
Novel Scorpion Toxin ω-Buthitoxin-Hf1a Selectively Inhibits Calcium Influx via CaV3.3 and CaV3.2 and Alleviates Allodynia in a Mouse Model of Acute Postsurgical Pain
 ,
,  , , , , and
, , , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. High-Throughput FLIPR Venom Screening at CaV3.2
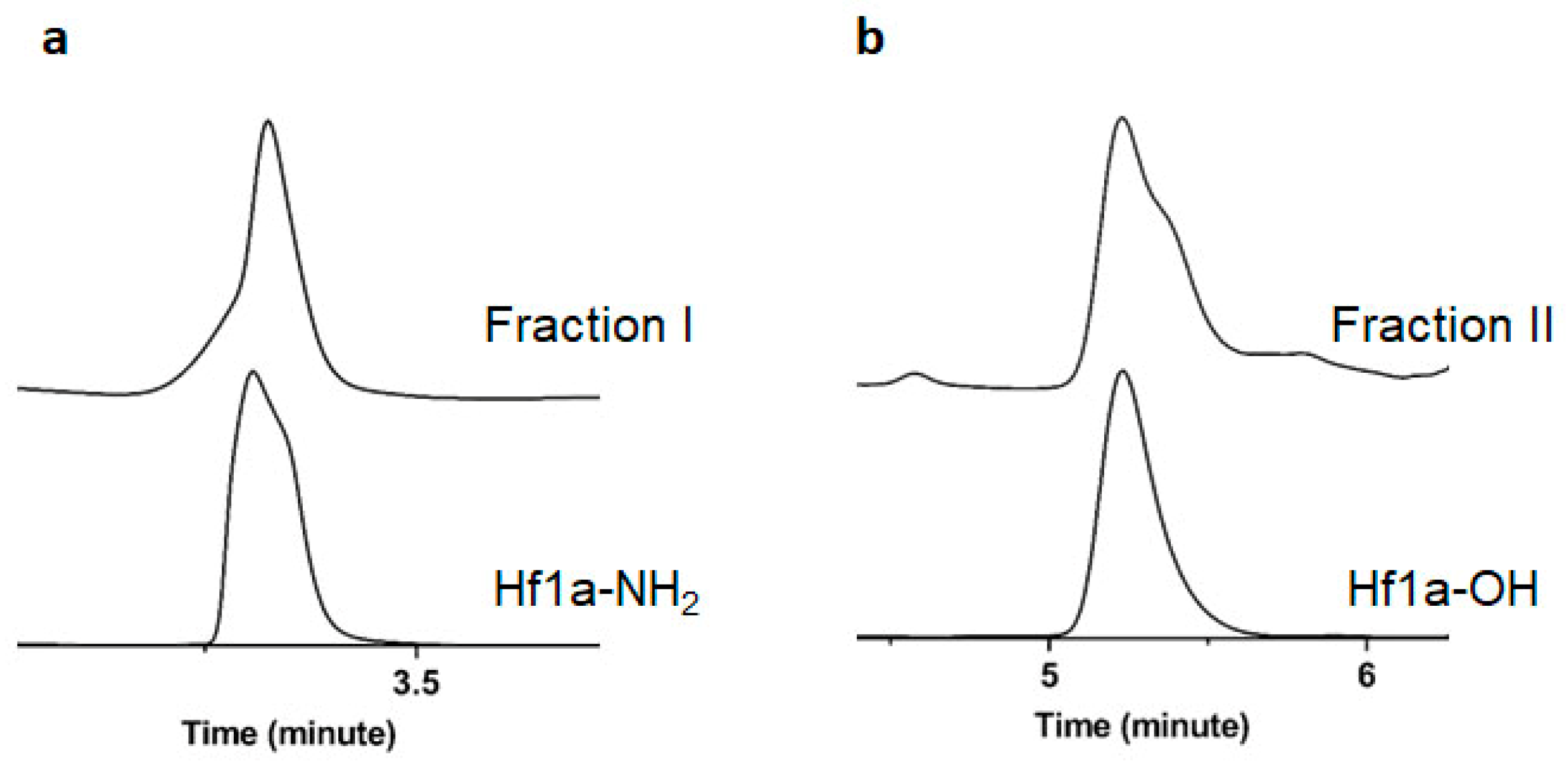
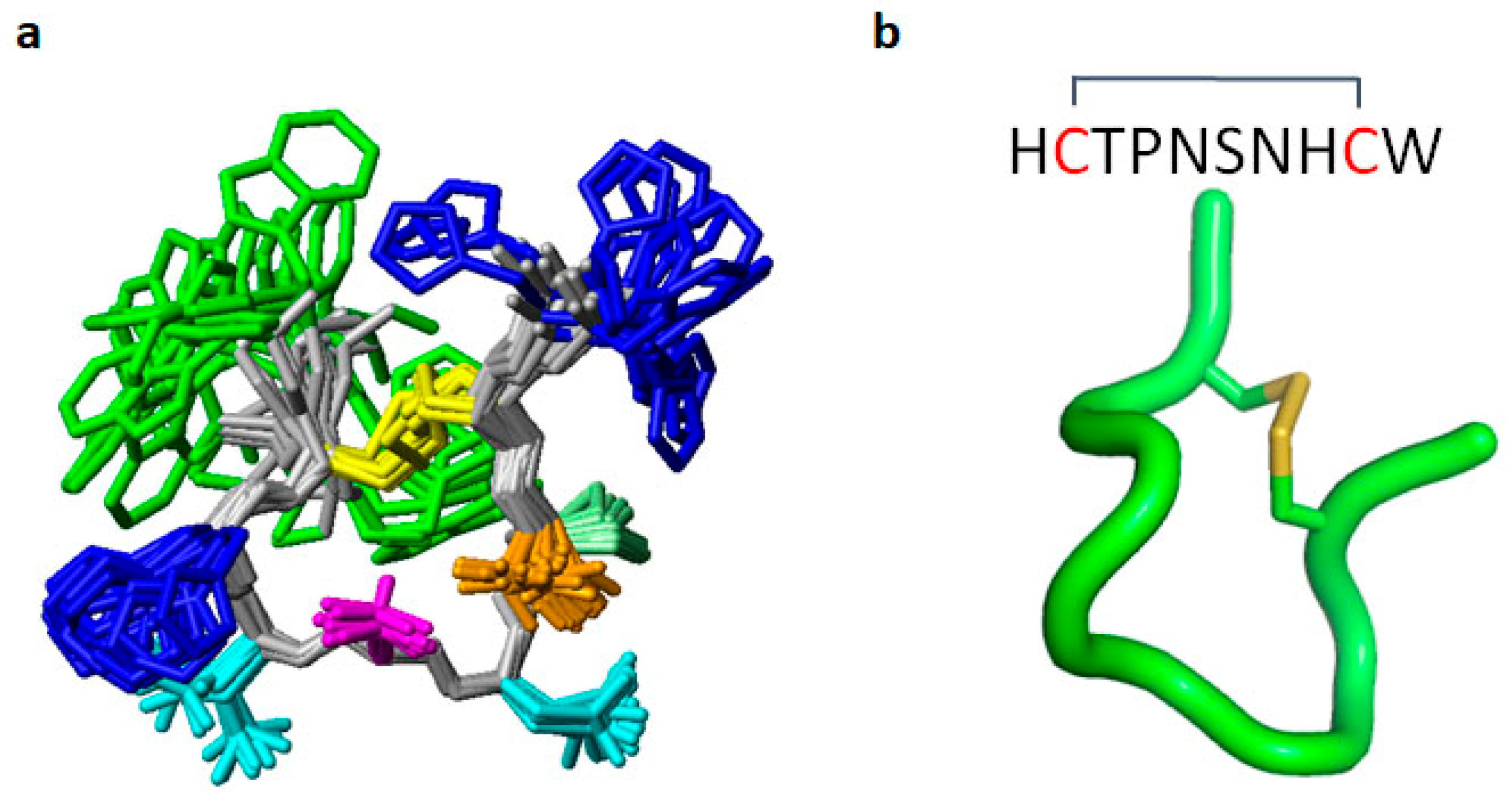
2.2. Determination of Peptide Sequence and NMR Structure
2.3. Evaluation of LVA and HVA CaVs Activities of the Synthetic ω-Buthitoxin-Hf1a
2.3.1. Electrophysiological Characterization of the Scorpion Peptide ω-Buthitoxin-Hf1a in QPatch Assays
2.3.2. Characterization of ω-Buthitoxin-Hf1a in HVA Calcium Channel FLIPR Assays
2.3.3. ω-Buthitoxin-Hf1a Docking in T-Type Calcium Channels
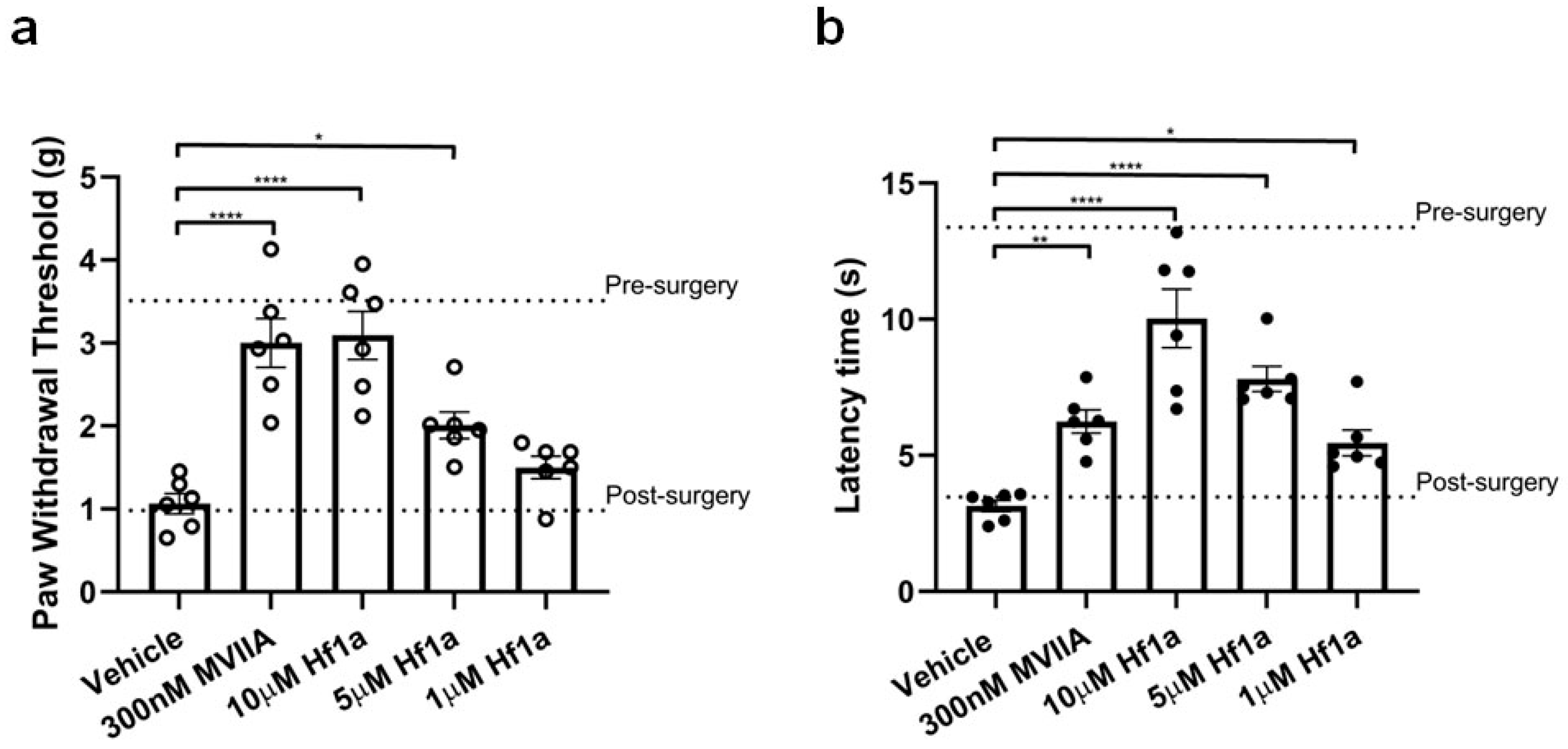
2.4. Local Effects of Hf1a-NH2 on Incision-Induced Mechanical and Thermal Allodynia
2.4.1. A mouse Model of Post-Surgical Pain Based on Plantar Incision Leads to Mechanical and Thermal Allodynia
2.4.2. Hf1a-NH2 Effectively Reduces Mechanical and Thermal Allodynia after Surgery
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. T-Type Calcium Channel Window Current FLIPR Assays
4.3. Crude Venom Collection and Preparation for Screening
4.4. Crude Venom HPLC Fractionation
4.5. Mass Spectrometry Analysis
4.5.1. Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry
4.5.2. Electrospray Ionization Mass Spectrometry
4.6. N-Terminal Sequencing
4.7. Peptide Synthesis
4.8. NMR Spectroscopy
4.9. Structure Calculations
4.10. Whole-Cell Patch-Clamp Electrophysiology
4.11. HVA Calcium Channel FLIPR Assays
4.12. Molecular Docking
4.13. Animals
4.14. Surgery
4.15. Mechanical Paw Withdrawal Threshold Measurements
4.16. Thermal Paw Withdrawal Latency Time Measurements
4.17. Locomotor Performance Assessment
4.18. Treatments
