

Influences of the Integrated Rice-Crayfish Farming System with Different Stocking Densities on the Paddy Soil Microbiomes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
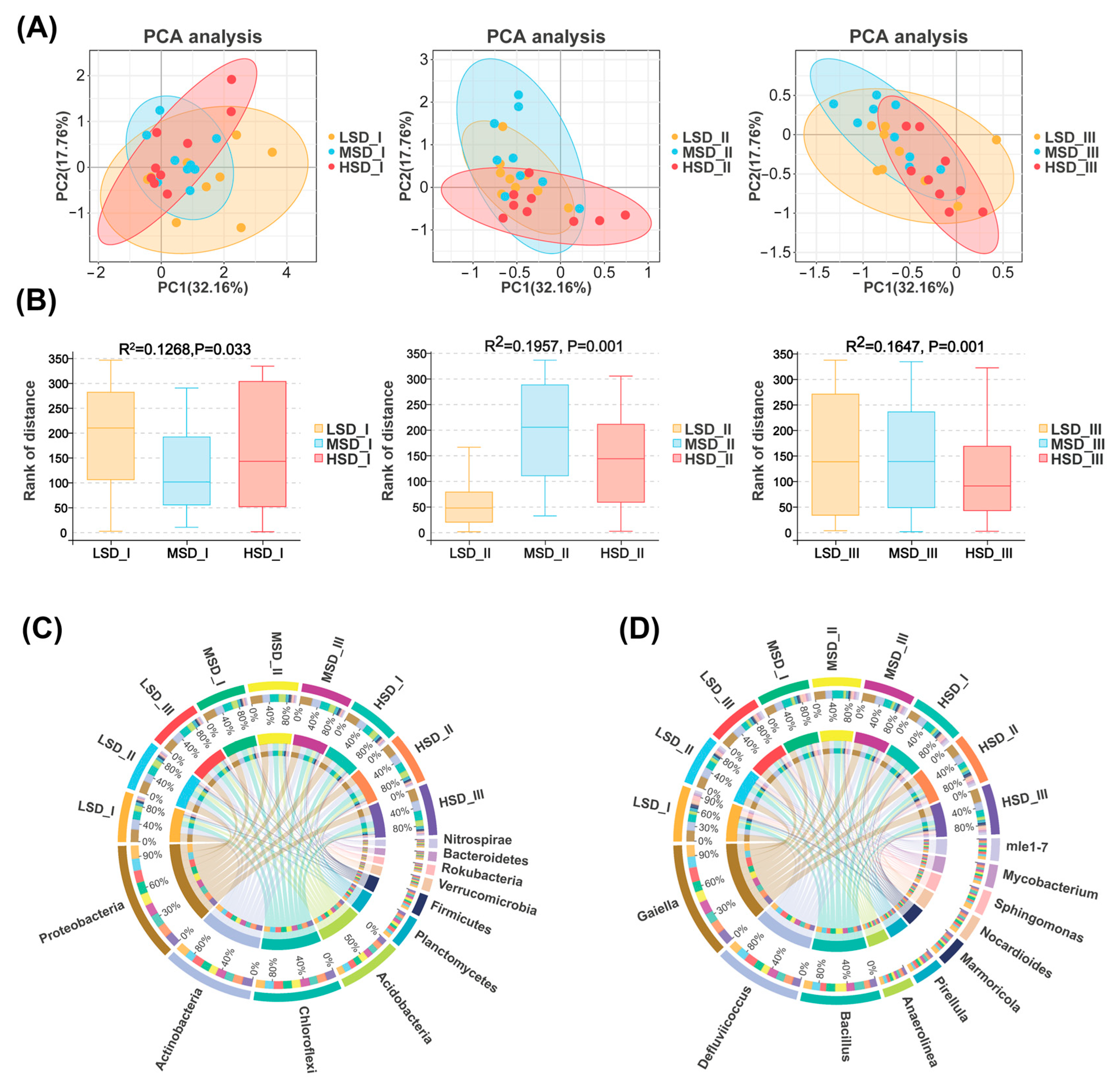
2.1. The Bacterial Community Diversity within Paddy Soil
2.2. The Bacterial Community Composition within Paddy Soil
2.3. Function Predictions for the Soil Bacterial Community
2.4. Co-Occurrence Network Patterns for the Soil Bacterial Community
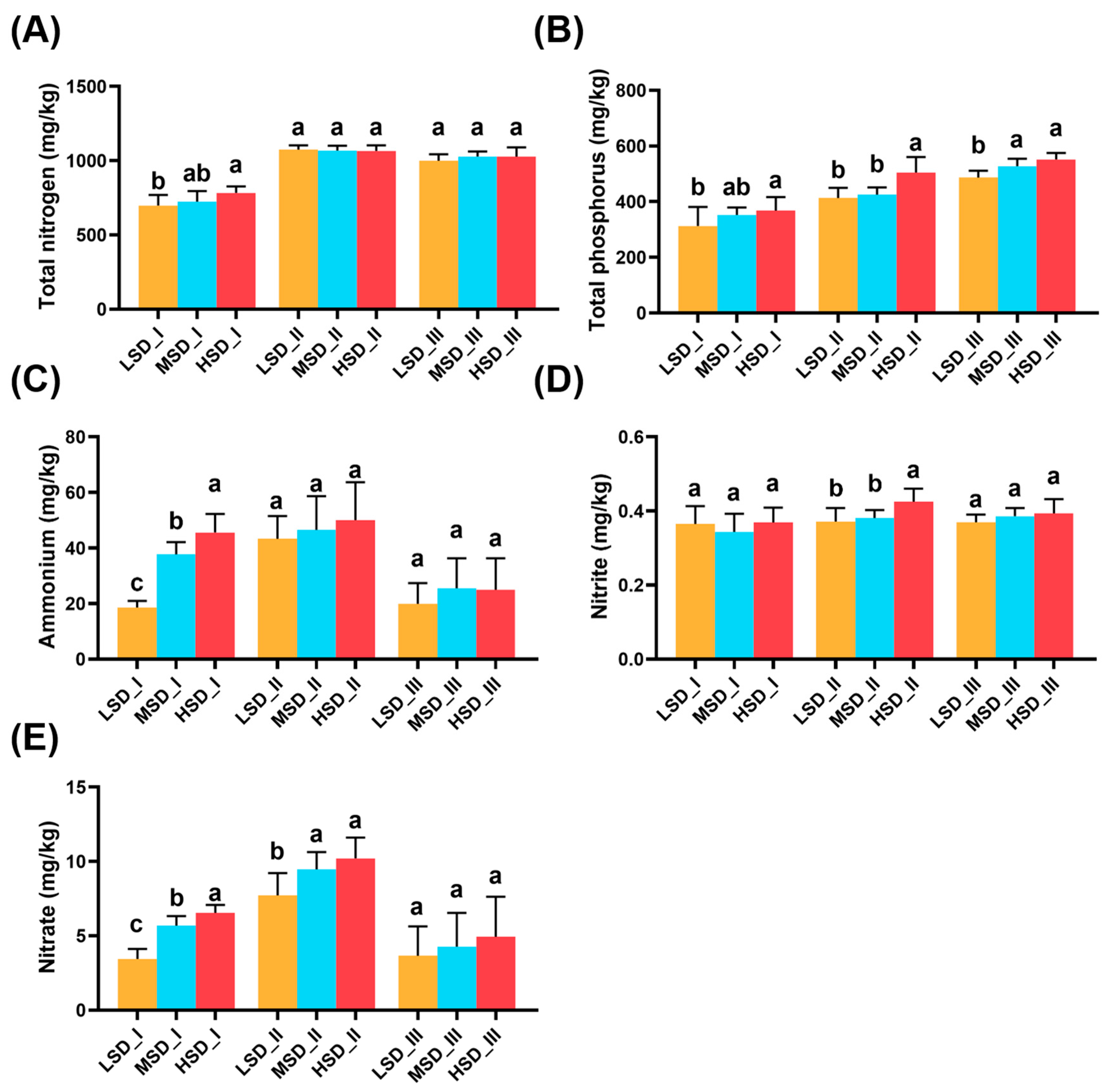
2.5. Variations in the Environmental Factors and Their Relationships with the Soil Bacterial Community
3. Discussion
4. Materials and Methods
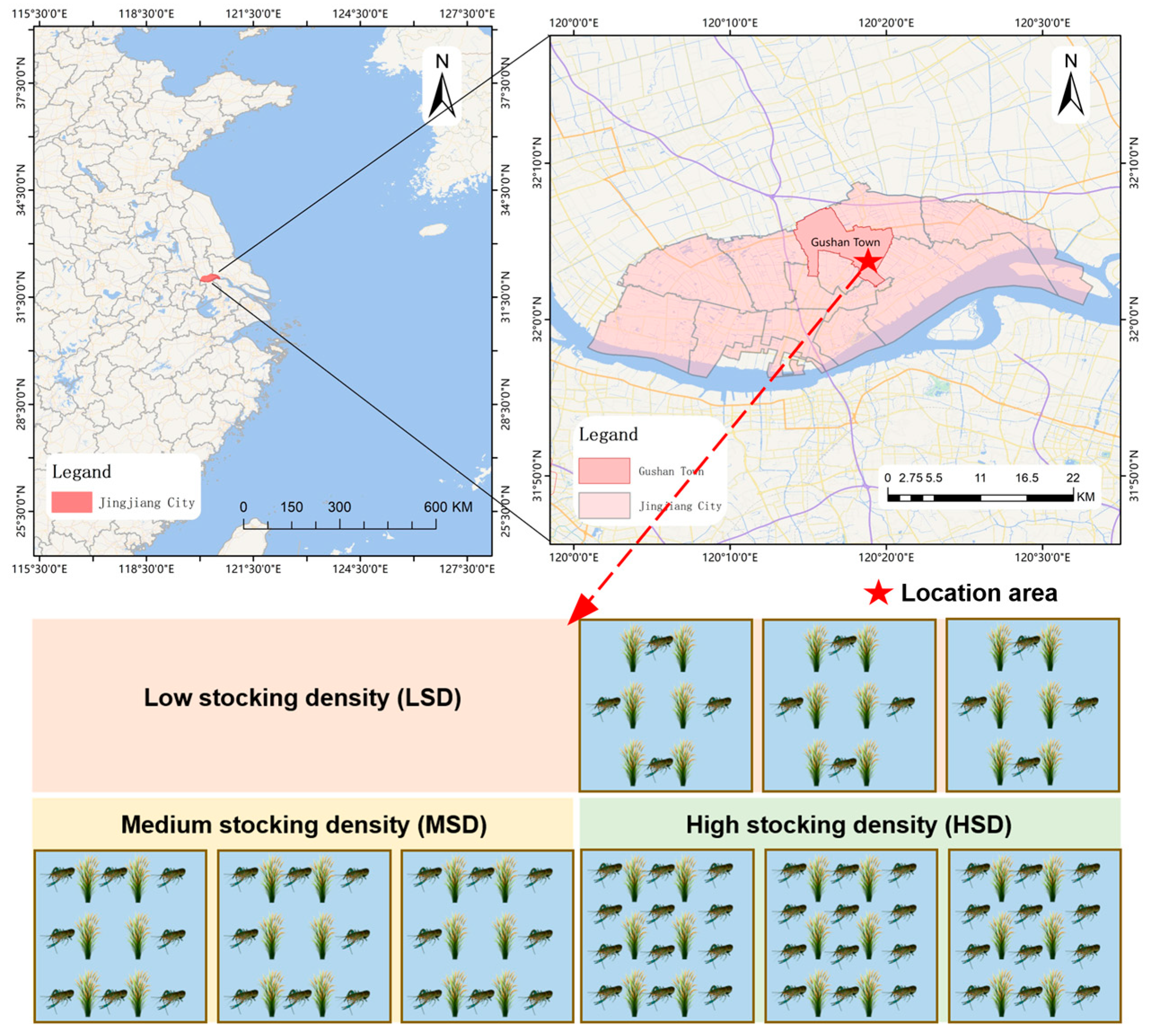
4.1. Experiment Design and Sample Collection
4.2. Environmental Factor Measurements
4.3. Bacterial DNA Extraction, 16S rRNA Sequencing, and Data Processing
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Verdegem, M.; Buschmann, A.H.; Latt, U.W.; Dalsgaard, A.J.T.; Lovatelli, A. The contribution of aquaculture systems to global aquaculture production. J. World Aquac. Soc. 2023, 54, 206–250. [Google Scholar] [CrossRef]
- Yi, X.; Yuan, J.; Zhu, Y.; Yi, X.; Zhao, Q.; Fang, K.; Cao, L. Comparison of the Abundance and Community Structure of N-Cycling Bacteria in Paddy Rhizosphere Soil under Different Rice Cultivation Patterns. Int. J. Mol. Sci. 2018, 19, 3772. [Google Scholar] [CrossRef] [PubMed]
- Boyd, C.E.; McNevin, A.A.; Davis, R.P. The contribution of fisheries and aquaculture to the global protein supply. Food Secur. 2022, 14, 805–827. [Google Scholar] [CrossRef] [PubMed]
- National Bureau of Statistics of China. Statistical Bulletin of the People’s Republic of China on National Economic and Social Development, 2022; National Bureau of Statistics of China: Beijing, China, 2023. [Google Scholar]
- Yu, X.; Hao, X.; Dang, Z.; Yang, L. Industrial development report on integrated rice-fish farming in China (2023). China Fish. 2023, 8, 19–26. [Google Scholar] [CrossRef]
- Ahmed, N.; Hornbuckle, J.; Turchini, G.M. Blue–green water utilization in rice–fish cultivation towards sustainable food production. Ambio 2022, 51, 1933–1948. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Phonexay, M.; Zhang, Z.; Li, C.; Li, J.; Zhang, W. Status of rice-fish farming and rice field fisheries in Northern Laos. Front. Sustain. Food Syst. 2023, 7, 1174172. [Google Scholar] [CrossRef]
