
Cirrhotic Cardiomyopathy Following Bile Duct Ligation in Rats—A Matter of Time?
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
2.1. Animal Model
2.2. Signs of Liver Injury and Cirrhosis
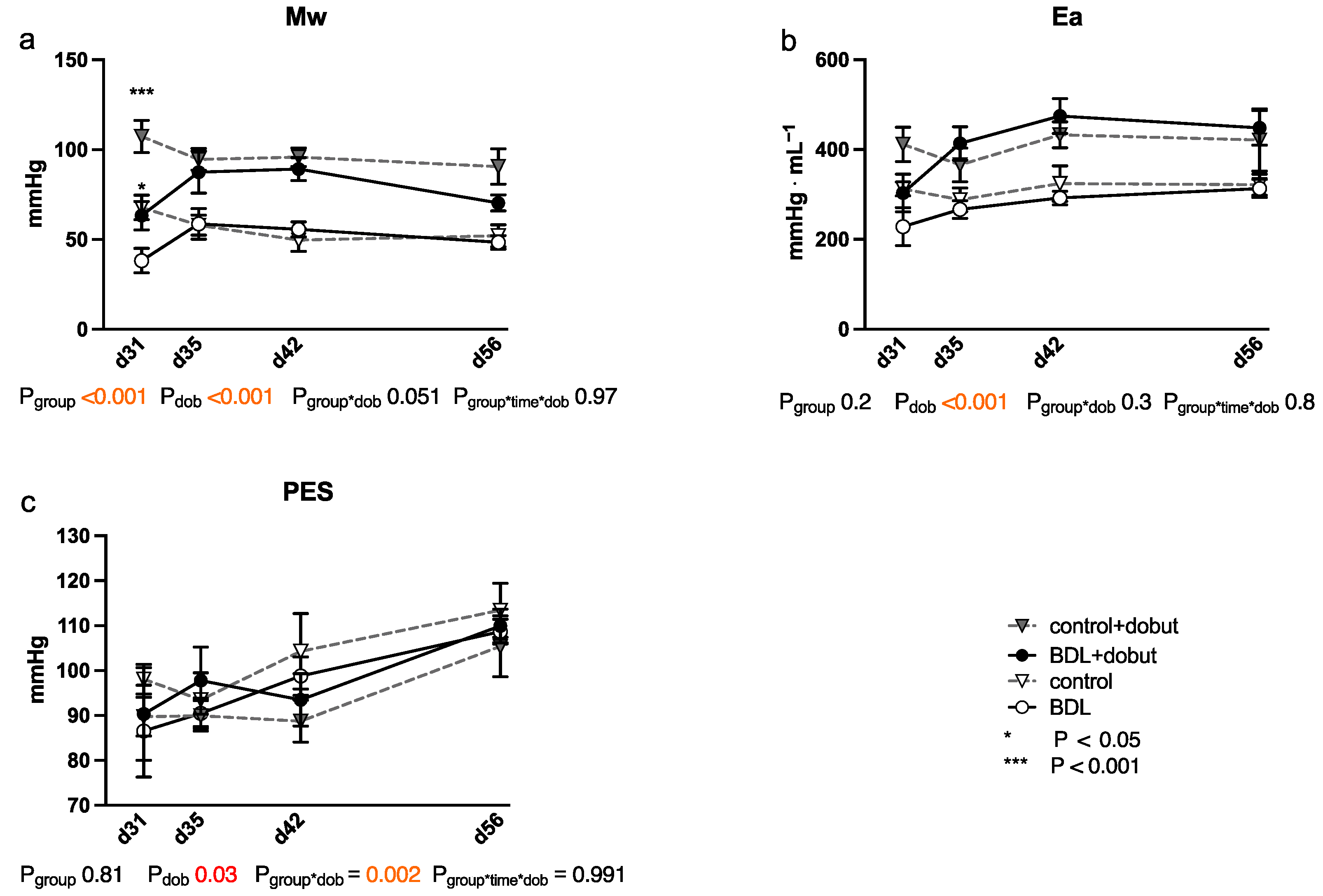
2.3. Impairment of Myocardial Function
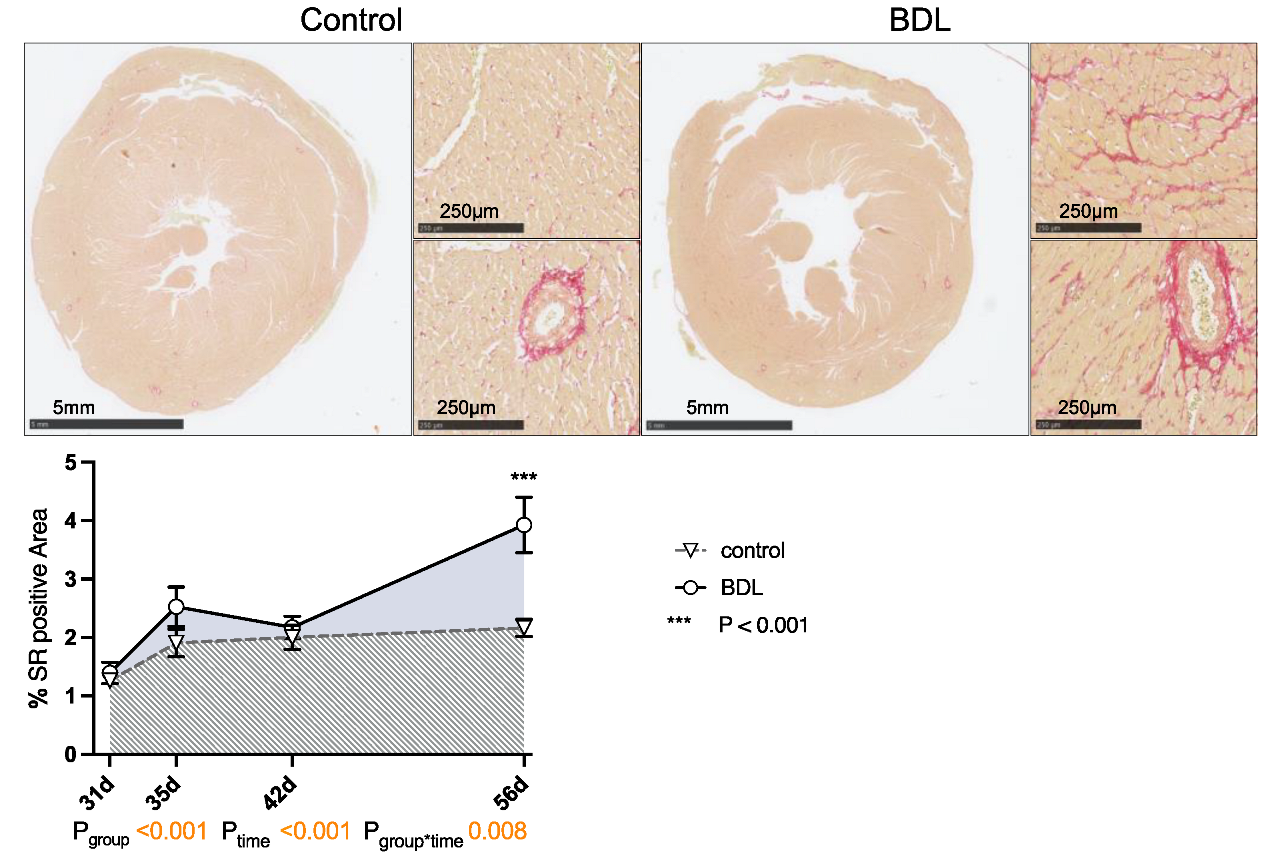
2.4. Cardiac Structure
2.5. Cardiac Injury and Inflammation
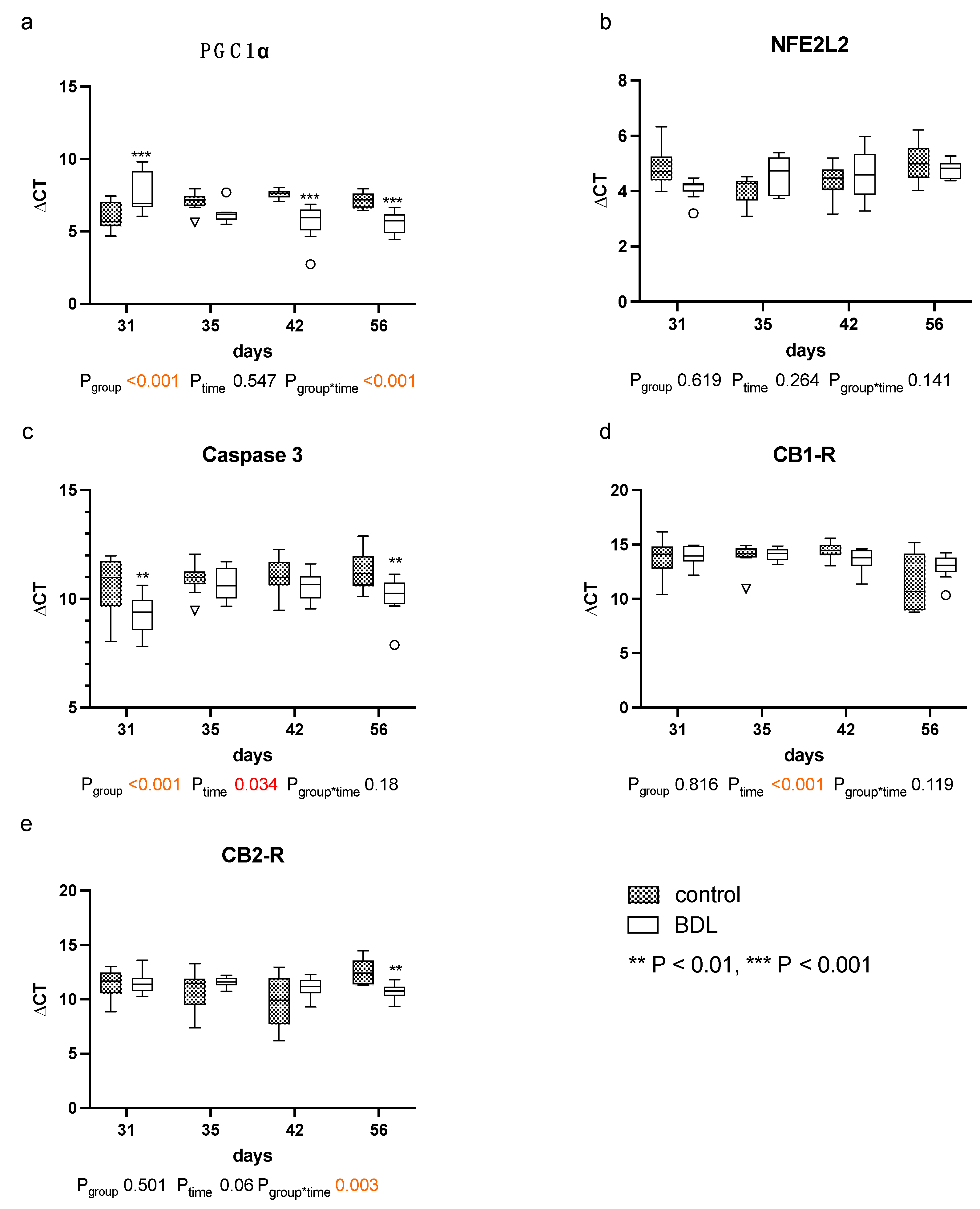
2.6. Myocardial Gene Expression
3. Discussion
4. Materials and Methods
4.1. Animal Model/Protocol
4.1.1. First Surgery
4.1.2. Second Surgery
4.1.3. Final Surgery
4.2. Hemodynamics
4.2.1. Basic Monitoring
4.2.2. Echocardiography
4.2.3. Pressure Volume Loops
4.3. Histological Staining
4.4. Serum Analysis
4.5. Quantitative Polymerase Chain Reaction
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2017 Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar]
- Asrani, S.K.; Larson, J.J.; Yawn, B.; Therneau, T.M.; Kim, W.R. Underestimation of Liver-Related Mortality in the United States. Gastroenterology 2013, 145, 375–382.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Amico, G.; Garcia-Tsao, G.; Pagliaro, L. Natural history and prognostic indicators of survival in cirrhosis: A systematic review of 118 studies. J. Hepatol. 2006, 44, 217–231. [Google Scholar] [CrossRef]
- Sørensen, H.T.; Thulstrup, A.M.; Mellemkjar, L.; Jepsen, P.; Christensen, E.; Olsen, J.H.; Vilstrup, H. Long-term survival and cause-specific mortality in patients with cirrhosis of the liver: A nationwide cohort study in Denmark. J. Clin. Epidemiol. 2003, 56, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, M.; Carugo, S.; Boari, G.; Pecci, V.; De Ceglia, S.; Maggiolini, S.; Mancia, G. Evidence of functional and structural cardiac abnormalities in cirrhotic patients with and without ascites. Hepatology 1997, 26, 1131–1137. [Google Scholar] [PubMed]
- Wong, F.; Girgrah, N.; Graba, J.; Allidina, Y.; Liu, P.; Blendis, L. The cardiac response to exercise in cirrhosis. Gut 2001, 49, 268–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Møller, S.; Hobolth, L.; Winkler, C.; Bendtsen, F.; Christensen, E. Determinants of the hyperdynamic circulation and central hypovolaemia in cirrhosis. Gut 2011, 60, 1254–1259. [Google Scholar] [CrossRef]
- Zardi, E.M.; Henriksen, J.H. Cirrhotic cardiomyopathy. J. Am. Coll Cardiol. 2010, 56, 539–549. [Google Scholar] [CrossRef] [Green Version]
- Møller, S.; Danielsen, K.V.; Wiese, S.; Hove, J.D.; Bendtsen, F. An update on cirrhotic cardiomyopathy. Expert Rev. Gastroenterol. Hepatol. 2019, 13, 497–505. [Google Scholar] [CrossRef]