4.19. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rein, J.O. The Scorpion Files. Norwegian University of Science and Technology. 2009. Available online: https://www.ntnu.no/ub/scorpion-files/ (accessed on 1 December 2023).
- Ortiz, E.; Gurrola, G.B.; Schwartz, E.F.; Possani, L.D. Scorpion venom components as potential candidates for drug development. Toxicon 2015, 93, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Petricevich, V.L. Scorpion venom and the inflammatory response. Mediat. Inflamm. 2010, 2010, 903295. [Google Scholar] [CrossRef] [PubMed]
- Possani, L.D.; Merino, E.; Corona, M.; Bolivar, F.; Becerril, B. Peptides and genes coding for scorpion toxins that affect ion-channels. Biochimie 2000, 82, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Tytgat, J.; Chandy, K.G.; Garcia, M.L.; Gutman, G.A.; Martin-Eauclaire, M.-F.; van der Walt, J.J.; Possani, L.D. A unified nomenclature for short-chain peptides isolated from scorpion venoms: α-KTx molecular subfamilies. Trends Pharmacol. Sci. 1999, 20, 444–447. [Google Scholar] [CrossRef] [PubMed]
- Jover, E.; Couraud, F.; Rochat, H. Two types of scorpion neurotoxins characterized by their binding to two separate receptor sites on rat brain synaptosomes. Biochem. Biophys. Res. Commun. 1980, 95, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, K.P.; Barhanin, J.; Lazdunski, M. Specific binding of toxin II from Centruroides suffusus suffusus to the sodium channel in electroplaque membranes. Biochemistry 1982, 21, 5628–5634. [Google Scholar] [CrossRef] [PubMed]
- Cestèle, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Chuang, R.S.; Jaffe, H.; Cribbs, L.; Perez-Reyes, E.; Swartz, K.J. Inhibition of T-type voltage-gated calcium channels by a new scorpion toxin. Nat. Neurosci. 1998, 1, 668–674. [Google Scholar] [CrossRef]
- Sidach, S.S.; Mintz, I.M. Kurtoxin, a gating modifier of neuronal high- and low-threshold ca channels. J. Neurosci. 2002, 22, 2023–2034. [Google Scholar] [CrossRef]