- Zhang, Y.; Guan, C.; Li, Z.; Luo, J.; Ren, B.; Chen, C.; Xu, Y.; Ding, J.; Huang, H. Review of Rice-Fish-Duck Symbiosis System in China-One of the Globally Important Ingenious Agricultural Heritage Systems (GIAHS). Sustainability 2023, 15, 1910. [Google Scholar] [CrossRef]
- Zhao, Z.; Chu, C.; Zhou, D.; Wang, Q.; Wu, S.; Zheng, X.; Song, K.; Lv, W. Soil bacterial community composition in rice-fish integrated farming systems with different planting years. Sci. Rep. 2021, 11, 10855. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Ren, W.; Tang, J.; Li, N.; Zhang, J.; Chen, X. The productivity of traditional rice-fish co-culture can be increased without increasing nitrogen loss to the environment. Agric. Ecosyst. Environ. 2013, 177, 28–34. [Google Scholar] [CrossRef]
- Chen, X.; Fan, L.; Qiu, L.; Dong, X.; Wang, Q.; Hu, G.; Meng, S.; Li, D.; Chen, J. Metagenomics Analysis Reveals Compositional and Functional Differences in the Gut Microbiota of Red Swamp Crayfish, Procambarus clarkii, Grown on Two Different Culture Environments. Front. Microbiol. 2021, 12, 735190. [Google Scholar] [CrossRef]
- Fan, L.; Li, F.; Chen, X.; Shen, L.; Chu, Y.; Qiu, L.; Hu, G.; Song, C.; Li, D.; Meng, S.; et al. Co-culture of red swamp crayfish Procambarus clarkia influenced glycoside hydrolase families and fungal communities in the rice-paddy soils. Appl. Soil Ecol. 2023, 186, 104816. [Google Scholar] [CrossRef]
- He, J.; Zhang, X.; Jiang, Z.; Jiang, Y.; Luo, S. Effects of integrated rice-red claw crayfish Cherax quadricarinatus on water environment. Jiangsu Agric. Sci. 2019, 47, 213–215. [Google Scholar]
- Yao, Q. Effects of Different Stocking Densities on the Growth, Digestibility, and Antioxidant Capacity of the Red Claw Crayfish, Cherax quadricarinatus, under Rice-Shrimp Symbiosis Mode; Jilin Agricultural University: Jilin, China, 2023. [Google Scholar]
- Fang, J.; Xu, Y.; Nie, Z.; Xu, G.; Jiang, Z.; Shao, N.; Xiao, Y.; Fang, J. Food sources of common carp in a Hani Terrace integrated rice-fish system (Yunnan Province, China). Aquac. Rep. 2022, 22, 100937. [Google Scholar] [CrossRef]
- Miao, W.; Liu, J.-M.; Zhang, J.; Tang, L.; Zhang, W.-Z.; Xu, Z.-J. Effects of rice-crab coculture on field ecological environment and rice growth: A review. Shengtaixue Zazhi 2020, 39, 2785–2791. [Google Scholar] [CrossRef]
- Naranjo-Páramo, J.; Hernandez-Llamas, A.; Villarreal, H. Effect of stocking density on growth, survival and yield of juvenile redclaw crayfish Cherax quadricarinatus (Decapoda: Parastacidae) in gravel-lined commercial nursery ponds. Aquaculture 2004, 242, 197–206. [Google Scholar] [CrossRef]
