
Antitumor Effect of Iscador on Breast Cancer Cell Lines with Different Metastatic Potential
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Cyto- and Photo-Toxicity
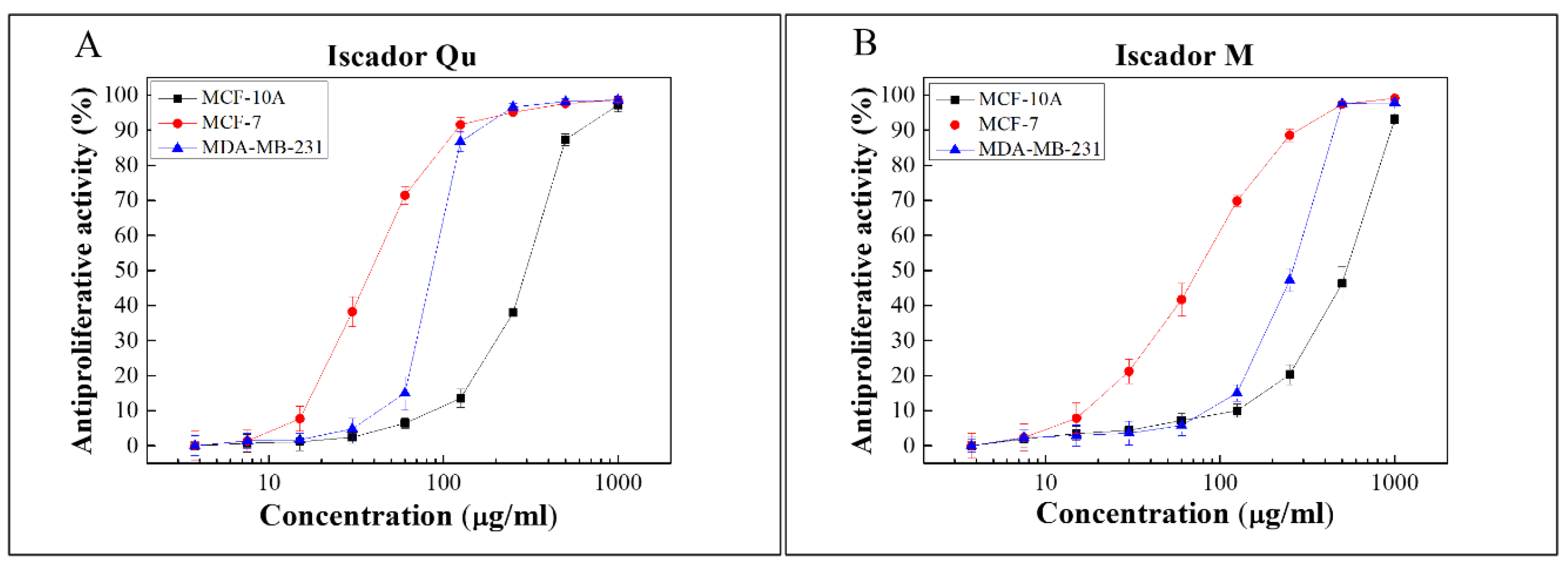
2.2. Antiproliferative Activity of Iscador Qu and Iscador M
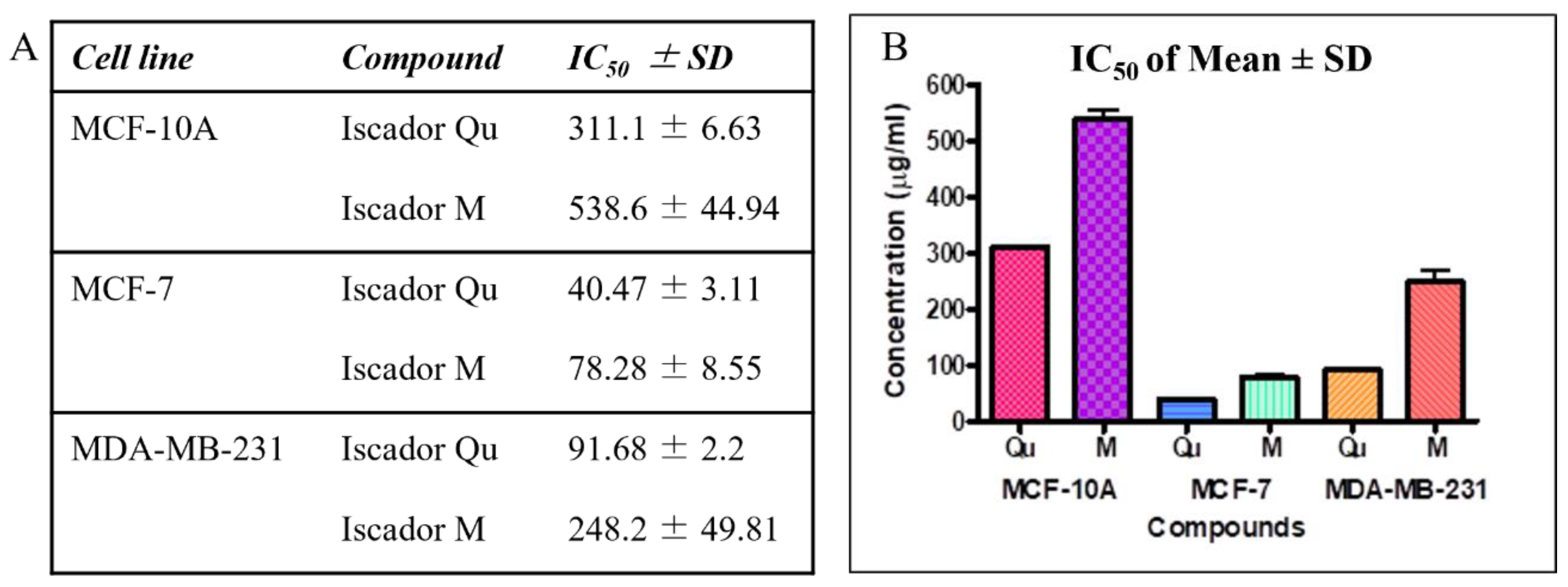
2.3. Selectivity of Iscador Qu and Iscador M to Malignant Cells
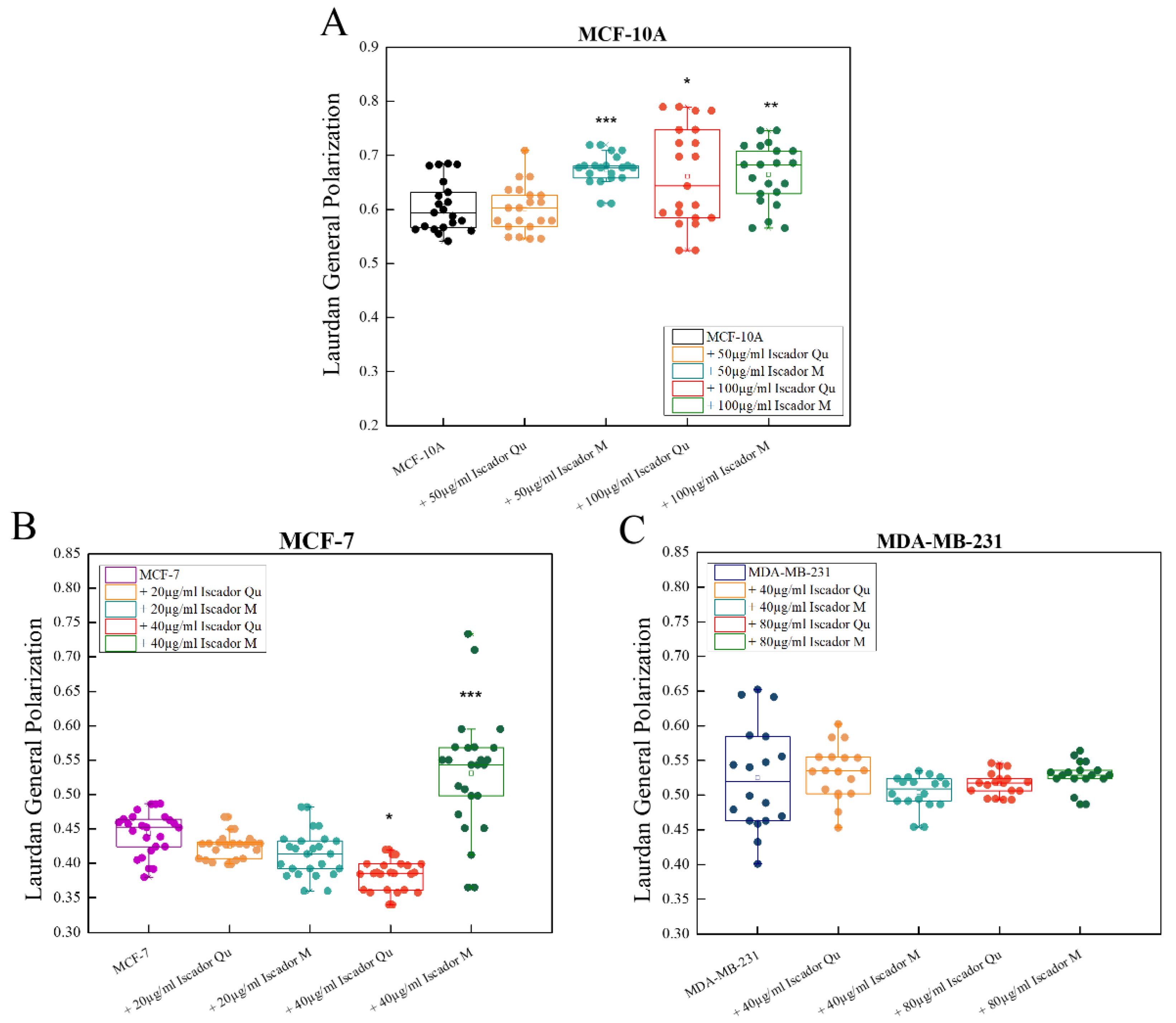
2.4. Changes in Membrane Lipid Order of the Studied Cell Lines under Iscador Treatment
2.5. Changes in ζ-Potential of the Studied Cell Lines under Iscador Treatment
2.6. Effect of Iscador Qu and Iscador M on Actin Cytoskeleton Organization and Migration
Actin Cytoskeleton
2.7. Cell Migration Assay
2.8. FACS Analysis
3. Materials and Methods
3.1. Drugs
3.2. Cell Lines
3.3. Cytotoxicity and Phototoxicity Testing
3.4. In Vitro Antiproliferative Activity
3.5. Assessment of the Plasma Membrane Lipid Order
3.6. ξ-Potential Measurements
3.7. Wound Healing (WH) Assay
3.8. Actin Labeling
3.9. FACS Analysis
3.10. Statistical Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kerr, A.J.; Dodwell, D.; Mcgale, P.; Holt, F.; Duane, F.; Mannu, G.; Darby, S.C.; Taylor, C.W. Adjuvant and neoadjuvant breast cancer treatments: A systematic review of their effects on mortality. Cancer Treat Rev. 2022, 105, 102375. [Google Scholar] [CrossRef]
- Elliott, M.J.; Wilson, B.; Cescon, D.W. Current Treatment and Future Trends of Immunotherapy in Breast Cancer. Curr. Cancer Drug Targets 2022, 22, 667–677. [Google Scholar]
- Lebert, J.; Lilly, E.J. Developments in the Management of Metastatic HER2-Positive Breast Cancer: A Review. Curr. Oncol. 2022, 29, 2539–2549. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, E.; Ulaner, G.A.; Chin, S.F. Molecular Classification of Breast Cancer. PET Clin. 2018, 13, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Asleh, K.; Riaz, N.; Nielsen, T.O. Heterogeneity of triple negative breast cancer: Current advances in subtyping and treatment implications. J. Exp. Clin. Cancer Res. 2022, 41, 265. [Google Scholar] [CrossRef]
- Huber-Keener, K.J. Cancer genetics and breast cancer. Best Pr. Res Clin. Obs. Gynaecol. 2022, 82, 3–11. [Google Scholar] [CrossRef]