- Izzy, M.; VanWagner, L.B.; Lin, G.; Altieri, M.; Findlay, J.Y.; Oh, J.K.; Watt, K.D.; Lee, S.S.; on behalf of The Cirrhotic Cardiomyopathy Consortium. Redefining Cirrhotic Cardiomyopathy for the Modern Era. Hepatology 2020, 71, 334–345. [Google Scholar] [CrossRef]
- Spann, A.; Coe, C.; Ajayi, T.; Montgomery, G.; Shwetar, M.; Oje, A.; Annis, J.; Slaughter, J.C.; Alexopoulos, S.; Brittain, E.; et al. Cirrhotic cardiomyopathy: Appraisal of the original and revised criteria in predicting posttransplant cardiac outcomes. Liver Transplant. 2022, 28, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Mechelinck, M.; Hartmann, B.; Hamada, S.; Becker, M.; Andert, A.; Ulmer, T.F.; Neumann, U.P.; Wirtz, T.H.; Koch, A.; Trautwein, C.; et al. Global Longitudinal Strain at Rest as an Independent Predictor of Mortality in Liver Transplant Candidates: A Retrospective Clinical Study. J. Clin. Med. 2020, 9, 2616. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.S. Mechanistic insights into the pathophysiology of cirrhotic cardiomyopathy. Anal. Biochem. 2021, 636, 114388. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Kim, H.K.; Lee, J.H.; Lee, Y.B.; Park, E.A.; Park, J.B.; Sohn, D.W. Myocardial structural and functional changes in patients with liver cirrhosis awaiting liver transplantation: A comprehensive cardiovascular magnetic resonance and echocardiographic study. J. Cardiovasc. Magn. Reson. 2020, 22, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bátkai, S.; Mukhopadhyay, P.; Harvey-White, J.; Kechrid, R.; Pacher, P.; Kunos, G. Endocannabinoids acting at CB1 receptors mediate the cardiac contractile dysfunction in vivo in cirrhotic rats. Am. J. Physiol. Circ. Physiol. 2007, 293, H1689–H1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaskari, S.A.; Liu, H.; D’Mello, C.; Kunos, G.; Lee, S.S. Blunted cardiac response to hemorrhage in cirrhotic rats is mediated by local macrophage-released endocannabinoids. J. Hepatol. 2015, 62, 1272–1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Kim, K.H.; Schilling, J.M.; Leem, J.; Dhanani, M.; Head, B.P.; Roth, D.M.; Zemljic-Harpf, A.E.; Patel, H.H. Protective role of cardiac-specific overexpression of caveolin-3 in cirrhotic cardiomyopathy. Am. J. Physiol. Liver Physiol. 2020, 318, G531–G541. [Google Scholar] [CrossRef]
- Desai, M.S.; Shabier, Z.; Taylor, M.; Lam, F.; Thevananther, S.; Kosters, A.; Karpen, S.J. Hypertrophic cardiomyopathy and dysregulation of cardiac energetics in a mouse model of biliary fibrosis. Hepatology 2010, 51, 2097–2107. [Google Scholar] [CrossRef] [Green Version]