- Talley, E.M.; Cribbs, L.L.; Lee, J.-H.; Daud, A.; Perez-Reyes, E.; Bayliss, D.A. Differential distribution of three members of a gene family encoding low voltage-activated (T-type) calcium channels. J. Neurosci. 1999, 19, 1895–1911. [Google Scholar] [CrossRef]
- Usoskin, D.; Furlan, A.; Islam, S.; Abdo, H.; Lönnerberg, P.; Lou, D.; Hjerling-Leffler, J.; Haeggström, J.; Kharchenko, O.; Kharchenko, P.V. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat. Neurosci. 2015, 18, 145–153. [Google Scholar] [CrossRef]
- Bourinet, E.; Alloui, A.; Monteil, A.; Barrere, C.; Couette, B.; Poirot, O.; Pages, A.; McRory, J.; Snutch, T.P.; Eschalier, A. Silencing of the CaV3.2 T-type calcium channel gene in sensory neurons demonstrates its major role in nociception. EMBO J. 2005, 24, 315–324. [Google Scholar] [CrossRef]
- Choi, S.; Na, H.; Kim, J.; Lee, J.; Lee, S.; Kim, D.; Park, J.; Chen, C.C.; Campbell, K.; Shin, H.S. Attenuated pain responses in mice lacking CaV3.2 T-type channels. Genes. Brain Behav. 2007, 6, 425–431. [Google Scholar] [CrossRef]
- Maeda, Y.; Aoki, Y.; Sekiguchi, F.; Matsunami, M.; Takahashi, T.; Nishikawa, H.; Kawabata, A. Hyperalgesia induced by spinal and peripheral hydrogen sulfide: Evidence for involvement of CaV3.2 T-type calcium channels. Pain 2009, 142, 127–132. [Google Scholar] [CrossRef]
- Shin, H.-S.; Cheong, E.-J.; Choi, S.; Lee, J.; Na, H.S. T-type Ca2+ channels as therapeutic targets in the nervous system. Curr. Opin. Pharmacol. 2008, 8, 33–41. [Google Scholar] [CrossRef]
- Kim, D.; Park, D.; Choi, S.; Lee, S.; Sun, M.; Kim, C.; Shin, H.-S. Thalamic control of visceral nociception mediated by T-type Ca2+ channels. Science 2003, 302, 117–119. [Google Scholar] [CrossRef]
- Todorovic, S.M.; Jevtovic-Todorovic, V. Targeting of CaV3.2 T-type calcium channels in peripheral sensory neurons for the treatment of painful diabetic neuropathy. Pflugers Arch. 2014, 466, 701–706. [Google Scholar] [CrossRef]
