Blumenthal, D.T.; Vogelbaum, M.A.; Colman, H.; Chakravarti, A.; Pugh, S.; Won, M.; et al. A Randomized Trial of Bevacizumab for Newly Diagnosed Glioblastoma. N. Engl. J. Med. 2014, 370, 699–708. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ali, S.; Clarke, J.; Cha, S. Bevacizumab in Recurrent Glioma: Patterns of Treatment Failure and Implications. Brain Tumor Res. Treat. 2017, 5, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Schalper, K.A.; Rodriguez-Ruiz, M.E.; Diez-Valle, R.; López-Janeiro, A.; Porciuncula, A.; Idoate, M.A.; Inogés, S.; de Andrea, C.; López-Diaz de Cerio, A.; Tejada, S.; et al. Neoadjuvant nivolumab modifies the tumor immune microenvironment in resectable glioblastoma. Nat. Med. 2019, 25, 470–476. [Google Scholar] [CrossRef]
- Pearson, J.R.D.; Regad, T. Targeting cellular pathways in glioblastoma multiforme. Signal Transduct. Target. Ther. 2017, 2, 17040. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Lage, M.; Lynch, T.M.; Bi, Y.; Cocito, C.; Way, G.P.; Pal, S.; Haller, J.; Yan, R.E.; Ziober, A.; Nguyen, A.; et al. Immune landscapes associated with different glioblastoma molecular subtypes. Acta Neuropathol. Commun. 2019, 7, 203. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Song, Z.; Zhang, T.; He, X.; Huang, K.; Zhang, Q.; Shen, J.; Pan, J. Identification of immune cell infiltration and immune-related genes in the tumor microenvironment of glioblastomas. Front. Immunol. 2020, 11, 585034. [Google Scholar] [CrossRef]
- Gabrusiewicz, K.; Ellert-Miklaszewska, A.; Lipko, M.; Sielska, M.; Frankowska, M.; Kaminska, B. Characteristics of the alternative phenotype of microglia/macrophages and its modulation in experimental gliomas. PLoS ONE 2011, 6, e23902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrusiewicz, K.; Rodriguez, B.; Wei, J.; Hashimoto, Y.; Healy, L.M.; Maiti, S.N.; Thomas, G.; Zhou, S.; Wang, Q.; Elakkad, A.; et al. Glioblastoma-infiltrated innate immune cells resemble M0 macrophage phenotype. JCI Insight 2016, 1, e85841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage Polarization: Different Gene Signatures in M1(LPS+) vs. Classically and M2(LPS–) vs. Alternatively Activated Macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, C.C.; Li, M. Macrophages/Microglia in the Glioblastoma Tumor Microenvironment. Int. J. Mol. Sci. 2021, 22, 5775. [Google Scholar] [CrossRef] [PubMed]
- Diserens, A.C.; de Tribolet, N.; Martin-Achard, A.; Gaide, A.C.; Schnegg, J.F.; Carrel, S. Characterization of an established human malignant glioma cell line: LN-18. Acta Neuropathol. 1981, 53, 21–28. [Google Scholar] [CrossRef]
- Patil, V.; Pal, J.; Somasundaram, K. Elucidating the cancer-specific genetic alteration spectrum of glioblastoma derived cell lines from whole exome and RNA sequencing. Oncotarget 2015, 6, 43452–43471. [Google Scholar] [CrossRef] [Green Version]
- Gramatzki, D.; Pantazis, G.; Schittenhelm, J.; Tabatabai, G.; Kohle, C.; Wick, W.; Schwarz, M.; Weller, M.; Tritschler, I. Aryl hydrocarbon receptor inhibition downregulates the TGF-[beta]/Smad pathway in human glioblastoma cells. Oncogene 2009, 28, 2593–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verhaak, R.G.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar] [CrossRef] [Green Version]
- Genin, M.; Clement, F.; Fattaccioli, A.; Raes, M.; Michiels, C. M1 and M2 macrophages derived from THP-1 cells differentially modulate the response of cancer cells to etoposide. BMC Cancer 2015, 15, 577. [Google Scholar] [CrossRef] [Green Version]