- Matyas, C.; Erdelyi, K.; Trojnar, E.; Zhao, S.; Varga, Z.V.; Paloczi, J.; Mukhopadhyay, P.; Nemeth, B.T.; Haskó, G.; Cinar, R.; et al. Interplay of Liver–Heart Inflammatory Axis and Cannabinoid 2 Receptor Signaling in an Experimental Model of Hepatic Cardiomyopathy. Hepatology 2019, 71, 1391–1407. [Google Scholar] [CrossRef]
- Nam, S.W.; Liu, H.; Wong, J.Z.; Feng, A.Y.; Chu, G.; Merchant, N.; Lee, S.S. Cardiomyocyte apoptosis contributes to pathogenesis of cirrhotic cardiomyopathy in bile duct-ligated mice. Clin. Sci. 2014, 127, 519–526. [Google Scholar] [CrossRef]
- Kökény, G.; Calvier, L.; Legchenko, E.; Chouvarine, P.; Mózes, M.M.; Hansmann, G. PPARgamma is a gatekeeper for extracellular matrix and vascular cell homeostasis: Beneficial role in pulmonary hypertension and renal/cardiac/pulmonary fibrosis. Curr. Opin. Nephrol. Hypertens 2020, 29, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Schirone, L.; Forte, M.; Palmerio, S.; Yee, D.; Nocella, C.; Angelini, F.; Pagano, F.; Schiavon, S.; Bordin, A.; Carrizzo, A.; et al. A Review of the Molecular Mechanisms Underlying the Development and Progression of Cardiac Remodeling. Oxidative Med. Cell Longev. 2017, 2017, 3920195. [Google Scholar] [CrossRef] [PubMed]
- Ward, C.A.; Liu, H.; Lee, S.S. Altered cellular calcium regulatory systems in a rat model of cirrhotic cardiomyopathy. Gastroenterology 2001, 121, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Gregolin, C.S.; Nascimento, M.D.; de Souza, S.L.B.; Mota, G.A.F.; Bomfim, G.F.; Luvizotto, R.D.A.M.; Sugizaki, M.M.; Bazan, S.G.Z.; de Campos, D.H.S.; Dias, M.C.; et al. Myocardial Dysfunction in Cirrhotic Cardiomyopathy is Associated with Alterations of Phospholamban Phosphorylation and IL-6 Levels. Arch. Med. Res. 2020, 52, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Lubel, J.S.; Herath, C.B.; Burrell, L.M.; Angus, P.W. Liver disease and the renin-angiotensin system: Recent discoveries and clinical implications. J. Gastroenterol. Hepatol. 2008, 23, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Warner, F.J.; Lubel, J.S.; McCaughan, G.W.; Angus, P.W. Liver fibrosis: A balance of ACEs? Clin. Sci. 2007, 113, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.S.; Penny, D.J. Bile acids induce arrhythmias: Old metabolite, new tricks. Heart 2013, 99, 1629–1630. [Google Scholar] [CrossRef] [Green Version]
- Joubert, P. Cholic acid and the heart: In vitro studies of the effect on heart rate and myocardial contractility in the rat. Clin. Exp. Pharmacol. Physiol. 1978, 5, 9–16. [Google Scholar] [CrossRef]
- Makishima, M.; Okamoto, A.Y.; Repa, J.J.; Tu, H.; Learned, R.M.; Luk, A.; Hull, M.V.; Lustig, K.D.; Mangelsdorf, D.J.; Shan, B. Identification of a Nuclear Receptor for Bile Acids. Science 1999, 284, 1362–1365. [Google Scholar] [CrossRef]