- Vuong, D.; Simpson, P.T.; Green, B.; Cummings, M.C.; Lakhani, S.R. Molecular classification of breast cancer. Virchows Arch. 2014, 465, 1–14. [Google Scholar] [CrossRef]
- Kooti, W.; Servatyari, K.; Behzadifar, M.; Asadi-Samani, M.; Sadeghi, F.; Nouri, B.; Zare Marzouni, H. Effective Medicinal Plant in Cancer Treatment, Part 2: Review Study. J. Evid. Based Complement. Altern. Med. 2017, 22, 982–995. [Google Scholar] [CrossRef] [Green Version]
- Pfüller, U. Chemical Constituents of European Mistletoe (Viscum album L.) Isolation and Characterisation of the Main Relevant Ingredients: Lectins, Viscotoxins, Oligo-/polysaccharides, Flavonoides, Alkaloids. In Mistletoe; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Fu, L.L.; Zhou, C.C.; Yao, S.; Yu, J.Y.; Liu, B.; Bao, J.K. Plant lectins: Targeting programmed cell death pathways as antitumor agents. Int. J. Biochem. Cell Biol. 2011, 43, 1442–1449. [Google Scholar] [CrossRef]
- Beztsinna, N.; de Matos, M.B.C.; Walther, J.; Heyder, C.; Hildebrandt, E.; Leneweit, G.; Mastrobattista, E.; Kok, R.J. Quantitative analysis of receptor-mediated uptake and pro-apoptotic activity of mistletoe lectin-1 by high content imaging. Sci. Rep. 2018, 8, 2768. [Google Scholar] [CrossRef] [Green Version]
- de Virgilio, M.; Lombardi, A.; Caliandro, R.; Fabbrini, M.S. Ribosome-inactivating proteins: From plant defense to tumor attack. Toxins 2010, 2, 2699–2737. [Google Scholar] [CrossRef] [Green Version]
- Khwaja, T.A.; Dias, C.B.; Pentecost, S. Recent studies on the anticancer activities of mistletoe (Viscum album) and its alkaloids. Oncology 1986, 43 (Suppl. S1), 42–50. [Google Scholar] [CrossRef] [PubMed]
- Gabius, H.J.; Gabius, S.; Joshi, S.S.; Koch, B.; Schroeder, M.; Manzke, W.M.; Westerhausen, A. From Ill-Defined Extracts to the Immunomodulatory Lectin—Will There Be a Reason for Oncological Application of Mistletoe. Planta Med. 1994, 60, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Bocci, V. Mistletoe (Viscum-Album) Lectins as Cytokine Inducers and Immunoadjuvant in Tumor-Therapy—A Review. J. Biol. Regul. Homeost. Agents 1993, 7, 1–6. [Google Scholar] [PubMed]