- Starr, T.; Bauler, T.J.; Malik-Kale, P.; Steele-Mortimer, O. The phorbol 12-myristate-13-acetate differentiation protocol is critical to the interaction of THP-1 macrophages with Salmonella Typhimurium. PLoS ONE 2018, 13, e0193601. [Google Scholar] [CrossRef]
- Bujor, A.M.; El Adili, F.; Parvez, A.; Marden, G.; Trojanowska, M. Fli1 Downregulation in Scleroderma Myeloid Cells Has Profibrotic and Proinflammatory Effects. Front. Immunol. 2020, 11, 800. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Friedman, M.S.; Shedden, K.; Hankenson, K.D.; Woolf, P.J. GAGE: Generally applicable gene set enrichment for pathway analysis. BMC Bioinform. 2009, 10, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiecień, I.; Polubiec-Kownacka, M.; Dziedzic, D.; Wołosz, D.; Rzepecki, P.; Domagała-Kulawik, J. CD163 and CCR7 as markers for macrophage polarization in lung cancer microenvironment. Cent. Eur. J. Immunol. 2019, 44, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Bertani, F.R.; Mozetic, P.; Fioramonti, M.; Iuliani, M.; Ribelli, G.; Pantano, F.; Santini, D.; Tonini, G.; Trombetta, M.; Businaro, L.; et al. Classification of M1/M2-polarized human macrophages by label-free hyperspectral reflectance confocal microscopy and multivariate analysis. Sci. Rep. 2017, 7, 8965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, T.; Chen, J.; Xu, G.; Zhang, Z.; Xue, J.; Zeng, H.; Jiang, J.; Chen, T.; Qin, Z.; Li, H.; et al. STAT1 and CXCL10 involve in M1 macrophage polarization that may affect osteolysis and bone remodeling in extrapulmonary tuberculosis. Gene 2022, 809, 146040. [Google Scholar] [CrossRef] [PubMed]
- Dipta, P.; Sarsenbayeva, A.; Shmuel, M.; Forno, F.; Eriksson, J.W.; Pereira, M.J.; Abalo, X.M.; Wabitsch, M.; Thaysen-Andersen, M.; Tirosh, B. Macrophage-derived secretome is sufficient to confer olanzapine-mediated insulin resistance in human adipocytes. Compr. Psychoneuroendocrinol. 2021, 7, 100073. [Google Scholar] [CrossRef] [PubMed]
- Takiguchi, H.; Yang, C.X.; Yang, C.W.T.; Sahin, B.; Whalen, B.A.; Milne, S.; Akata, K.; Yamasaki, K.; Yang, J.S.W.; Cheung, C.Y.; et al. Macrophages with reduced expressions of classical M1 and M2 surface markers in human bronchoalveolar lavage fluid exhibit pro-inflammatory gene signatures. Sci. Rep. 2021, 11, 8282. [Google Scholar] [CrossRef]
- Xu, Z.A.-O.; Gu, Y.; Wang, C.Z.; Jin, Y.; Wen, X.M.; Ma, J.C.; Tang, L.J.; Mao, Z.W.; Qian, J.; Lin, J.A.-O. The M2 macrophage marker CD206: A novel prognostic indicator for acute myeloid leukemia. Oncoimmunology 2020, 9, 1683347. [Google Scholar] [CrossRef] [Green Version]
- Szulzewsky, F.; Pelz, A.; Feng, X.; Synowitz, M.; Markovic, D.; Langmann, T.; Holtman, I.R.; Wang, X.; Eggen, B.J.L.; Boddeke, H.W.G.M.; et al. Glioma-Associated Microglia/Macrophages Display an Expression Profile Different from M1 and M2 Polarization and Highly Express Gpnmb and Spp1. PLoS ONE 2015, 10, e0116644. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Hardie, J.; Landis, R.F.; Mas-Rosario, J.A.; Chattopadhyay, A.N.; Keshri, P.; Sun, J.; Rizzo, E.M.; Gopalakrishnan, S.; Farkas, M.A.-O.; et al. High-content and high-throughput identification of macrophage polarization phenotypes. Chem. Sci. 2020, 11, 8231–8239. [Google Scholar] [CrossRef]