- Kawamata, Y.; Fujii, R.; Hosoya, M.; Harada, M.; Yoshida, H.; Miwa, M.; Fukusumi, S.; Habata, Y.; Itoh, T.; Shintani, Y.; et al. A G Protein-coupled Receptor Responsive to Bile Acids. J. Biol. Chem. 2003, 278, 9435–9440. [Google Scholar] [CrossRef] [Green Version]
- Pu, J.; Yuan, A.; Shan, P.; Gao, E.; Wang, X.; Wang, Y.; Lau, W.B.; Koch, W.; Ma, X.-L.; Ben He, B. Cardiomyocyte-expressed farnesoid-X-receptor is a novel apoptosis mediator and contributes to myocardial ischaemia/reperfusion injury. Eur. Heart J. 2012, 34, 1834–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, M.S.; Mathur, B.; Eblimit, Z.; Vasquez, H.; Taegtmeyer, H.; Karpen, S.J.; Penny, D.J.; Moore, D.D.; Anakk, S. Bile acid excess induces cardiomyopathy and metabolic dysfunctions in the heart. Hepatology 2016, 65, 189–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, M.; Wang, S.C.; Shirai, M.; Scaglia, F.; Xie, M.; Sakai, S.; Tanaka, T.; Kulkarni, P.A.; Barger, P.M.; Youker, K.A.; et al. Activation of cardiac Cdk9 represses PGC-1 and confers a predisposition to heart failure. EMBO J. 2004, 23, 3559–3569. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, I.; Tharakan, B.; Bhat, G. Current concepts in apoptosis: The physiological suicide program revisited. Cell Mol. Biol. Lett. 2006, 11, 506–525. [Google Scholar] [CrossRef]
- Chen, X.; Andresen, B.; Hill, M.; Zhang, J.; Booth, F.; Zhang, C. Role of Reactive Oxygen Species in Tumor Necrosis Factor-alpha Induced Endothelial Dysfunction. Curr. Hypertens. Rev. 2008, 4, 245–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze-Osthoff, K.; Bakker, A.C.; Vanhaesebroeck, B.; Beyaert, R.; Jacob, W.A.; Fiers, W. Cytotoxic activity of tumor necrosis factor is mediated by early damage of mitochondrial functions. Evidence for the involvement of mitochondrial radical generation. J. Biol. Chem. 1992, 267, 5317–5323. [Google Scholar] [CrossRef] [PubMed]
- Garnier, A.; Fortin, D.; Deloménie, C.; Momken, I.; Veksler, V.; Ventura-Clapier, R. Depressed mitochondrial transcription factors and oxidative capacity in rat failing cardiac and skeletal muscles. J. Physiol. 2003, 551, 491–501. [Google Scholar] [CrossRef]
- Northup, P.G.; Wanamaker, R.C.; Lee, V.D.; Adams, R.B.; Berg, C.L. Model for End-Stage Liver Disease (MELD) Predicts Nontransplant Surgical Mortality in Patients with Cirrhosis. Ann. Surg. 2005, 242, 244–251. [Google Scholar] [CrossRef]
- Millwala, F. Outcomes of patients with cirrhosis undergoing non-hepatic surgery: Risk assessment and management. World J. Gastroenterol. 2007, 13, 4056–4063. [Google Scholar] [CrossRef] [Green Version]