- Naranjo-Páramo, J.; Hernández-Llamas, A.; Vargas-Mendieta, M.; Villarreal-Colmenares, H. Stochastic dynamic model analysis of the effect of stocking density on the monosex production of male redclaw crayfish Cherax quadricarinatus reared in commercial gravel-lined ponds. Aquaculture 2021, 535, 736351. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, Z.; Lin, W.; Huang, X.; Zhang, C.; Li, M.; Su, J.; Jiang, Y.; Li, X.; Liu, W.; et al. Rice and red claw crayfish Cherax quadricarinatus cultivation experiment in the integrtaed rice-crayfish farming system. J. Aquac. 2022, 43, 42–44. [Google Scholar]
- Angeler, D.G.; Sánchez-Carrillo, S.; García, G.; Alvarez-Cobelas, M. The influence of Procambarus clarkii (Cambaridae, Decapoda) on water quality and sediment characteristics in a Spanish floodplain wetland. Hydrobiologia 2001, 464, 89–98. [Google Scholar] [CrossRef]
- Palmas, F.; Podda, C.; Frau, G.; Cau, A.; Moccia, D.; Peddio, S.; Solari, P.; Pusceddu, A.; Sabatini, A. Invasive crayfish (Procambarus clarkii, Girard, 1852) in a managed brackish wetland (Sardinia, Italy): Controlling factors and effects on sedimentary organic matter. Estuar. Coast. Shelf Sci. 2019, 231, 106459. [Google Scholar] [CrossRef]
- Hou, Y.; Jia, R.; Ji, P.; Li, B.; Zhu, J. Organic matter degradation and bacterial communities in surface sediment influenced by Procambarus clarkia. Front. Microbiol. 2022, 13, 985555. [Google Scholar] [CrossRef]
- Yamamoto, Y. Contribution of bioturbation by the red swamp crayfishProcambarus clarkii to the recruitment of bloom-forming cyanobacteria from sediment. J. Limnol. 2010, 69, 102–111. [Google Scholar] [CrossRef]
- Ilhéu, M.; Bernardo, J. Aspects of trophic ecology of red swamp crayfish (Procambarus clarkii Girard) in Alentejo, South of Portugal. Actas VI Congr. Esp. Limnol. 1993, 417–423. [Google Scholar] [CrossRef]
- Momot, W. Redefining the Role of Crayfish in Aquatic Ecosystems. Rev. Fish. Sci. 1995, 3, 33–63. [Google Scholar] [CrossRef]
- Day, J.; Diener, C.; Otwell, A.; Tams, K.; Bebout, B.; Detweiler, A.; Lee, M.; Scott, M.; Ta, W.; Ha, M.; et al. Lettuce (Lactuca sativa) productivity influenced by microbial inocula under nitrogen-limited conditions in aquaponics. PLoS ONE 2021, 16, e0247534. [Google Scholar] [CrossRef]
- Herlambang, A.; Murwantoko, M.; Istiqomah, I. Dynamic change in bacterial communities in the integrated rice–fish farming system in Sleman, Yogyakarta, Indonesia. Aquac. Res. 2021, 52, 15432. [Google Scholar] [CrossRef]
