

LINC01393, a Novel Long Non-Coding RNA, Promotes the Cell Proliferation, Migration and Invasion through MiR-128-3p/NUSAP1 Axis in Glioblastoma
 ,
,
Abstract
:1. Introduction
2. Results
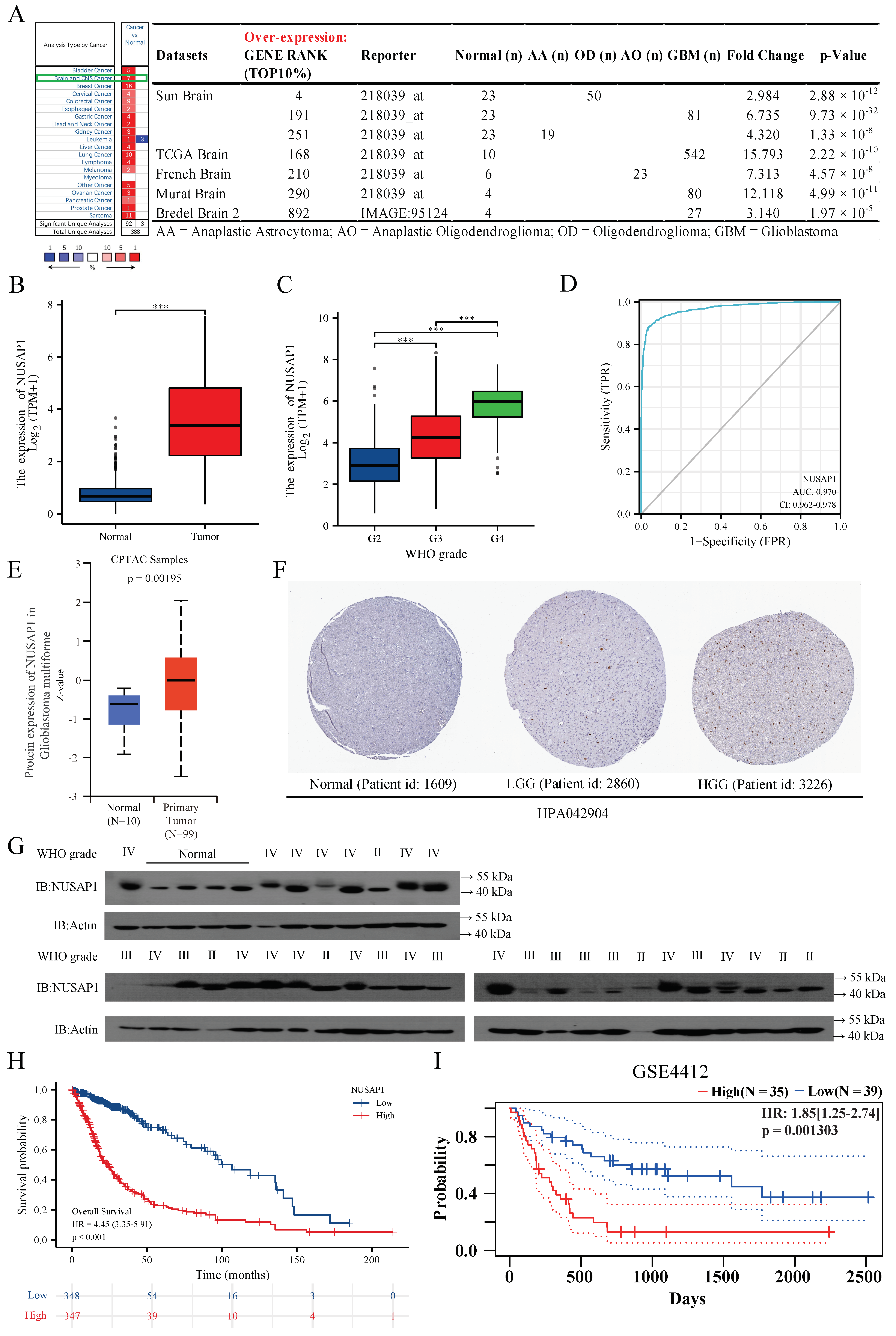
2.1. NUSAP1 Expression Is Elevated in Glioma with Higher Malignancy
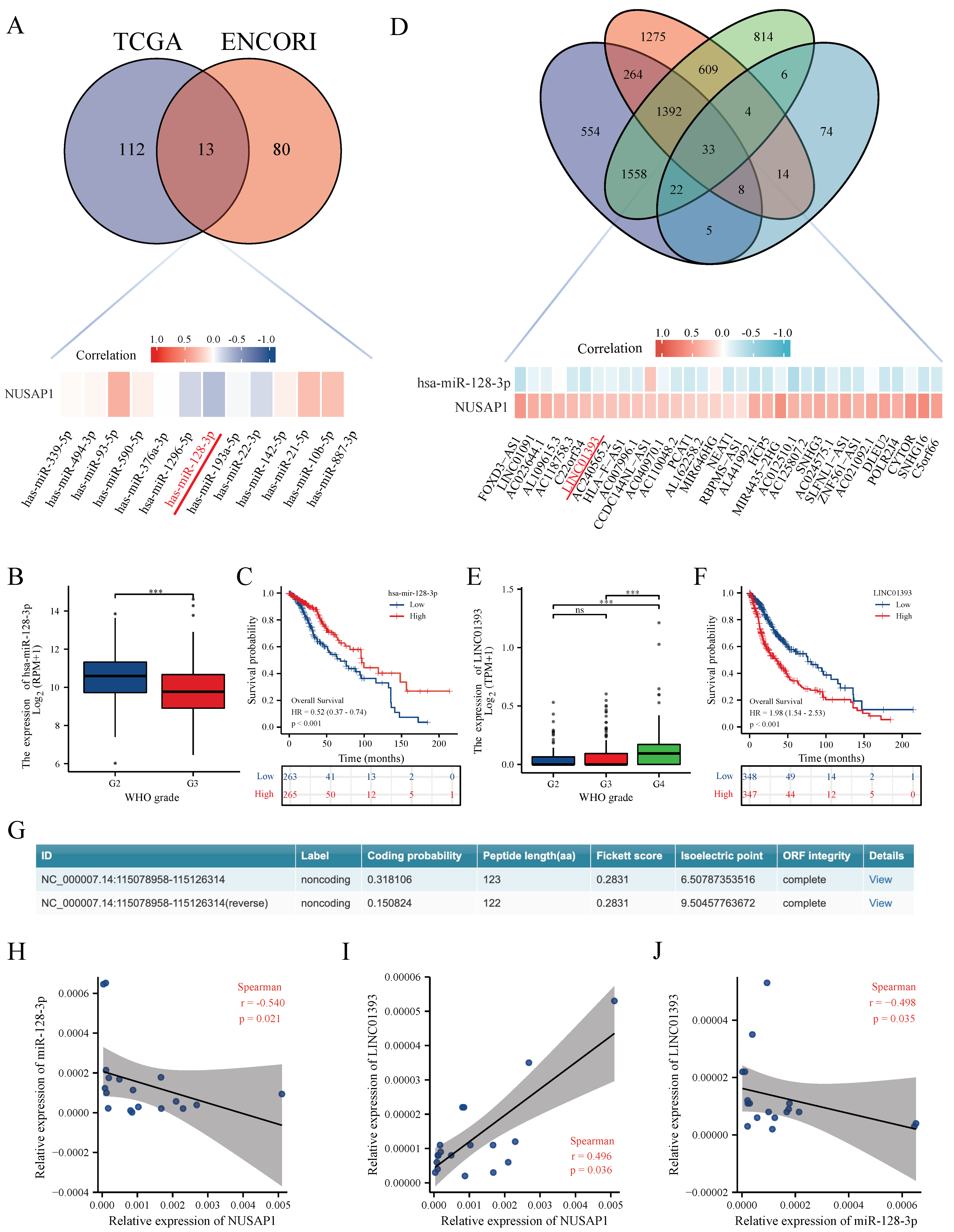
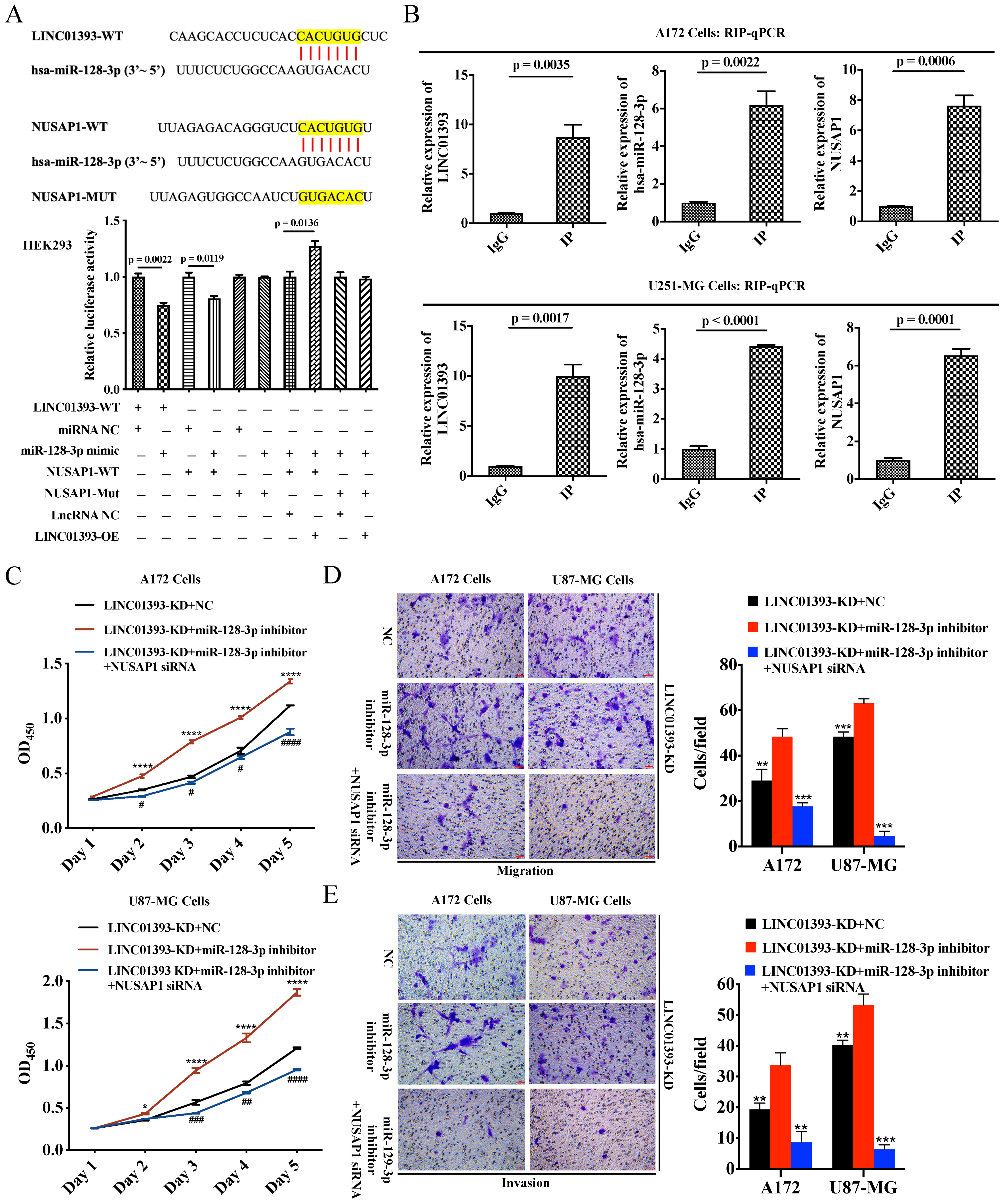
2.2. LINC01393 May Regulate the Expression of NUSAP1 by Acting as a ceRNA of miR-128-3p in Glioma
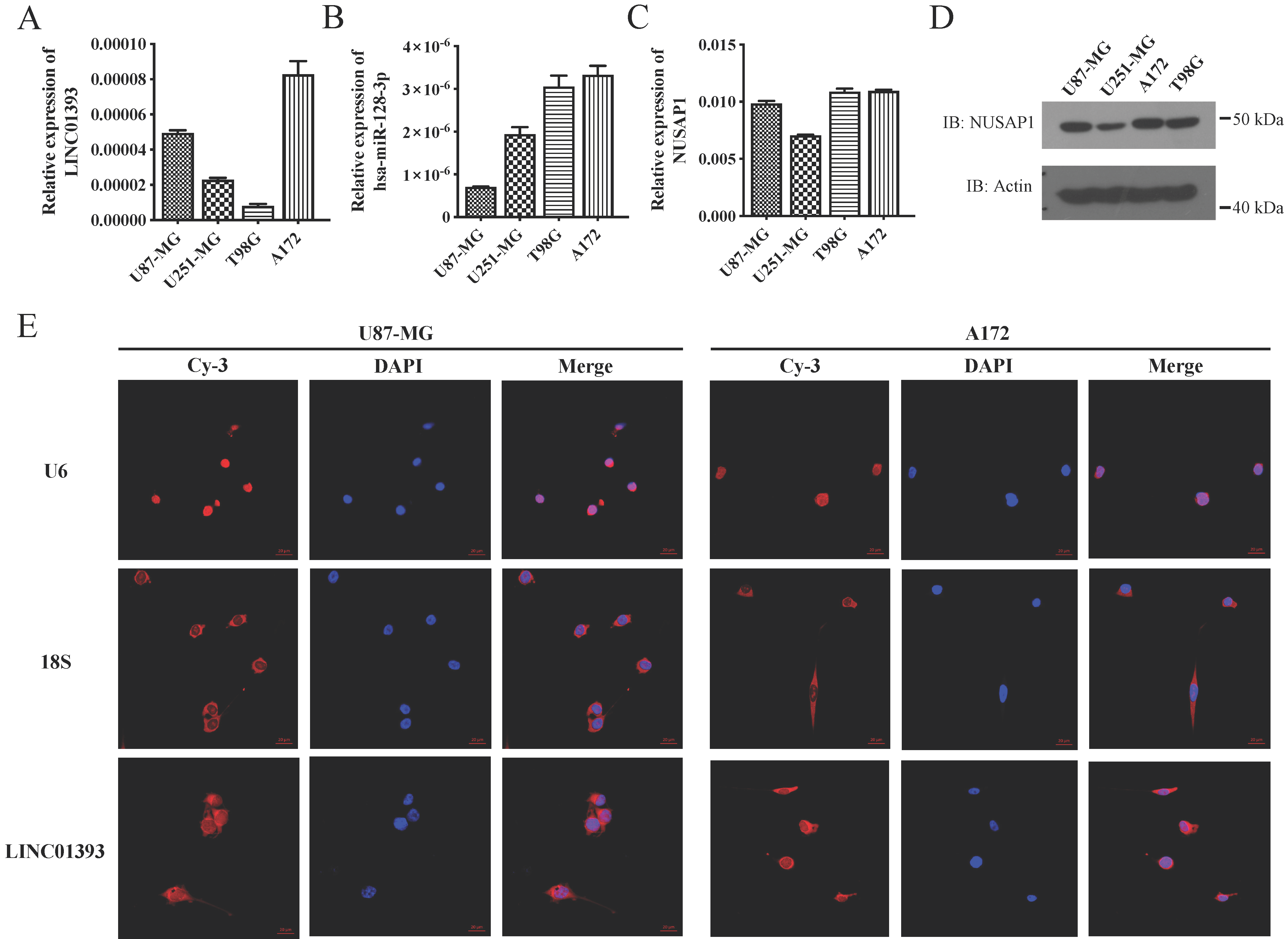
2.3. LINC01393, miR-128-3p, and NUSAP1 Expression in Glioma Cells
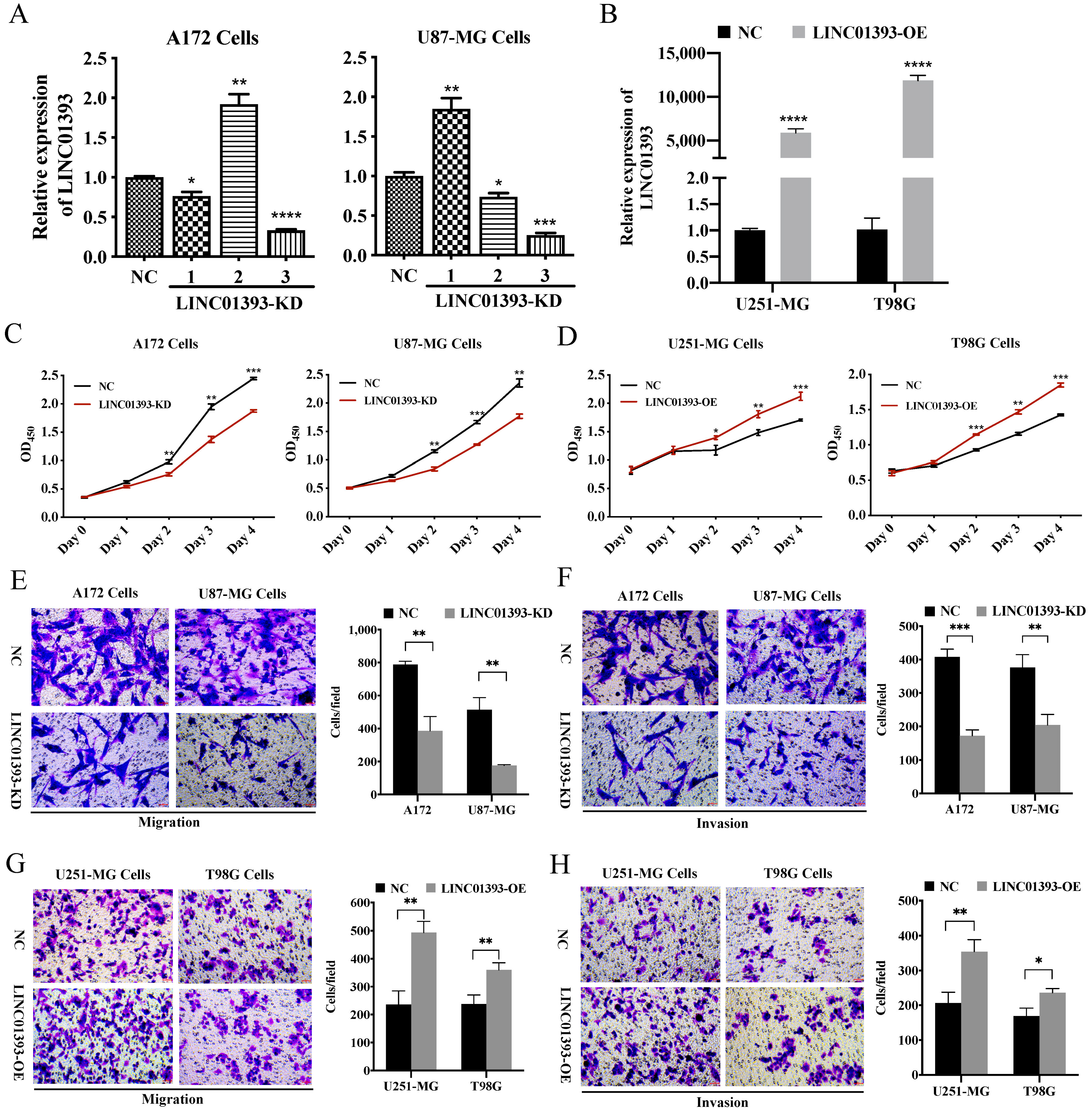
2.4. LINC01393 Promoted Proliferation, Migration and Invasion of GBM Cells
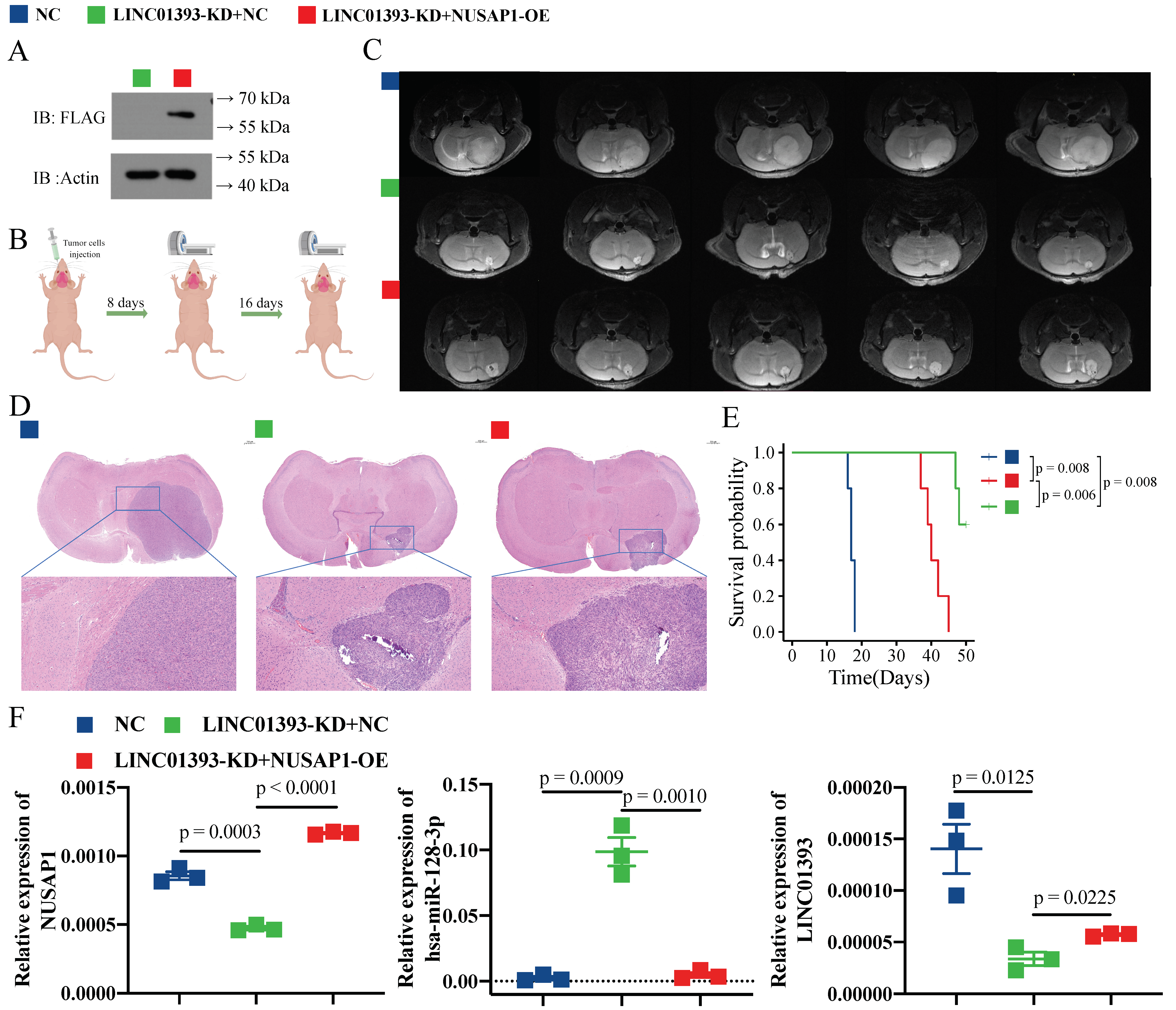
2.5. LINC01393 Functions as a ceRNA against miR-128-3p to Promote NUSAP1 Expression In Vivo
2.6. Knockdown of LINC01393 Inhibits GBM Progression In Vivo, and could Be Partly Reversed by NUSAP1 Overexpression
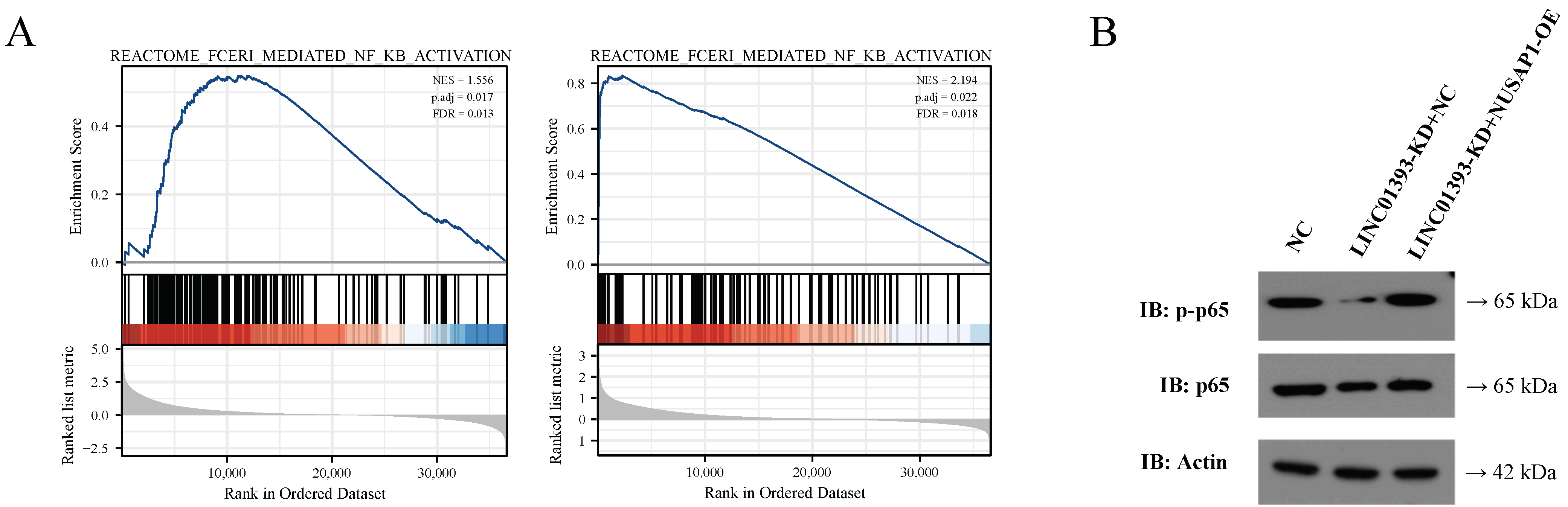
2.7. LINC01393/miR-128-3p/NUSAP1 Axis induce NF-κB Pathway Activation
3. Methods
3.1. NUSAP1 Expression of Multiple Databases Analysis
3.2. Candidate Upstream miRNAs and lncRNAs Prediction of NUSAP1
3.3. Human Tissues Collection
3.4. Cell Culture
3.5. Fluorescence In Situ Hybridization (FISH)
3.6. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Assays
3.7. Cell Transfection
3.8. LINC01393 Silencing and Overexpression
3.9. Western Blot
3.10. Cell Counting Kit (CCK)-8 Assay
3.11. Transwell Migration and Matrigel Invasion Assays
3.12. Luciferase Reporter Assays
3.13. RNA Immunoprecipitation Assay
3.14. Gene Set Enrichment Analysis (GSEA)
3.15. Animals
3.16. Orthotopic Intracranial GBM Model
3.17. H&E Staining
3.18. Statistical Analysis
4. Discussion
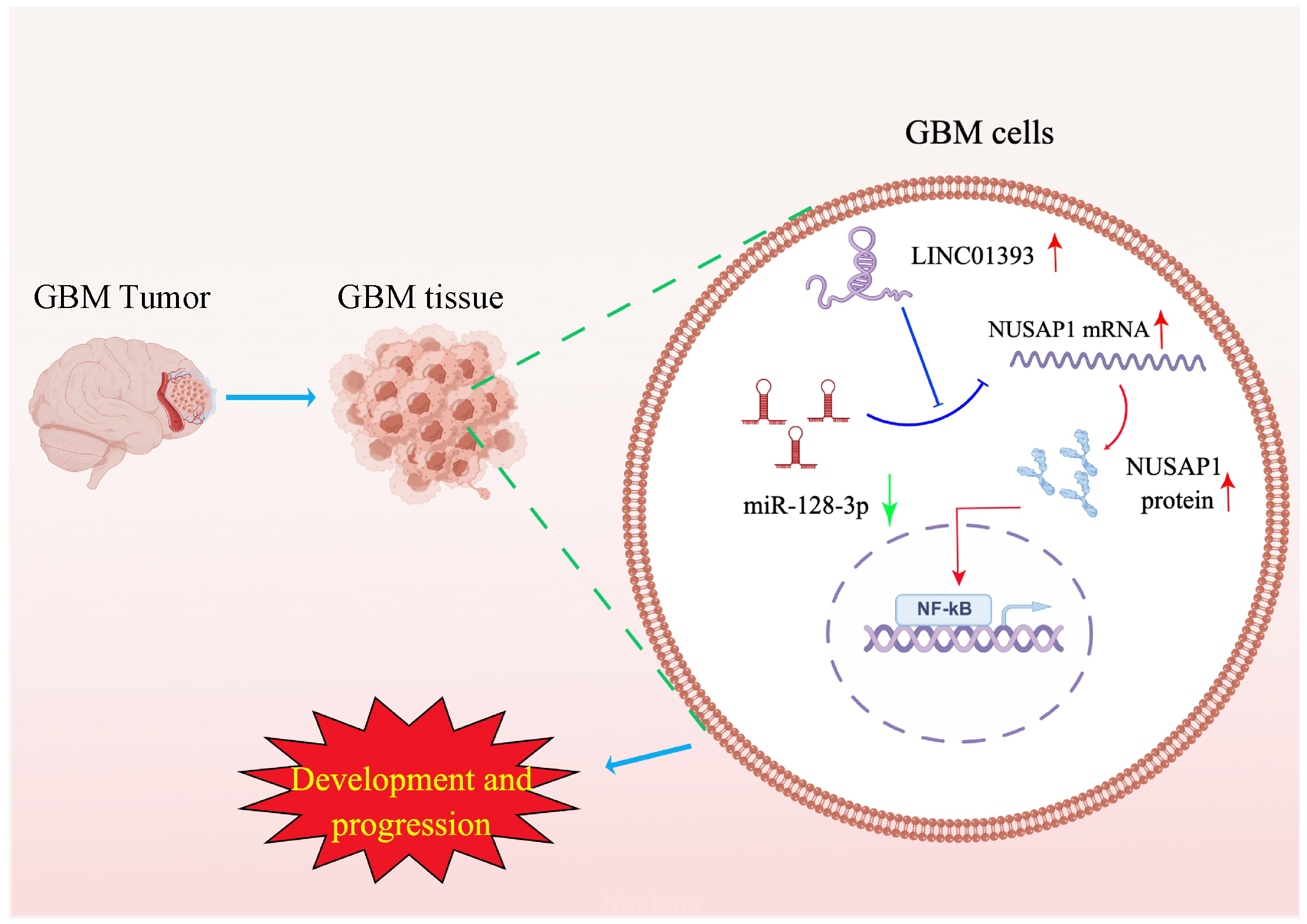
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bai, R.Y.; Staedtke, V.; Riggins, G.J. Molecular targeting of glioblastoma: Drug discovery and therapies. Trends Mol. Med. 2011, 17, 301–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stupp, R.; Taillibert, S.; Kanner, A.; Read, W.; Steinberg, D.; Lhermitte, B.; Toms, S.; Idbaih, A.; Ahluwalia, M.S.; Fink, K.; et