- Newman, K.L.; Johnson, K.M.; Cornia, P.B.; Wu, P.; Itani, K.; Ioannou, G.N. Perioperative Evaluation and Management of Patients with Cirrhosis: Risk Assessment, Surgical Outcomes, and Future Directions. Clin. Gastroenterol. Hepatol. 2019, 18, 2398–2414.e3. [Google Scholar] [CrossRef]
- Westermann, D.; Van Linthout, S.; Dhayat, S.; Dhayat, N.; Schmidt, A.; Noutsias, M.; Song, X.-Y.; Spillmann, F.; Riad, A.; Schultheiss, H.-P.; et al. Tumor necrosis factor-alpha antagonism protects from myocardial inflammation and fibrosis in experimental diabetic cardiomyopathy. Basic Res. Cardiol. 2007, 102, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Honold, L.; Nahrendorf, M. Resident and Monocyte-Derived Macrophages in Cardiovascular Disease. Circ. Res. 2018, 122, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Frangogiannis, N.G. Immune cells in repair of the infarcted myocardium. Microcirculation 2017, 24, e12305. [Google Scholar] [CrossRef] [PubMed]
- Frangogiannis, N.G. Cardiac fibrosis: Cell biological mechanisms, molecular pathways and therapeutic opportunities. Mol. Asp. Med. 2019, 65, 70–99. [Google Scholar] [CrossRef]
- Tag, C.G.; Weiskirchen, S.; Hittatiya, K.; Tacke, F.; Tolba, R.H. Induction of experimental obstructive cholestasis in mice. Lab. Anim. 2015, 49, 70–80. [Google Scholar] [CrossRef]
- Inserte, J.; Perelló, A.; Agulló, L.; Ruiz-Meana, M.; Schlüter, K.; Escalona, N.; Graupera, M.; Bosch, J.; Garcia-Dorado, D. Left ventricular hypertrophy in rats with biliary cirrhosis. Hepatology 2003, 38, 589–598. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, Y.; Huet, P.M.; Lee, S.S. Differential effects of jaundice and cirrhosis on beta-adrenoceptor signaling in three rat models of cirrhotic cardiomyopathy. J. Hepatol. 1999, 30, 485–491. [Google Scholar] [CrossRef]
- Mukhopadhyay, P.; Rajesh, M.; Cao, Z.; Horváth, B.; Park, O.; Wang, H.; Erdelyi, K.; Holovac, E.; Wang, Y.; Liaudet, L.; et al. Poly (ADP-ribose) polymerase-1 is a key mediator of liver inflammation and fibrosis. Hepatology 2013, 59, 1998–2009. [Google Scholar] [CrossRef] [Green Version]
- Varga, Z.V.; Erdelyi, K.; Paloczi, J.; Cinar, R.; Zsengeller, Z.K.; Jourdan, T.; Matyas, C.; Nemeth, B.T.; Guillot, A.; Xiang, X.; et al. Disruption of Renal Arginine Metabolism Promotes Kidney Injury in Hepatorenal Syndrome in Mice. Hepatology 2018, 68, 1519–1533. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Kannan, N.; Talwar, R.; Tyagi, A.K.; Madan, R.; Jaiswal, P.; Kumar, A. Suture granuloma: A rare differential diagnosis of residual/recurrent gastrointestinal stromal tumor of stomach. Int. Cancer Conf. J. 2015, 5, 5–8. [Google Scholar] [CrossRef]