- Moriarty, D. The role of microorganisms in aquaculture ponds. Aquaculture 1997, 151, 333–349. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, Q.; Li, Z.; Cheng, W.; Sun, J.; Guo, Z.; Li, Y.; Zhou, J.; Meng, D.; Li, H.; et al. Environmental factors shaping the diversity of bacterial communities that promote rice production. BMC Microbiol. 2018, 18, 51. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, A.C.; Smith, K.L. Soil microbial diversity and the sustainability of agricultural soils. Plant Soil 1995, 170, 75–86. [Google Scholar] [CrossRef]
- Kennedy, A.C. Bacterial diversity in agroecosystems. In Invertebrate Biodiversity as Bioindicators of Sustainable Landscapes; Paoletti, M.G., Ed.; Elsevier: Amsterdam, The Netherlands, 1999; pp. 65–76. [Google Scholar]
- Parkinson, D.; Coleman, D.C. Microbial communities, activity and biomass. Agric. Ecosyst. Environ. 1991, 34, 3–33. [Google Scholar] [CrossRef]
- Smith, R.G.; Gross, K.L.; Robertson, G.P. Effects of Crop Diversity on Agroecosystem Function: Crop Yield Response. Ecosystems 2008, 11, 355–366. [Google Scholar] [CrossRef]
- Zhang, J.; Tang, H.; Zhu, J.; Lin, X.; Feng, Y. Effects of elevated ground-level ozone on paddy soil bacterial community and assembly mechanisms across four years. Sci. Total Environ. 2019, 654, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Jiang, J.; Pan, Y.; Dong, Y.; Chen, Z.; Zhang, G.; Gao, S.; Sun, H.; Guan, X.; Wang, B.; et al. Temporal dynamics of bacterial communities in the water and sediments of sea cucumber (Apostichopus japonicus) culture ponds. Aquaculture 2020, 528, 735498. [Google Scholar] [CrossRef]
- Prijambada, I.D.; Sitompul, R.A.; Widada, J.; Widianto, D. Impact of Agricultural Intensification Practices on Bacterial Community in Agro-ecosystems of Southern Sumatra, Indonesia. Int. J. Agric. Biol. 2012, 14, 816–820. [Google Scholar]
- Singh, U.; Choudhary, A.K.; Sharma, S. Agricultural practices modulate the bacterial communities, and nitrogen cycling bacterial guild in rhizosphere: Field experiment with soybean. J. Sci. Food Agric. 2021, 101, 2687–2695. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, Z.; Cai, H.; Lu, W.; Li, J. Long-term agricultural contamination shaped diversity response of sediment microbiome. J. Environ. Sci. 2021, 99, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Deng, Q.; Wan, L.; Cao, X.; Zhou, Y.; Song, C. Bacterial Communities and Enzymatic Activities in Sediments of Long-Term Fish and Crab Aquaculture Ponds. Microorganisms 2021, 9, 501. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C. Source-sink balance and carbon allocation below ground in plants exposed to ozone: Tansley review. New Phytol. 2003, 157, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Vellend, M. Conceptual Synthesis in Community Ecology. Q. Rev. Biol. 2010, 85, 183–206. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Li, B.; Xu, G.; Li, D.; Zhang, C.; Jia, R.; Li, Q.; Zhu, J. Dynamic and Assembly of Benthic Bacterial Community in an Industrial-Scale In-Pond Raceway Recirculating Culture System. Front. Microbiol. 2021, 12, 797817. [Google Scholar] [CrossRef]
- Yan, H.; Li, F.; Liu, G. Diminishing influence of negative relationship between species richness and evenness on the modeling of grassland α-diversity metrics. Front. Ecol. Evol. 2023, 11, 1108739. [Google Scholar] [CrossRef]
- He, B.; Duan, X.; Rong, L.; Zhang, R.; Li, Y.; Lu, H. Land use controls soil bacterial diversity in the dry-hot valley region, Southern China. Arch. Agron. Soil Sci. 2020, 66, 694–705. [Google Scholar] [CrossRef]
- Alvarez, A.; Maria Saez, J.; Davila Costa, J.S.; Leticia Colin, V.; Soledad Fuentes, M.; Antonio Cuozzo, S.; Susana Benimeli, C.; Alejandra Polti, M.; Julia Amoroso, M. Actinobacteria: Current research and perspectives for bioremediation of pesticides and heavy metals. Chemosphere 2017, 166, 41–62. [Google Scholar] [CrossRef] [PubMed]
- Bull, A.T.; Stach, J.E.M.; Ward, A.C.; Goodfellow, M. Marine actinobacteria: Perspectives, challenges, future directions. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2005, 87, 65–79. [Google Scholar] [CrossRef]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Dong, D.; Wang, H.; Mueller, K.; Qin, Y.; Wang, H.; Wu, W. Metagenomic analysis of microbial consortia enriched from compost: New insights into the role of Actinobacteria in lignocellulose decomposition. Biotechnol. Biofuels 2016, 9, 22. [Google Scholar] [CrossRef]
- de Souza, E.M.; Lamb, T.I.; Lamb, T.A.; Silva, A.d.S.; de Carvalho, S.d.F.; Nyland, V.; Barbosa Lopes, M.C.; Grohs, M.; Marconatto, L.; dos Anjos Borges, L.G.; et al. Rhizospheric Soil from Rice Paddy Presents Isolable Bacteria Able to Induce Cold Tolerance in Rice Plants. J. Soil Sci. Plant Nutr. 2021, 21, 1993–2006. [Google Scholar] [CrossRef]