- Schink, M. Mistletoe therapy for human cancer: The role of the natural killer cells. Anti-Cancer Drugs 1997, 8, S47–S51. [Google Scholar] [CrossRef]
- Zarkovic, N.; Trbojevic-Cepe, M.; Ilic, Z.; Hrzenjak, M.; Grainca, S.; Jurin, M. Comparison of the effects of high and low concentrations of the separated Viscum album L. lectins and of the plain mistletoe plant preparation (Isorel) on the growth of normal and tumor cells in vitro. Period. Biol. 1995, 97, 61–67. [Google Scholar]
- He, M.W.; Guo, S.; Li, Z.L. In situ characterizing membrane lipid phenotype of breast cancer cells using mass spectrometry profiling. Sci. Rep. 2015, 5, 11298. [Google Scholar] [CrossRef] [Green Version]
- Yousefvand, S.; Fattahi, F.; Hosseini, S.M.; Urech, K.; Schaller, G. Viscotoxin and lectin content in foliage and fruit of Viscum album L. on the main host trees of Hyrcanian forests. Sci. Rep. 2022, 12, 10383. [Google Scholar] [CrossRef]
- Szurpnicka, A.; Zjawiony, J.K.; Szterk, A. Therapeutic potential of mistletoe in CNS-related neurological disorders and the chemical composition of Viscum species. J. Ethnopharmacol. 2019, 231, 241–252. [Google Scholar] [CrossRef]
- Nikolai, G.; Friedl, P.; Werner, M.; Niggemann, B.; Zanker, K.S. Effect of a mistletoe extract (Iscador(R) QuFrF) on viability and migratory behavior of human peripheral CD4+ and CD8+ T lymphocytes in three-dimensional collagen lattices. In Vitro Cell. Dev. Biol.-Anim. 1997, 33, 710–716. [Google Scholar] [CrossRef]
- Stein, G.M.; Bussing, A.; Schietzel, M. Stimulation of the maturation of dendritic cells in vitro by a fermented mistletoe extract. Anticancer Res. 2002, 22, 4215–4219. [Google Scholar]
- Stirpe, F.; Sandvig, K.; Olsnes, S.; Pihl, A. Action of Viscumin, a Toxic Lectin from Mistletoe, on Cells in Culture. J. Biol. Chem. 1982, 257, 13271–13277. [Google Scholar] [CrossRef] [PubMed]
- Lyu, S.Y.; Park, W.B.; Choi, K.H.; Kim, W.H. Involvement of caspase-3 in apoptosis induced by Viscum album var. coloratum agglutinin in HL-60 cells. Biosci. Biotechnol. Biochem. 2001, 65, 534–541. [Google Scholar] [CrossRef] [Green Version]