- Panilaitis, B.; Altman, G.H.; Chen, J.; Jin, H.-J.; Karageorgiou, V.; Kaplan, D.L. Macrophage responses to silk. Biomaterials 2003, 24, 3079–3085. [Google Scholar] [CrossRef] [PubMed]
- Triantafyllou, E.; Woollard, K.J.; McPhail, M.J.W.; Antoniades, C.G.; Possamai, L.A. The Role of Monocytes and Macrophages in Acute and Acute-on-Chronic Liver Failure. Front. Immunol. 2018, 9, 2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woolbright, B.L.; Jaeschke, H. Inflammation and Cell Death During Cholestasis: The Evolving Role of Bile Acids. Gene Expr. 2019, 19, 215–228. [Google Scholar] [CrossRef]
- Sabbah, H.N. Targeting the Mitochondria in Heart Failure: A Translational Perspective. JACC Basic Transl. Sci. 2020, 5, 88–106. [Google Scholar] [CrossRef]
- Wang, F.; Yang, J.; Sun, J.; Dong, Y.; Zhao, H.; Shi, H.; Fu, L. Testosterone replacement attenuates mitochondrial damage in a rat model of myocardial infarction. J. Endocrinol. 2015, 225, 101–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, M.; Coxito, P.M.; Sardão, V.A.; Palmeira, C.M.; Oliveira, P.J. Bile acids are toxic for isolated cardiac mitochondria: A possible cause for hepatic-derived cardiomyopathies? Cardiovasc. Toxicol. 2005, 5, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Di Sessa, A.; Umano, G.R.; Del Giudice, E.M.; Santoro, N. From the liver to the heart: Cardiac dysfunction in obese children with non-alcoholic fatty liver disease. World J. Hepatol. 2017, 9, 69–73. [Google Scholar] [CrossRef]
- Junge, N.; Junge, C.; Schröder, J.; Pfister, E.; Leiskau, C.; Hohmann, D.; Beerbaum, P.; Baumann, U. Pediatric cirrhotic cardiomyopathy: Impact on liver transplant outcomes. Liver Transplant. 2018, 24, 820–830. [Google Scholar] [CrossRef] [Green Version]
- Russell, W.M.S.; Burch, R.L.; Hume, C.W. The Principles of Humane Experimental Technique; Methuen: London, UK, 1959; Volume 238. [Google Scholar]
- Federal Office Of Justice. Animal Protection Act (TierSchG); Federal Office of Justice: Bonn, Germany, 1972; Available online: https://www.gesetze-im-internet.de/tierschg/BJNR012770972.html (accessed on 5 February 2019).
- Krueger, J.C.; Habigt, M.A.; Helmedag, M.J.; Uhlig, M.; Moss, M.; Bleich, A.; Tolba, R.H.; Rossaint, R.; Hein, M.; Mechelinck, M. Evaluation of score parameters for severity assessment of surgery and liver cirrhosis in rats. Anim. Welf. 2023, 32, e29. [Google Scholar] [CrossRef]