- Wen, X.J.; Li, Z.J.; Chen, Z.X.; Fang, Z.Y.; Yang, C.X.; Li, H.; Zeng, Y.M. Intrathecal administration of CaV3.2 and CaV3.3 antisense oligonucleotide reverses tactile allodynia and thermal hyperalgesia in rats following chronic compression of dorsal root of ganglion. Acta Pharmacol. Sin. 2006, 27, 1547–1552. [Google Scholar] [CrossRef]
- Montera, M.; Goins, A.; Cmarko, L.; Weiss, N.; Westlund, K.N.; Alles, S.R. Trigeminal neuropathic pain is alleviated by inhibition of CaV3.3 T-type calcium channels in mice. Channels 2021, 15, 31–37. [Google Scholar] [CrossRef]
- Woolf, C.J. Central sensitization: Implications for the diagnosis and treatment of pain. Pain 2011, 152, S2–S15. [Google Scholar] [CrossRef]
- McGreevy, K.; Bottros, M.M.; Raja, S.N. Preventing chronic pain following acute pain: Risk factors, preventive strategies, and their efficacy. Eur. J. Pain Suppl. 2011, 5, 365–376. [Google Scholar] [CrossRef]
- Gan, T.J.; Habib, A.S.; Miller, T.E.; White, W.; Apfelbaum, J.L. Incidence, patient satisfaction, and perceptions of post-surgical pain: Results from a US national survey. Curr. Med. Res. Opin. 2014, 30, 149–160. [Google Scholar] [CrossRef]
- Kehlet, H.; Jensen, T.S.; Woolf, C.J. Persistent postsurgical pain: Risk factors and prevention. Lancet 2006, 367, 1618–1625. [Google Scholar] [CrossRef]
- Osteen, J.D.; Herzig, V.; Gilchrist, J.; Emrick, J.J.; Zhang, C.; Wang, X.; Castro, J.; Garcia-Caraballo, S.; Grundy, L.; Rychkov, G.Y. Selective spider toxins reveal a role for the NaV1.1 channel in mechanical pain. Nature 2016, 534, 494–499. [Google Scholar] [CrossRef]
- King, G.F.; Vetter, I. No gain, no pain: NaV1.7 as an analgesic target. ACS Chem. Neurosci. 2014, 5, 749–751. [Google Scholar] [CrossRef]
- Ashraf, S. Editorial: Voltage-Gated Calcium 2.2 Channels: Therapeutic Target for Chronic Arthritic Pain? Arthritis Rheumatol. 2015, 67, 1416–1418. [Google Scholar] [CrossRef]