- Lyu, S.Y.; Park, W.B. Mistletoe lectin (Viscum album coloratum) modulates proliferation and cytokine expressions in murine splenocytes. J. Biochem. Mol. Biol. 2006, 39, 662–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podlech, O.; Harter, P.N.; Mittelbronn, M.; Poschel, S.; Naumann, U. Fermented Mistletoe Extract as a Multimodal Antitumoral Agent in Gliomas. Evid.-Based Complement. Altern. Med. 2012, 2012, 501796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szurpnicka, A.; Kowalczuk, A.; Szterk, A. Biological activity of mistletoe: In vitro and in vivo studies and mechanisms of action. Arch. Pharm. Res. 2020, 43, 593–629. [Google Scholar] [CrossRef]
- Marvibaigi, M.; Supriyanto, E.; Amini, N.; Abdul Majid, F.A.; Jaganathan, S.K. Preclinical and clinical effects of mistletoe against breast cancer. Biomed Res. Int. 2014, 2014, 785479. [Google Scholar] [CrossRef] [Green Version]
- Thies, A.; Nugel, D.; Pfüller, U.; Moll, I.; Schumacher, U. Influence of mistletoe lectins and cytokines induced by them on cell proliferation of human melanoma cells in vitro. Toxicology 2005, 207, 105–116. [Google Scholar] [CrossRef]
- Schaller, G.; Urech, K.; Giannattasio, M. Cytotoxicity of different viscotoxins and extracts from the European subspecies of Viscum album L. Phytother. Res. 1996, 10, 473–477. [Google Scholar] [CrossRef]
- Felenda, J.E.; Turek, C.; Stintzing, F.C. Antiproliferative potential from aqueous Viscum album L. preparations and their main constituents in comparison with ricin and purothionin on human cancer cells. J. Ethnopharmacol. 2019, 236, 100–107. [Google Scholar] [CrossRef]
- Pelzer, F.; Troger, W.; Nat, D.R. Complementary Treatment with Mistletoe Extracts During Chemotherapy: Safety, Neutropenia, Fever, and Quality of Life Assessed in a Randomized Study. J. Altern. Complement. Med. 2018, 24, 954–961. [Google Scholar] [CrossRef]
- Siddiqui, A.J.; Jahan, S.; Singh, R.; Saxena, J.; Ashraf, S.A.; Khan, A.; Choudhary, R.K.; Balakrishnan, S.; Badraoui, R.; Bardakci, F.; et al. Plants in Anticancer Drug Discovery: From Molecular Mechanism to Chemoprevention. Biomed Res. Int. 2022, 2022, 5425485. [Google Scholar] [CrossRef]
- Tzoneva, R.; Stoyanova, T.; Petrich, A.; Popova, D.; Uzunova, V.; Momchilova, A.; Chiantia, S. Effect of Erufosine on Membrane Lipid Order in Breast Cancer Cell Models. Biomolecules 2020, 10, 802. [Google Scholar] [CrossRef]