al. Effect of tumor-treating fields plus maintenance temozolomide vs maintenance temozolomide alone on survival in patients with glioblastoma: A randomized clinical trial. JAMA 2017, 318, 2306–2316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U. Radiotherapy plus Concomitant and Adjuvant Temozolomide for Glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinot, O.L.; Wick, W.; Mason, W.; Henriksson, R.; Saran, F.; Nishikawa, R.; Carpentier, A.F.; Hoang-Xuan, K.; Kavan, P.; Cernea, D.; et al. Bevacizumab plus Radiotherapy–Temozolomide for Newly Diagnosed Glioblastoma. N. Engl. J. Med. 2014, 370, 709–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raemaekers, T.; Ribbeck, K.; Beaudouin, J.; Annaert, W.; Van Camp, M.; Stockmans, I.; Smets, N.; Bouillon, R.; Ellenberg, J.; Carmeliet, G. NuSAP, a novel microtubule-associated protein involved in mitotic spindle organization. J. Cell Biol. 2003, 162, 1017–1029. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Hooiveld, G.J.; Seehawer, M.; Caruso, S.; Heinzmann, F.; Schneider, A.T.; Frank, A.K.; Cardenas, D.V.; Sonntag, R.; Luedde, M.; et al. microRNA 193a-5p Regulates Levels of Nucleolar- and Spindle-Associated Protein 1 to Suppress Hepatocarcinogenesis. Gastroenterology 2018, 155, 1951–1966e.26. [Google Scholar] [CrossRef]
- Li, H.; Zhang, W.; Yan, M.; Qiu, J.; Chen, J.; Sun, X.; Chen, X.; Song, L.; Zhang, Y. Nucleolar and spindle associated protein 1 promotes metastasis of cervical carcinoma cells by activating Wnt/β-catenin signaling. J. Exp. Clin. Cancer Res. 2019, 38, 33. [Google Scholar] [CrossRef]
- Chen, L.; Yang, L.; Qiao, F.; Hu, X.; Li, S.; Yao, L.; Yang, X.-L.; Shao, Z.-M. High Levels of Nucleolar Spindle-Associated Protein and Reduced Levels of BRCA1 Expression Predict Poor Prognosis in Triple-Negative Breast Cancer. PLoS ONE 2015, 10, e0140572. [Google Scholar] [CrossRef]
- Bogunovic, D.; O’Neill, D.W.; Belitskaya-Levy, I.; Vacic, V.; Yu, Y.; Adams, S.; Darvishian, F.; Berman, R.; Shapiro, R.; Pavlick, A.C.; et al. Immune profile and mitotic index of metastatic melanoma lesions enhance clinical staging in predicting patient survival. Proc. Natl. Acad. Sci. USA 2009, 106, 20429–20434. [Google Scholar] [CrossRef] [Green Version]