- Altier, C.; Zamponi, G.W. Targeting Ca2+ channels to treat pain: T-type versus N-type. Trends Pharmacol. Sci. 2004, 25, 465–470. [Google Scholar] [CrossRef]
- Snutch, T.P.; Zamponi, G.W. Recent Advances in the Development of T-type Calcium Channel Blockers for Pain Intervention. Br. J. Pharmacol. 2018, 175, 2375–2383. [Google Scholar] [CrossRef]
- Perez-Reyes, E. Molecular physiology of low-voltage-activated t-type calcium channels. Physiol. Rev. 2003, 83, 117–161. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Van Deusen, A.L.; Vitko, I.; Babu, D.A.; Davies, L.A.; Huynh, N.; Cheng, H.; Yang, N.; Barrett, P.Q.; Perez-Reyes, E. Validation of high throughput screening assays against three subtypes of Cav3 T-type channels using molecular and pharmacologic approaches. Assay. Drug Dev. Technol. 2007, 5, 191–204. [Google Scholar] [CrossRef]
- King, G.F.; Gentz, M.C.; Escoubas, P.; Nicholson, G.M. A rational nomenclature for naming peptide toxins from spiders and other venomous animals. Toxicon 2008, 52, 264–276. [Google Scholar] [CrossRef]
- Pineda, S.S.; Chaumeil, P.-A.; Kunert, A.; Kaas, Q.; Thang, M.W.; Le, L.; Nuhn, M.; Herzig, V.; Saez, N.J.; Cristofori-Armstrong, B. ArachnoServer 3.0: An online resource for automated discovery, analysis and annotation of spider toxins. Bioinformatics 2018, 34, 1074–1076. [Google Scholar] [CrossRef]
- He, L.; Yu, Z.; Geng, Z.; Huang, Z.; Zhang, C.; Dong, Y.; Gao, Y.; Wang, Y.; Chen, Q.; Sun, L. Structure, gating, and pharmacology of human CaV3.3 channel. Nat. Commun. 2022, 13, 2084. [Google Scholar] [CrossRef]
- Mueller, A.; Starobova, H.; Morgan, M.; Dekan, Z.; Cheneval, O.; Schroeder, C.I.; Alewood, P.F.; Deuis, J.R.; Vetter, I. Antiallodynic effects of the selective NaV1. 7 inhibitor Pn3a in a mouse model of acute postsurgical pain: Evidence for analgesic synergy with opioids and baclofen. Pain 2019, 160, 1766–1780. [Google Scholar] [CrossRef]