- Naito, Y.; Takagi, T.; Higashimura, Y. Heme oxygenase-1 and anti-inflammatory M2 macrophages. Arch. Biochem. Biophys. 2014, 564, 83–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vats, D.; Mukundan, L.; Odegaard, J.I.; Zhang, L.; Smith, K.L.; Morel, C.R.; Greaves, D.R.; Murray, P.J.; Chawla, A. Oxidative metabolism and PGC-1β attenuate macrophage-mediated inflammation. Cell Metab. 2006, 4, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, P.D.; Ebert, D.; Muruganujan, A.; Mushayahama, T.; Albou, L.-P.; Mi, H. PANTHER: Making genome-scale phylogenetics accessible to all. Protein Sci. 2022, 31, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Burgler, S. Role of CD38 Expression in Diagnosis and Pathogenesis of Chronic Lymphocytic Leukemia and Its Potential as Therapeutic Target. Crit. Rev.™ Immunol. 2015, 35, 417–432. [Google Scholar] [CrossRef]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage activation and polarization: Nomenclature and experimental guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Scobie, M.R.; Houke, H.R.; Rice, C.D. Modulation of glioma-inflammation crosstalk profiles in human glioblastoma cells by indirubin-3′-(2,3 dihydroxypropyl)-oximether (E804) and 7-bromoindirubin-3′-oxime (7BIO). Chem.-Biol. Interact. 2019, 312, 108816. [Google Scholar] [CrossRef]
- Chiba, S.; Ikushima, H.; Ueki, H.; Yanai, H.; Kimura, Y.; Hangai, S.; Nishio, J.; Negishi, H.; Tamura, T.; Saijo, S.; et al. Recognition of tumor cells by Dectin-1 orchestrates innate immune cells for anti-tumor responses. eLife 2014, 3, e04177. [Google Scholar] [CrossRef]
- Shan, Y.; He, X.; Song, W.; Han, D.; Niu, J.; Wang, J. Role of IL-6 in the invasiveness and prognosis of glioma. Int. J. Clin. Exp. Med. 2015, 8, 9114–9120. [Google Scholar]
- Song, L.; Liu, L.; Wu, Z.; Li, Y.; Ying, Z.; Lin, C.; Wu, J.; Hu, B.; Cheng, S.-Y.; Li, M.; et al. TGF-β induces miR-182 to sustain NF-κB activation in glioma subsets. J. Clin. Investig. 2012, 122, 3563–3578. [Google Scholar] [CrossRef] [Green Version]
- Bhat, K.P.; Balasubramaniyan, V.; Vaillant, B.; Ezhilarasan, R.; Hummelink, K.; Hollingsworth, F.; Wani, K.; Heathcock, L.; James, J.D.; Goodman, L.D.; et al. Mesenchymal Differentiation Mediated by NF-κB Promotes Radiation Resistance in Glioblastoma. Cancer Cell 2013, 24, 331–346. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol ID | Log FC | Log CPM | LR | P | FDR |
|---|---|---|---|---|---|
| HLA-DRA | 3.01 | 2.79 | 258.12 | 4.40 × 10−58 | 4.60 × 10−57 |
| IL4R (CD124) | 1.23 | 5.72 | 386.22 | 5.52 × 10−86 | 8.90 × 10−85 |
| CSF1 (M-SCF) | 1.67 | 1.30 | 46.50 | 9.14 × 10−12 | 2.70 × 10−11 |
| CSF1R (CD115) | 1.87 | 8.19 | 1528.51 | 0 | 0 |
| ITGAM (CD11b) | 2.71 | 6.03 | 1762.36 | 0 | 0 |
| ATP6V1F (CD14) | 1.07 | 6.81 | 533.69 | 4.45 × 10−118 | 9.76 × 10−117 |
| Fold Enrichment | P | FDR | Ontology | |
|---|---|---|---|---|
| Up-regulated by LN18 supernatants | ||||
| immune complex clearance (GO:0002434) | >100.00 | 9.40 × 10−5 | 1.84 × 10−2 | BP |
| negative regulation of vascular endothelial cell proliferation (GO:1905563) | >100.00 | 1.97 × 10−4 | 3.24 × 10−2 | BP |
| antigen processing and presentation, endogenous lipid antigen via MHC class Ib (GO:0048006) | >100.00 | 1.97 × 10−4 | 3.21 × 10−2 | BP |
| antigen processing and presentation, exogenous lipid antigen via MHC class Ib (GO:0048007) | 91.92 | 3.36 × 10−4 | 4.35 × 10−2 | BP |
| regulation of astrocyte activation (GO:0061888) | >100.00 | 1.97 × 10−4 | 3.18 × 10−2 | BP |
| leukocyte chemotaxis (GO:0030595) | 19.70 | 1.23 × 10−9 | 6.40 × 10−6 | BP |
| leukocyte migration (GO:0050900) | 12.43 | 5.64 × 10−8 | 8.04 × 10−5 | BP |
| negative regulation of dendritic cell apoptotic process (GO:2000669) | 91.92 | 3.36 × 10−4 | 4.38 × 10−2 | BP |