- Balls, M. Statement on the scientific validity of the 3T3 NRU PT test (an in vitro test for phototoxic potential). ATLA 1998, 26, 7–8. [Google Scholar]
- Halle, W.; Halder, M.; Worth, A.; Genschow, E. The Registry of Cytotoxicity: Toxicity Testing in Cell Cultures to Predict Acute Toxicity (LD50) and to Reduce Testing in Animals. Altern. Lab. Anim. 2003, 31, 89. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Delgado, A.V.; Gonzalez-Caballero, F.; Hunter, R.J.; Koopal, L.K.; Lyklema, J. Measurement and interpretation of electrokinetic phenomena. J. Colloid. Interface Sci. 2007, 309, 194–224. [Google Scholar] [CrossRef] [PubMed]
- Khwaja, T.A.; Manjikian, S.P. Characterization of biologically active components of mistletoe. Cancer Res. 1990, 31, 412–416. [Google Scholar]
- Konopa, J.; Woynarowski, J.M.; Lewandowska-Gumieniak, M. Isolation of viscotoxins. Cytotoxic basic polypeptides from Viscum album L. Hoppe Seylers Z. Physiol. Chem. 1980, 361, 1525–1533. [Google Scholar] [CrossRef]
- Lee, R.T.; Gabius, H.J.; Lee, Y.C. Ligand binding characteristics of the major mistletoe lectin. J. Biol. Chem. 1992, 267, 23722–23727. [Google Scholar] [CrossRef]
- Hajto, T.; Hostanska, K.; Gabius, H.J. Modulatory potency of the beta-galactoside-specific lectin from mistletoe extract (Iscador) on the host defense system in vivo in rabbits and patients. Cancer Res. 1989, 49, 4803–4808. [Google Scholar] [PubMed]
- Urech, K.; Schaller, G.; Jaggy, C. Viscotoxins, mistletoe lectins and their isoforms in mistletoe (Viscum album L.) extracts Iscador—Analytical results on pharmaceutical processing of mistletoe. Arzneim.-Forsch.-Drug Res. 2006, 56, 428–434. [Google Scholar]
- Urech, K.; Schaller, G.; Ziska, P.; Giannattasio, M. Comparative study on the cytotoxic effect of viscotoxin and mistletoe lectin on tumour cells in culture. Phytother. Res. 1995, 9, 49–55. [Google Scholar] [CrossRef]
- Lladó, V.; López, D.J.; Ibarguren, M.; Alonso, M.; Soriano, J.B.; Escribá, P.V.; Busquets, X. Regulation of the cancer cell membrane lipid composition by NaCHOleate: Effects on cell signaling and therapeutical relevance in glioma. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2014, 1838, 1619–1627. [Google Scholar] [CrossRef] [Green Version]