- Wadia, P.P.; Coram, M.; Armstrong, R.J.; Mindrinos, M.; Butte, A.J.; Miklos, D.B. Antibodies specifically target AML antigen NuSAP1 after allogeneic bone marrow transplantation. Blood 2010, 115, 2077–2087. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; He, J.; Li, Y.; Lv, S.; Cui, H. NUSAP1 potentiates chemoresistance in glioblastoma through its SAP domain to stabilize ATR. Signal Transduct Target Ther. 2020, 5, 44. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Xue, Z.; Qiu, C.; Feng, Z.; Qi, Q.; Wang, J.; Jin, W.; Zhong, Z.; Liu, X.; Li, W.; et al. Knockdown of NUSAP1 inhibits cell proliferation and invasion through downregulation of TOP2A in human glioblastoma. Cell Cycle 2022, 21, 1842–1855. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xu, B.; Yang, C.; Wang, W.; Zhong, D.; Zhao, Z.; He, L.; Hu, Y.; Jiang, L.; Li, J.; et al. Nucleolar and spindle associated protein 1 promotes the aggressiveness of astrocytoma by activating the Hedgehog signaling pathway. J. Exp. Clin. Cancer Res. 2017, 36, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagano, T.; Fraser, P. No-Nonsense Functions for Long Noncoding RNAs. Cell 2011, 145, 178–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sana, J.; Faltejskova, P.; Svoboda, M.; Slaby, O. Novel classes of non-coding RNAs and cancer. J. Transl. Med. 2012, 10, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, J.; Liu, X.; Wang, P.; Xue, Y.; Ma, J.; Qu, C.; Liu, Y. RETRACTED: CRNDE Promotes Malignant Progression of Glioma by Attenuating miR-384/PIWIL4/STAT3 Axis. Mol. Ther. 2016, 24, 1199–1215. [Google Scholar] [CrossRef] [Green Version]
- Rahnama, S.; Bakhshinejad, B.; Farzam, F.; Bitaraf, A.; Ghazimoradi, M.H.; Babashah, S. Identification of dysregulated competing endogenous RNA networks in glioblastoma: A way toward improved therapeutic opportunities. Life Sci. 2021, 277, 119488. [Google Scholar] [CrossRef]