- Hasan, M.M.; Starobova, H.; Mueller, A.; Vetter, I.; Lewis, R.J. Subcutaneous ω-conotoxins alleviate mechanical pain in rodent models of acute peripheral neuropathy. Mar. Drugs 2021, 19, 106. [Google Scholar] [CrossRef]
- Xiao, Y.; Liang, S. Inhibition of neuronal tetrodotoxin-sensitive Na+ channels by two spider toxins: Hainantoxin-III and hainantoxin-IV. Eur. J. Pharmacol. 2003, 477, 1–7. [Google Scholar] [CrossRef]
- Bende, N.S.; Kang, E.; Herzig, V.; Bosmans, F.; Nicholson, G.M.; Mobli, M.; King, G.F. The insecticidal neurotoxin Aps III is an atypical knottin peptide that potently blocks insect voltage-gated sodium channels. Biochem. Pharmacol. 2013, 85, 1542–1554. [Google Scholar] [CrossRef]
- Karbat, I.; Frolow, F.; Froy, O.; Gilles, N.; Cohen, L.; Turkov, M.; Gordon, D.; Gurevitz, M. Molecular basis of the high insecticidal potency of scorpion α-toxins. J. Biol. Chem. 2004, 279, 31679–31686. [Google Scholar] [CrossRef]
- Gurevitz, M.; Karbat, I.; Cohen, L.; Ilan, N.; Kahn, R.; Turkov, M.; Stankiewicz, M.; Stühmer, W.; Dong, K.; Gordon, D. The insecticidal potential of scorpion β-toxins. Toxicon 2007, 49, 473–489. [Google Scholar] [CrossRef] [PubMed]
- Cribbs, L.L.; Lee, J.-H.; Yang, J.; Satin, J.; Zhang, Y.; Daud, A.; Barclay, J.; Williamson, M.P.; Fox, M.; Rees, M. Cloning and characterization of α1H from human heart, a member of the T-type Ca2+ channel gene family. Circ. Res. 1998, 83, 103–109. [Google Scholar] [CrossRef]
- Dubel, S.J.; Altier, C.; Chaumont, S.; Lory, P.; Bourinet, E.; Nargeot, J. Plasma membrane expression of T-type calcium channel α1 subunits is modulated by high voltage-activated auxiliary subunits. J. Biol. Chem. 2004, 279, 29263–29269. [Google Scholar] [CrossRef]
- Wang, D.; Neupane, P.; Ragnarsson, L.; Capon, R.; Lewis, R. Synthesis of Pseudellone Analogs and Characterization as Novel T-type Calcium Channel Blockers. Mar. Drugs 2018, 16, 475. [Google Scholar] [CrossRef]