- Kämpfer, P.; Steiof, M.; Becker, P.M.; Dott, W. Characterization of chemoheterotrophic bacteria associated with the in situ bioremediation of a waste-oil contaminated site. Microb. Ecol. 1993, 26, 161–188. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, B.X.; Ren, H.; Zhang, J. Composition and functional diversity of microbial community across a mangrove-inhabited mudflat as revealed by 16S rDNA gene sequences. Sci. Total Environ. 2018, 633, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; He, R.; Wang, W.; Zhao, D.; Zeng, J.; Huang, R.; Duan, M.; Yu, Z. Composition and co-occurrence patterns of Phragmites australis rhizosphere bacterial community. Aquat. Ecol. 2021, 55, 695–710. [Google Scholar] [CrossRef]
- Yao, M.; Rui, J.; Li, J.; Dai, Y.; Bai, Y.; Hedenec, P.; Wang, J.; Zhang, S.; Pei, K.; Liu, C.; et al. Rate-specific responses of prokaryotic diversity and structure to nitrogen deposition in the Leymus chinensis steppe. Soil Biol. Biochem. 2014, 79, 81–90. [Google Scholar] [CrossRef]
- Louca, S.; Polz, M.F.; Mazel, F.; Albright, M.B.N.; Huber, J.A.; O’Connor, M.I.; Ackermann, M.; Hahn, A.S.; Srivastava, D.S.; Crowe, S.A.; et al. Function and functional redundancy in microbial systems. Nat. Ecol. Evol. 2018, 2, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Jia, R.; Sun, W.; Ding, H.; Li, B.; Zhu, J. Red Claw Crayfish Cherax quadricarinatus Cultivation Influences the Dynamics and Assembly of Benthic Bacterial Communities in Paddy Fields. Environments 2023, 10, 178. [Google Scholar] [CrossRef]
- Hunt, D.E.; Ward, C.S. A network-based approach to disturbance transmission through microbial interactions. Front. Microbiol. 2015, 6, 153683. [Google Scholar] [CrossRef]
- Shaw, G.T.-W.; Liu, A.-C.; Weng, C.-Y.; Chen, Y.-C.; Chen, C.-Y.; Weng, F.C.-H.; Wang, D.; Chou, C.-Y. A network-based approach to deciphering a dynamic microbiome’s response to a subtle perturbation. Sci. Rep. 2020, 10, 19530. [Google Scholar] [CrossRef]
- Li, P.; Wu, G.; Li, Y.; Hu, C.; Ge, L.; Zheng, X.; Zhang, J.; Chen, J.; Zhang, H.; Bai, N.; et al. Long-term rice-crayfish-turtle co-culture maintains high crop yields by improving soil health and increasing soil microbial community stability. Geoderma 2022, 413, 115745. [Google Scholar] [CrossRef]
- Nicholaus, R.; Lukwambe, B.; Zhao, L.; Yang, W.; Zhu, J.; Zheng, Z. Bioturbation of blood clam Tegillarca granosa on benthic nutrient fluxes and microbial community in an aquaculture wastewater treatment system. Int. Biodeterior. Biodegrad. 2019, 142, 73–82. [Google Scholar] [CrossRef]