- Zhong, C.; Yu, Q.; Peng, Y.; Zhou, S.; Liu, Z.; Deng, Y.; Guo, L.; Zhao, S.; Chen, G. Novel LncRNA OXCT1-AS1 indicates poor prognosis and contributes to tumorigenesis by regulating miR-195/CDC25A axis in glioblastoma. J. Exp. Clin. Cancer Res. 2021, 40, 123. [Google Scholar] [CrossRef]
- Liao, K.; Lin, Y.; Gao, W.; Xiao, Z.; Medina, R.; Dmitriev, P.; Cui, J.; Zhuang, Z.; Zhao, X.; Qiu, Y.; et al. Blocking lncRNA MALAT1/miR-199a/ZHX1 Axis Inhibits Glioblastoma Proliferation and Progression. Mol. Nucleic Acids 2019, 18, 388–399. [Google Scholar] [CrossRef] [Green Version]
- Li, X.M.; Yu, Z.; Huai, M.; Han, H.Y.; Cao, S.S. miR-758-3p inhibits the proliferation and invasion of non-small cell lung cancer cells by targeting NUSAP1. Zhonghua Zhong Liu Za Zhi 2021, 43, 113–117. [Google Scholar] [CrossRef]
- Guo, X.; Li, Y.; Che, X.; Hou, K.; Qu, X.; Li, C. microRNA-569 inhibits tumor metastasis in pancreatic cancer by directly targeting NUSAP1. Aging 2022, 14, 3652–3665. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, W.; Kadier, A.; Zhang, H.; Yao, X. MicroRNA-769-5p suppresses cell growth and migration via targeting NUSAP1 in bladder cancer. J. Clin. Lab. Anal. 2020, 34, e23193. [Google Scholar] [CrossRef]
- Kang, Y.J.; Yang, D.C.; Kong, L.; Hou, M.; Meng, Y.Q.; Wei, L.; Gao, G. CPC2: A fast and accurate coding potential calculator based on sequence intrinsic features. Nucleic Acids Res. 2017, 45, W12–W16. [Google Scholar] [CrossRef] [Green Version]
- Blum, M.; Chang, H.Y.; Chuguransky, S.; Grego, T.; Kandasaamy, S.; Mitchell, A.; Nuka, G.; Paysan-Lafosse, T.; Qureshi, M.; Raj, S.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res. 2021, 49, D344–D354. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Guo, P.; Lin, Y.; Mao, Q.; Guo, L.; Ge, J.; Li, X.; Jiang, J.; Lin, X.; Qiu, Y. Role of glycosyltransferase PomGnT1 in glioblastoma progression. Neuro. Oncol. 2015, 17, 211–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Chen, F.; Ren, X.; Yang, Y.; Luo, J.; Yuan, J.; Tong, Q. RNA-Binding Protein COL14A1, TNS1, NUSAP1 and YWHAE Are Valid Biomarkers to Predict Peritoneal Metastasis in Gastric Cancer. Front. Oncol. 2022, 12, 830688. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Chen, G.; Qiao, X.; Chen, H.; Sun, L.; Ma, Q.; Li, N.; Wang, J.; Dai, C.; Xu, F. NUSAP1 Could be a Potential Target for Preventing NAFLD Progression to Liver Cancer. Front. Pharmacol. 2022, 13, 823140. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Hu, X.; Yun, X.; Liu, J.; Yang, J.; Tian, Z.; Zhang, X.; Zhang, Y.; Wang, X. Nucleolar and spindle associated protein 1 enhances chemoresistance through DNA damage repair pathway in chronic lymphocytic leukemia by binding with RAD51. Cell Death Dis. 2021, 12, 1083. [Google Scholar] [CrossRef] [PubMed]
- Gulzar, Z.G.; McKenney, J.K.; Brooks, J.D. Increased expression of NuSAP in recurrent prostate cancer is mediated by E2F1. Oncogene 2013, 32, 70–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kretschmer, C.; Sterner-Kock, A.; Siedentopf, F.; Schoenegg, W.; Schlag, P.M.; Kemmner, W. Identification of early molecular markers for breast cancer. Mol. Cancer 2011, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Zou, J.; Zhou, L.; Zhong, M.; He, Y.; Huang, S.; Chen, J.; Li, J.; Xiong, J.; Fang, Z.; et al. NUSAP1 Promotes Gastric Cancer Tumorigenesis and Progression by Stabilizing the YAP1 Protein. Front. Oncol. 2010, 10, 591698. [Google Scholar] [CrossRef]
- Fujiwara, T.; Harigae, H.; Okitsu, Y.; Takahashi, S.; Yokoyama, H.; Yamada, M.F.; Ishizawa, K.; Kameoka, J.; Kaku, M.; Sasaki, T. Expression analyses and transcriptional regulation of mouse nucleolar spindle-associated protein gene in erythroid cells: Essential role of NF-Y. Br. J. Haematol. 2006, 135, 583–590. [Google Scholar] [CrossRef]
- Hussain, S.; Benavente, S.B.; Nascimento, E.; Dragoni, I.; Kurowski, A.; Gillich, A.; Humphreys, P.; Frye, M. The nucleolar RNA methyltransferase Misu (NSun2) is required for mitotic spindle stability. J. Cell Biol. 2009, 186, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tay, Y.; Rinn, J.; Pandolfi, P.P. The multilayered complexity of ceRNA crosstalk and competition. Nature 2014, 505, 344–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Li, M.J.; Zhang, J.; Liang, Q.; Xuan, C.; Wu, J.; Jiang, P.; Li, W.; Zhu, Y.; Wang, P.; Fernandez, D.; et al. Exploring