- Bernardes, N.; Fialho, A.M. Perturbing the Dynamics and Organization of Cell Membrane Components: A New Paradigm for Cancer-Targeted Therapies. Int. J. Mol. Sci. 2018, 19, 3871. [Google Scholar] [CrossRef] [Green Version]
- Pakiet, A.; Kobiela, J.; Stepnowski, P.; Sledzinski, T.; Mika, A. Changes in lipids composition and metabolism in colorectal cancer: A review. Lipids Health Dis. 2019, 18, 29. [Google Scholar] [CrossRef] [Green Version]
- Zalba, S.; ten Hagen, T.L.M. Cell membrane modulation as adjuvant in cancer therapy. Cancer Treat. Rev 2017, 52, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Steinkühler, J.; Sezgin, E.; Urbančič, I.; Eggeling, C.; Dimova, R. Mechanical properties of plasma membrane vesicles correlate with lipid order, viscosity and cell density. Commun. Biol. 2019, 2, 337. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.G.; Kondo, T. Cancer cell heterogeneity and plasticity: From molecular understanding to therapeutic targeting. Semin. Cancer Biol. 2022, 82, 1–2. [Google Scholar] [CrossRef]
- Szlasa, W.; Zendran, I.; Zalesinska, A.; Tarek, M.; Kulbacka, J. Lipid composition of the cancer cell membrane. J. Bioenerg. Biomembr. 2020, 52, 321–342. [Google Scholar] [CrossRef]
- Todor, I.N.; Lukyanova, N.Y.; Chekhun, V.F. The lipid content of cisplatin- and doxorubicin-resistant MCF-7 human breast cancer cells. Exp. Oncol. 2012, 34, 97–100. [Google Scholar]
- Barceló-Coblijn, G.; Martin, M.L.; de Almeida, R.F.M.; Noguera-Salvà, M.A.; Marcilla-Etxenike, A.; Guardiola-Serrano, F.; Lüth, A.; Kleuser, B.; Halver, J.E.; Escribá, P.V. Sphingomyelin and sphingomyelin synthase (SMS) in the malignant transformation of glioma cells and in 2-hydroxyoleic acid therapy. Proc. Natl. Acad. Sci. USA 2011, 108, 19569–19574. [Google Scholar] [CrossRef] [Green Version]
- Händel, C.; Schmidt, B.U.S.; Schiller, J.; Dietrich, U.; Möhn, T.; Kießling, T.; Pawlizak, S.; Fritsch, A.W.; Horn, L.-C.; Briest, S.; et al. Cell membrane softening in human breast and cervical cancer cells. New J. Phys. 2015, 17, 083008. [Google Scholar] [CrossRef]