- Braunschweiler, L.; Ernst, R. Coherence transfer by isotropic mixing: Application to proton correlation spectroscopy. J. Magn. Reson. 1983, 53, 521–528. [Google Scholar] [CrossRef]
- Jeener, J.; Meier, B.; Bachmann, P.; Ernst, R. Investigation of exchange processes by two-dimensional NMR spectroscopy. J. Chem. Phys. 1979, 71, 4546–4553. [Google Scholar] [CrossRef]
- Wüthrich, K. NMR with proteins and nucleic acids. Europhys. News 1986, 17, 11–13. [Google Scholar] [CrossRef]
- Keller, R. The Computer Aided Resonance Assignment Tutorial; CANTINA Verlag: Arth, Switzerland, 2004. [Google Scholar]
- Schroeder, C.I.; Rosengren, K.J. Three-dimensional structure determination of peptides using solution nuclear magnetic resonance spectroscopy. In Snake and Spider Toxins: Methods and Protocols; Humana: New York, NY, USA, 2020; pp. 129–162. [Google Scholar]
- Shen, Y.; Bax, A. Protein backbone and sidechain torsion angles predicted from NMR chemical shifts using artificial neural networks. J. Biomol. NMR 2013, 56, 227–241. [Google Scholar] [CrossRef]
- Cierpicki, T.; Otlewski, J. Amide proton temperature coefficients as hydrogen bond indicators in proteins. J. Biomol. NMR 2001, 21, 249–261. [Google Scholar] [CrossRef]
- Güntert, P. Automated NMR structure calculation with CYANA. In Protein NMR Techniques; Humana: New York, NY, USA, 2004; pp. 353–378. [Google Scholar]
- Brünger, A.T.; Adams, P.D.; Clore, G.M.; DeLano, W.L.; Gros, P.; Grosse-Kunstleve, R.W.; Jiang, J.-S.; Kuszewski, J.; Nilges, M.; Pannu, N.S. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr. D 1998, 54, 905–921. [Google Scholar]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. D 2010, 66, 12–21. [Google Scholar] [CrossRef]
- Koradi, R.; Billeter, M.; Wüthrich, K. MOLMOL: A program for display and analysis of macromolecular structures. J. Mol. Graph. 1996, 14, 51–55. [Google Scholar] [CrossRef]