genetic associations with ceRNA regulation in the human genome. Nucleic Acids Res. 2017, 45, 5653–5665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grelet, S.; Link, L.A.; Howley, B.; Obellianne, C.; Palanisamy, V.; Gangaraju, V.K.; Diehl, J.A.; Howe, P.H. A regulated PNUTS mRNA to lncRNA splice switch mediates EMT and tumour progression. Nat. Cell Biol. 2017, 19, 1105–1115. [Google Scholar] [CrossRef] [Green Version]
- Godlewski, J.; Nowicki, M.O.; Bronisz, A.; Williams, S.; Otsuki, A.; Nuovo, G.; Raychaudhury, A.; Newton, H.B.; Chiocca, E.A.; Lawler, S. Targeting of the Bmi-1 Oncogene/Stem Cell Renewal Factor by MicroRNA-128 Inhibits Glioma Proliferation and Self-Renewal. Cancer Res. 2008, 68, 9125–9130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeOcesano-Pereira, C.; Machado, R.A.C.; Chudzinski-Tavassi, A.M.; Sogayar, M.C. Emerging Roles and Potential Applications of Non-Coding RNAs in Glioblastoma. Int. J. Mol. Sci. 2020, 21, 2611. [Google Scholar] [CrossRef] [Green Version]
- Papagiannakopoulos, T.; Friedmann-Morvinski, D.; Neveu, P.; Dugas, J.C.; Gill, R.M.; Huillard, E.; Liu, C.; Zong, H.; Rowitch, D.H.; Barres, B.A.; et al. Pro-neural miR-128 is a glioma tumor suppressor that targets mitogenic kinases. Oncogene 2012, 31, 1884–1895. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, M.; Candido, M.F.; Valera, E.T.; Brassesco, M.S. The multifaceted NF-kB: Are there still prospects of its inhibition for clinical intervention in pediatric central nervous system tumors? Cell Mol. Life Sci. 2021, 78, 6161–6200. [Google Scholar] [CrossRef] [PubMed]
- Friedmann-Morvinski, D.; Narasimamurthy, R.; Xia, Y.; Myskiw, C.; Soda, Y.; Verma, I.M. Targeting NF-κB in glioblastoma: A therapeutic approach. Sci. Adv. 2016, 2, e1501292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, K.P.L.; Balasubramaniyan, V.; Vaillant, B.; Ezhilarasan, R.; Hummelink, K.; Hollingsworth, F.; Wani, K.; Heathcock, L.; James, J.D.; Goodman, L.D.; et al. Mesenchymal Differentiation Mediated by NF-κB Promotes Radiation Resistance in Glioblastoma. Cancer Cell. 2013, 24, 331–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, S.B.; Cheng, Y.K.; Deng, R.; Deng, Y.; Li, P.; Zhang, L.; Lan, P. The Predictive Value of Estrogen Receptor 1 on Adjuvant Chemotherapy in Locally Advanced Colorectal Cancer: A Retrospective Analysis With Independent Validation and Its Potential Mechanism. Front. Oncol. 