- Nicholaus, R.; Zheng, Z. The effects of bioturbation by the Venus clam Cyclina sinensis on the fluxes of nutrients across the sediment–water interface in aquaculture ponds. Aquac. Int. 2014, 22, 913–924. [Google Scholar] [CrossRef]
- Qin, Y.; Hou, J.; Deng, M.; Liu, Q.; Wu, C.; Ji, Y.; He, X. Bacterial abundance and diversity in pond water supplied with different feeds. Sci. Rep. 2016, 6, 35232. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Tang, J.; Zhang, C.; Qin, J.; Wang, Y. Bacterial composition, abundance and diversity in fish polyculture and mussel–fish integrated cultured ponds in China. Aquac. Res. 2016, 48, 13221. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, Y.; Tang, J.; Dai, Y. Bacterial communities in Chinese grass carp (Ctenopharyngodon idellus) farming ponds. Aquac. Res. 2013, 45, 138–149. [Google Scholar] [CrossRef]
- Lin, Q.; Li, L.; Adams, J.M.; Hedenec, P.; Tu, B.; Li, C.; Li, T.; Li, X. Nutrient resource availability mediates niche differentiation and temporal co-occurrence of soil bacterial communities. Appl. Soil Ecol. 2021, 163, 103965. [Google Scholar] [CrossRef]
- Kolukirik, M.; Ince, O.; Cetecioglu, Z.; Celikkol, S.; Ince, B.K. Spatial and temporal changes in microbial diversity of the Marmara Sea Sediments. Mar. Pollut. Bull. 2011, 62, 2384–2394. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.N.; Li, X. Effects of straw returnning and biochar on soil bacterial community diversity and co-occurrence network in paddy field. Appl. Ecol. Environ. Res. 2023, 21, 5675–5691. [Google Scholar] [CrossRef]
- Li, Y.M.; Yang, Y.; Wu, T.E.; Zhang, H.; Wei, G.H.; Li, Z.F. Rhizosphere bacterial and fungal spatial distribution and network pattern of Astragalus mongholicus in representative planting sites differ the bulk soil. Appl. Soil Ecol. 2021, 168, 104114. [Google Scholar] [CrossRef]
- Niu, G.X.; Hasi, M.; Wang, R.Z.; Wang, Y.L.; Geng, Q.Q.; Hu, S.Y.; Xu, X.H.; Yang, J.J.; Wang, C.H.; Han, X.G.; et al. Soil microbial community responses to long-term nitrogen addition at different soil depths in a typical steppe. Appl. Soil Ecol. 2021, 167, 104054. [Google Scholar] [CrossRef]
- Wang, M.Y.; Pu, W.M.; Wang, S.Z.; Zeng, X.N.; Sui, X.; Wang, X. pH-Related Changes in Soil Bacterial Communities in the Sanjiang Plain, Northeast China. Microorganisms 2023, 11, 2950. [Google Scholar] [CrossRef]
- Wu, B.H.; Luo, H.Y.; Wang, X.T.; Liu, H.K.; Peng, H.; Sheng, M.P.; Xu, F.; Xu, H. Effects of environmental factors on soil bacterial community structure and diversity in different contaminated districts of Southwest China mine tailings. Sci. Total Environ. 2022, 802, 149899. [Google Scholar] [CrossRef] [PubMed]