- Sousa, S.R.; Vetter, I.; Ragnarsson, L.; Lewis, R.J. Expression and pharmacology of endogenous Cav channels in SH-SY5Y human neuroblastoma cells. PLoS ONE 2013, 8, e59293. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, G.; Wu, Q.; Wu, K.; Li, R.; Lei, J.; Pan, X.; Yan, N. Cryo-EM structures of apo and antagonist-bound human Cav3. 1. Nature 2019, 576, 492–497. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Hargreaves, K.; Dubner, R.; Brown, F.; Flores, C.; Joris, J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain 1988, 32, 77–88. [Google Scholar] [CrossRef]
- Leblond, H.; L‘Espérance, M.; Orsal, D.; Rossignol, S. Treadmill locomotion in the intact and spinal mouse. J. Neurosci. 2003, 23, 11411–11419. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Herzig, V.; Dekan, Z.; Rosengren, K.J.; Payne, C.D.; Hasan, M.M.; Zhuang, J.; Bourinet, E.; Ragnarsson, L.; Alewood, P.F.; et al. Novel Scorpion Toxin ω-Buthitoxin-Hf1a Selectively Inhibits Calcium Influx via CaV3.3 and CaV3.2 and Alleviates Allodynia in a Mouse Model of Acute Postsurgical Pain. Int. J. Mol. Sci. 2024, 25, 4745. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094745
Wang D, Herzig V, Dekan Z, Rosengren KJ, Payne CD, Hasan MM, Zhuang J, Bourinet E, Ragnarsson L, Alewood PF, et al. Novel Scorpion Toxin ω-Buthitoxin-Hf1a Selectively Inhibits Calcium Influx via CaV3.3 and CaV3.2 and Alleviates Allodynia in a Mouse Model of Acute Postsurgical Pain. International Journal of Molecular Sciences. 2024; 25(9):4745. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094745
Chicago/Turabian StyleWang, Dan, Volker Herzig, Zoltan Dekan, K. Johan Rosengren, Colton D. Payne, Md. Mahadhi Hasan, Jiajie Zhuang, Emmanuel Bourinet, Lotten Ragnarsson, Paul F. Alewood, and et al. 2024. "Novel Scorpion Toxin ω-Buthitoxin-Hf1a Selectively Inhibits Calcium Influx via CaV3.3 and CaV3.2 and Alleviates Allodynia in a Mouse Model of Acute Postsurgical Pain" International Journal of Molecular Sciences 25, no. 9: 4745. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094745