- Bondar, O.V.; Saifullina, D.V.; Shakhmaeva, I.I.; Mavlyutova, I.I.; Abdullin, T.I. Monitoring of the Zeta Potential of Human Cells upon Reduction in Their Viability and Interaction with Polymers. Acta Nat. 2012, 4, 78–81. [Google Scholar] [CrossRef]
- Srdic-Rajic, T.; Santibanez, J.F.; Kanjer, K.; Tisma-Miletic, N.; Cavic, M.; Galun, D.; Jevric, M.; Kardum, N.; Konic-Ristic, A.; Zoranovic, T. Iscador Qu inhibits doxorubicin-induced senescence of MCF7 cells. Sci. Rep. 2017, 7, 3763. [Google Scholar] [CrossRef] [Green Version]
- Raphael, T.J.; Kuttan, G. Effect of naturally occurring triterpenoids ursolic acid and glycyrrhizic acid on the cell-mediated immune responses of metastatic tumor-bearing animals. Immunopharmacol. Immunotoxicol. 2008, 30, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Maldacker, J. Preclinical Investigations with Mistletoe (Viscum album L.) Extract Iscador. Arzneimittelforschung 2006, 56, 497–507. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Selectivity Index (SI) | |||
|---|---|---|---|
| MCF-7 | MDA-MB-231 | ||
| Iscador Qu | Iscador M | Iscador Qu | Iscador M |
| 7.69 | 6.88 | 3.39 | 2.17 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robev, B.; Iliev, I.; Tsoneva, I.; Momchilova, A.; Nesheva, A.; Kostadinova, A.; Staneva, G.; Nikolova, B. Antitumor Effect of Iscador on Breast Cancer Cell Lines with Different Metastatic Potential. Int. J. Mol. Sci. 2023, 24, 5247. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065247
Robev B, Iliev I, Tsoneva I, Momchilova A, Nesheva A, Kostadinova A, Staneva G, Nikolova B. Antitumor Effect of Iscador on Breast Cancer Cell Lines with Different Metastatic Potential. International Journal of Molecular Sciences. 2023; 24(6):5247. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065247
Chicago/Turabian StyleRobev, Bozhil, Ivan Iliev, Iana Tsoneva, Albena Momchilova, Alexandrina Nesheva, Aneliya Kostadinova, Galya Staneva, and Biliana Nikolova. 2023. "Antitumor Effect of Iscador on Breast Cancer Cell Lines with Different Metastatic Potential" International Journal of Molecular Sciences 24, no. 6: 5247. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065247