2020, 10, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Zhang, J.; Liu, Y.; Zhang, W.; Zhou, J.; Duan, R.; Pu, P.; Kang, C.; Han, L. A novel cell cycle-associated lncRNA, HOXA11-AS, is transcribed from the 5-prime end of the HOXA transcript and is a biomarker of progression in glioma. Cancer Lett. 2016, 373, 251–259. [Google Scholar] [CrossRef]
- Salagierski, M.; Verhaegh, G.W.; Jannink, S.A.; Smit, F.P.; Hessels, D.; Schalken, J.A. Differential expression of PCA3 and its overlapping PRUNE2 transcript in prostate cancer. Prostate 2010, 70, 70–78. [Google Scholar] [CrossRef]
- Wahlestedt, C. Targeting long non-coding RNA to therapeutically upregulate gene expression. Nat. Rev. Drug Discov. 2013, 12, 433–446. [Google Scholar] [CrossRef]
- Yoon, S.; Rossi, J.J. Therapeutic Potential of Small Activating RNAs (saRNAs) in Human Cancers. Curr. Pharm. Biotechnol. 2018, 19, 604–610. [Google Scholar] [CrossRef]
- Katsushima, K.; Natsume, A.; Ohka, F.; Shinjo, K.; Hatanaka, A.; Ichimura, N.; Sato, S.; Takahashi, S.; Kimura, H.; Totoki, Y.; et al. Targeting the Notch-regulated non-coding RNA TUG1 for glioma treatment. Nat. Commun. 2016, 7, 13616. [Google Scholar] [CrossRef]
- Ozes, A.R.; Wang, Y.; Zong, X.; Fang, F.; Pilrose, J.; Nephew, K.P. Therapeutic targeting using tumor specific peptides inhibits long non-coding RNA HOTAIR activity in ovarian and breast cancer. Sci. Rep. 2017, 7, 894. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Total (N) | Univariate Analysis | Multivariate Analysis | ||
|---|---|---|---|---|---|
| Hazard Ratio (95% CI) | p Value | Hazard Ratio (95% CI) | p Value | ||
| WHO grade | 614 | <0.001 | |||
| G2 | 215 | Reference | Reference | ||
| G3 | 239 | 3.066 (2.007–4.684) | <0.001 | 2.323 (1.412–3.820) | <0.001 |
| G4 | 160 | 19.142 (12.557–29.180) | <0.001 | 7.788 (2.476–24.501) | <0.001 |