- Arunrat, N.; Sansupa, C.; Kongsurakan, P.; Sereenonchai, S.; Hatano, R. Soil Microbial Diversity and Community Composition in Rice-Fish Co-Culture and Rice Monoculture Farming System. Biology 2022, 11, 1242. [Google Scholar] [CrossRef]
- Wei, D.D.; Xing, C.G.; Hou, D.W.; Zeng, S.Z.; Zhou, R.J.; Yu, L.F.; Wang, H.; Deng, Z.X.; Weng, S.P.; He, J.G.; et al. Distinct bacterial communities in the environmental water, sediment and intestine between two crayfish-plant coculture ecosystems. Appl. Microbiol. Biotechnol. 2021, 105, 5087–5101. [Google Scholar] [CrossRef]
- HJ 717-2014; Ministry of Ecology and Environment of China. Soil quality-Determination of Total Nitrogen-Modified Kjeldahl Method. China Environmental Science Press: Beijing, China, 2014.
- HJ 632-2011; Ministry of Ecology and Environment of China. Soil-Determination of Total Phosphorus by Alkali Fusion–Mo-Sb Anti Spectrophotometric Method. China Environmental Science Press: Beijing, China, 2014.
- HJ 634-2012; Ministry of Ecology and Environment of China. Soil-Determination of Ammonium, Nitrite and Nitrate by Extraction with Potassium Chloride Solution-Spectrophotometric Methods. China Environmental Science Press: Beijing, China, 2012.
- Wang, J.; Shi, X.; Zheng, C.; Suter, H.; Huang, Z. Different responses of soil bacterial and fungal communities to nitrogen deposition in a subtropical forest. Sci. Total Environ. 2021, 755, 142449. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef]
- Barberán, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, Y.; Jia, R.; Sun, W.; Li, B.; Zhu, J. Influences of the Integrated Rice-Crayfish Farming System with Different Stocking Densities on the Paddy Soil Microbiomes. Int. J. Mol. Sci. 2024, 25, 3786. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25073786
Hou Y, Jia R, Sun W, Li B, Zhu J. Influences of the Integrated Rice-Crayfish Farming System with Different Stocking Densities on the Paddy Soil Microbiomes. International Journal of Molecular Sciences. 2024; 25(7):3786. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25073786
Chicago/Turabian StyleHou, Yiran, Rui Jia, Wei Sun, Bing Li, and Jian Zhu. 2024. "Influences of the Integrated Rice-Crayfish Farming System with Different Stocking Densities on the Paddy Soil Microbiomes" International Journal of Molecular Sciences 25, no. 7: 3786. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25073786