- Tag, C.G.; Sauer-Lehnen, S.; Weiskirchen, S.; Borkham-Kamphorst, E.; Tolba, R.H.; Tacke, F.; Weiskirchen, R. Bile duct ligation in mice: Induction of inflammatory liver injury and fibrosis by obstructive cholestasis. J. Vis. Exp. 2015, 96, e52438. [Google Scholar]
- Mátyás, C.; Kovács, A.; Németh, B.T.; Oláh, A.; Braun, S.; Tokodi, M.; Barta, B.A.; Benke, K.; Ruppert, M.; Lakatos, B.K.; et al. Comparison of speckle-tracking echocardiography with invasive hemodynamics for the detection of characteristic cardiac dysfunction in type-1 and type-2 diabetic rat models. Cardiovasc. Diabetol. 2018, 17, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roehl, A.B.; Funcke, S.; Becker, M.M.; Goetzenich, A.; Bleilevens, C.; Rossaint, R.; Steendijk, P.; Hein, M. Xenon and Isoflurane Reduce Left Ventricular Remodeling after Myocardial Infarction in the Rat. Anesthesiology 2013, 118, 1385–1394. [Google Scholar] [CrossRef] [Green Version]
- Antonini-Canterin, F.; Pavan, D.; Di Bello, V.; Nicolosi, G.L.; Poli, S.; Vriz, O. The ventricular-arterial coupling: From basic pathophysiology to clinical application in the echocardiography laboratory. J. Cardiovasc. Echogr. 2013, 23, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef] [Green Version]
- Morkin, E. Control of cardiac myosin heavy chain gene expression. Microsc. Res. Tech. 2000, 50, 522–531. [Google Scholar] [CrossRef]
- Fu, S.; Ping, P.; Wang, F.; Luo, L. Synthesis, secretion, function, metabolism and application of natriuretic peptides in heart failure. J. Biol. Eng. 2018, 12, 2. [Google Scholar] [CrossRef] [Green Version]
- Maruyama, K.; Imanaka-Yoshida, K. The Pathogenesis of Cardiac Fibrosis: A Review of Recent Progress. Int. J. Mol. Sci. 2022, 23, 2617. [Google Scholar] [CrossRef] [PubMed]
- Zhihao, L.; Jingyu, N.; Lan, L.; Michael, S.; Rui, G.; Xiyun, B.; Xiaozhi, L.; Guanwei, F. SERCA2a: A key protein in the Ca2+ cycle of the heart failure. Heart Fail. Rev. 2019, 25, 523–535. [Google Scholar] [CrossRef]
- Gaskari, S.A.; Liu, H.; Moezi, L.; Li, Y.; Baik, S.K.; Lee, S.S. Role of endocannabinoids in the pathogenesis of cirrhotic cardiomyopathy in bile duct-ligated rats. Br. J. Pharmacol. 2005, 146, 315–323. [Google Scholar] [CrossRef]
- Arany, Z.; He, H.; Lin, J.; Hoyer, K.; Handschin, C.; Toka, O.; Ahmad, F.; Matsui, T.; Chin, S.; Wu, P.-H.; et al. Transcriptional coactivator PGC-1α controls the energy state and contractile function of cardiac muscle. Cell Metab. 2005, 1, 259–271. [Google Scholar] [CrossRef] [Green Version]