| IDH status | 663 | <0.001 | |||
| WT | 237 | Reference | Reference | ||
| Mut | 426 | 0.101 (0.076–0.134) | <0.001 | 0.426 (0.249–0.729) | 0.002 |
| 1p/19q codeletion | 666 | <0.001 | |||
| Non-codel | 498 | Reference | Reference | ||
| Codel | 168 | 0.215 (0.138–0.337) | <0.001 | 0.765 (0.429–1.364) | 0.364 |
| Gender | 672 | 0.123 | |||
| Female | 283 | Reference | |||
| Male | 389 | 1.219 (0.946–1.570) | 0.126 | ||
| Age | 672 | <0.001 | |||
| ≤60 | 533 | Reference | Reference | ||
| >60 | 139 | 4.746 (3.633–6.201) | <0.001 | 4.071 (2.417–6.858) | <0.001 |
| PTO | 446 | <0.001 | |||
| PD | 103 | Reference | Reference | ||
| SD | 145 | 0.381 (0.250–0.580) | <0.001 | 0.419 (0.252–0.699) | <0.001 |
| PR | 63 | 0.135 (0.054–0.336) | <0.001 | 0.200 (0.072–0.558) | 0.002 |
| CR | 135 | 0.129 (0.062–0.269) | <0.001 | 0.197 (0.092–0.423) | <0.001 |
| LINC01393 | 672 | <0.001 | |||
| Low | 336 | Reference | Reference | ||
| High | 336 | 2.174 (1.684–2.808) | <0.001 | 1.759 (1.139–2.717) | 0.011 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Hu, J.; Li, S.; Zhou, C.; Feng, M.; Li, L.; Gao, Y.; Chen, X.; Wu, X.; Cao, Y.; et al. LINC01393, a Novel Long Non-Coding RNA, Promotes the Cell Proliferation, Migration and Invasion through MiR-128-3p/NUSAP1 Axis in Glioblastoma. Int. J. Mol. Sci. 2023, 24, 5878. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065878
Li D, Hu J, Li S, Zhou C, Feng M, Li L, Gao Y, Chen X, Wu X, Cao Y, et al. LINC01393, a Novel Long Non-Coding RNA, Promotes the Cell Proliferation, Migration and Invasion through MiR-128-3p/NUSAP1 Axis in Glioblastoma. International Journal of Molecular Sciences. 2023; 24(6):5878. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065878
Chicago/Turabian StyleLi, Deheng, Junda Hu, Sen Li, Changshuai Zhou, Mingtao Feng, Liangdong Li, Yang Gao, Xin Chen, Xiaojun Wu, Yiqun Cao, and et al. 2023. "LINC01393, a Novel Long Non-Coding RNA, Promotes the Cell Proliferation, Migration and Invasion through MiR-128-3p/NUSAP1 Axis in Glioblastoma" International Journal of Molecular Sciences 24, no. 6: 5878. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24065878