| positive regulation of microglial cell activation (GO:1903980) | 80.43 | 4.19 × 10−4 | 4.97 × 10−2 | BP |
| positive regulation of neuroinflammatory response (GO:0150078) | 64.34 | 2.24 × 10−5 | 6.50 × 10−3 | BP |
| positive regulation of inflammatory response (GO:0050729) | 15.86 | 3.89 × 10−7 | 3.39 × 10−4 | BP |
| Up-regulated by T98G supernatants | ||||
| immune complex clearance (GO:0002434) | >100.00 | 1.54 × 10−4 | 2.01 × 10−2 | BP |
| antigen processing and presentation, endogenous lipid antigen via MHC class Ib (GO:0048006) | >100.00 | 3.22 × 10−4 | 3.63 × 10−2 | BP |
| negative regulation of glial cell apoptotic process (GO:0034351) | 83.70 | 1.29 × 10−5 | 3.06 × 10−3 | BP |
| negative regulation of interleukin-10 production (GO:0032693) | 39.65 | 8.77 × 10−5 | 1.26 × 10−2 | BP |
| Fold Enrichment | P | FDR | Ontology | |
|---|---|---|---|---|
| Down-regulated by LN-18 supernatants | ||||
| peptide antigen assembly with MHC class II protein complex (GO:0002503) | 50.96 | 2.44 × 10−6 | 2.55 × 10−3 | BP |
| MHC class II protein complex assembly (GO:0002399) | 50.96 | 2.44 × 10−6 | 2.39 × 10−3 | BP |
| MHC protein complex assembly (GO:0002396) | 40.77 | 5.27 × 10−6 | 3.59 × 10−3 | BP |
| antigen processing and presentation of peptide antigen via MHC class II (GO:0002495) | 25.48 | 2.79 × 10−5 | 1.07 × 10−3 | BP |
| antigen processing and presentation of exogenous peptide antigen via MHC class II (GO:0019886) | 27.18 | 2.22 × 10−5 | 9.93 × 10−3 | BP |
| antigen processing and presentation of exogenous peptide antigen (GO:0002478) | 14.83 | 1.98 × 10−4 | 4.09 × 10−2 | BP |
| Down-regulated by T98G supernatants | ||||
| peptide antigen assembly with MHC class II protein complex (GO:0002503) | 45.96 | 5.98 × 10−5 | 5.21 × 10−2 | BP |
| MHC class II protein complex assembly (GO:0002399) | 45.96 | 5.98 × 10−5 | 4.94 × 10−2 | BP |
| defense response to Gram-negative bacterium (GO:0050829) | 13.47 | 4.49 × 10−5 | 5.41 × 10−2 | BP |
| cellular biogenic amine metabolic process (GO:0006576) | 12.90 | 5.46 × 10−5 | 5.35 × 10−2 | BP |
| cellular amine metabolic process (GO:0044106) | 12.90 | 5.46 × 10−5 | 5.04 × 10−2 | BP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scobie, M.R.; Abood, A.; Rice, C.D. Differential Transcriptome Responses in Human THP-1 Macrophages Following Exposure to T98G and LN-18 Human Glioblastoma Secretions: A Simplified Bioinformatics Approach to Understanding Patient-Glioma-Specific Effects on Tumor-Associated Macrophages. Int. J. Mol. Sci. 2023, 24, 5115. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065115
Scobie MR, Abood A, Rice CD. Differential Transcriptome Responses in Human THP-1 Macrophages Following Exposure to T98G and LN-18 Human Glioblastoma Secretions: A Simplified Bioinformatics Approach to Understanding Patient-Glioma-Specific Effects on Tumor-Associated Macrophages. International Journal of Molecular Sciences. 2023; 24(6):5115. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065115
Chicago/Turabian StyleScobie, Micaela R., Abdullah Abood, and Charles D. Rice. 2023. "Differential Transcriptome Responses in Human THP-1 Macrophages Following Exposure to T98G and LN-18 Human Glioblastoma Secretions: A Simplified Bioinformatics Approach to Understanding Patient-Glioma-Specific Effects on Tumor-Associated Macrophages" International Journal of Molecular Sciences 24, no. 6: 5115. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065115