- Kasai, S.; Shimizu, S.; Tatara, Y.; Mimura, J.; Itoh, K. Regulation of Nrf2 by Mitochondrial Reactive Oxygen Species in Physiology and Pathology. Biomolecules 2020, 10, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curran-Everett, D. Multiple comparisons: Philosophies and illustrations. Am. J. Physiol. Integr. Comp. Physiol. 2000, 279, R1–R8. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Days after BDL | 28 Days | 31 Days | 35 Days | 42 Days | 56 Days | Ptime | Pgroup*time | Pgroup | |
|---|---|---|---|---|---|---|---|---|---|
| n Control/BDL | n = 40/39 | n = 10/10 | n = 10/9 | n = 10/10 | n = 10/10 | ||||
| CK | Control | 221 ± 28 | 283 ± 74 | 243 ± 30 | 275 ± 59 | 416 ± 22 | 0.055 | 0.58 | 0.26 |
| BDL | 204 ± 22 | 266 ± 40 | 418 ± 204 | 194 ± 30 | 234 ± 81 | ||||
| Pgroup | 0.99 | 0.99 | 0.95 | 0.81 | 0.60 | ||||
| CKMB | Control | 262 ± 26 | 321 ± 80 | 262 ± 66 | 358 ± 94 | 445 ± 113 | 0.053 | 0.91 | 0.34 |
| BDL | 193 ± 19 | 248 ± 53 | 234 ± 76 | 280 ± 53 | 462 ± 145 | ||||
| Pgroup | 0.15 | 0.93 | 0.99 | 0.99 | 0.99 | ||||
| Crea | Control | 33.3 ± 1.2 | 45 ± 4.4 | 36.1 ± 3 | 31 ± 2.4 | 34.1 ± 2.5 | non-parametric test | non-parametric test | non-parametric test |
| BDL | 40 ± 1.6 | 52.9 ± 6.7 | 43.3 ± 2.2 | 37.5 ± 1.7 | 44.4 ± 2.3 | ||||
| Pgroup | 0.006 | 0.53 | 0.024 | 0.013 | 0.013 | ||||
| WBC | Control | 11 ± 1 | 10 ± 1 | 11 ± 1 | 9 ± 1 | 9 ± 1 | 0.79 | 0.28 | <0.001 |
| BDL | 26 ± 2 | 24 ± 2 | 28 ± 7 | 28 ± 4 | 35 ± 5 | ||||
| Pgroup | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||
| TNFa | Control | 2.21 ± 0.35 | 0.89 ± 0.36 | 1.25 ± 0.38 | 0.05 ± 0 | 1.22 ± 0.83 | 0.01 | <0.001 | <0.001 |
| BDL | 4.4 ± 0.36 | 7.22 ± 2.76 | 5.7 ± 0.97 | 8.12 ± 2.73 | 20.1 ± 3.9 | ||||
| Pgroup | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uhlig, M.; Hein, M.; Habigt, M.A.; Tolba, R.H.; Braunschweig, T.; Helmedag, M.J.; Arici, M.; Theißen, A.; Klinkenberg, A.; Klinge, U.; et al. Cirrhotic Cardiomyopathy Following Bile Duct Ligation in Rats—A Matter of Time? Int. J. Mol. Sci. 2023, 24, 8147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24098147
Uhlig M, Hein M, Habigt MA, Tolba RH, Braunschweig T, Helmedag MJ, Arici M, Theißen A, Klinkenberg A, Klinge U, et al. Cirrhotic Cardiomyopathy Following Bile Duct Ligation in Rats—A Matter of Time? International Journal of Molecular Sciences. 2023; 24(9):8147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24098147
Chicago/Turabian StyleUhlig, Moritz, Marc Hein, Moriz A. Habigt, René H. Tolba, Till Braunschweig, Marius J. Helmedag, Melissa Arici, Alexander Theißen, Axel Klinkenberg, Uwe Klinge, and et al. 2023. "Cirrhotic Cardiomyopathy Following Bile Duct Ligation in Rats—A Matter of Time?" International Journal of Molecular Sciences 24, no. 9: 8147. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24098147