
Influence of Human Bone Marrow Mesenchymal Stem Cells Secretome from Acute Myeloid Leukemia Patients on the Proliferation and Death of K562 and K562-Lucena Leukemia Cell Lineages

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
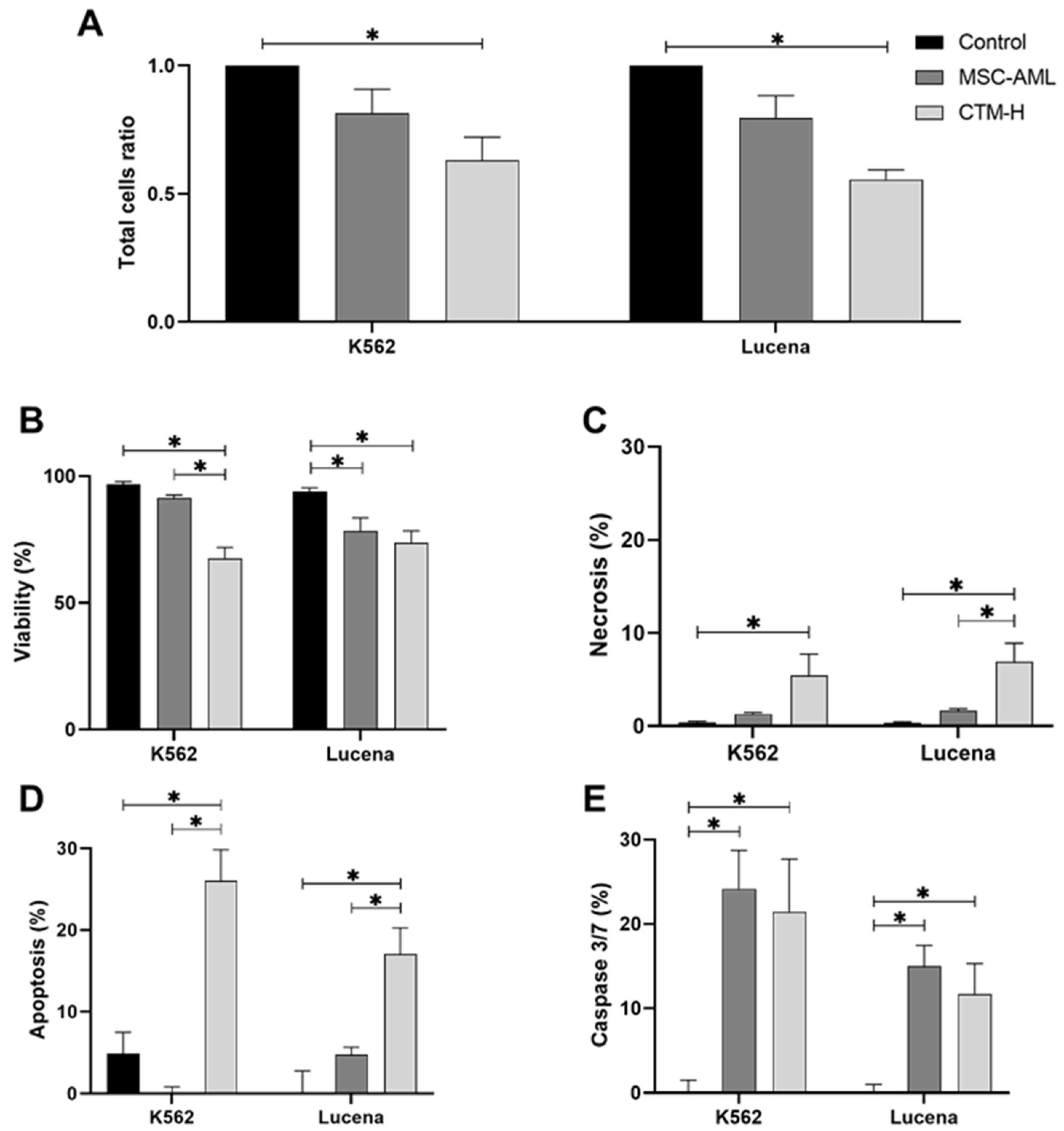
2.1. Cell Death Assays
2.2. Molecular Biology Assays
2.3. Cytokine Array Assay
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transwell Culture
4.1.1. Cell Lineages Culture
4.1.2. Transwell Cultures
4.2. Cell Death Assays
4.2.1. Total Cell Count and Cell Viability Assay
4.2.2. Apoptosis and Necrosis
4.2.3. Detection of Caspase 3/7 Activity
4.2.4. Cell Cycle Analysis
4.3. Molecular Biology Assays
4.3.1. Cyclin D1 (CCND1) and Cyclin D2 (CCND2) Gene Expression
4.3.2. ABC Transporter Gene Expression
4.4. Cytokine Array Assay
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burke, V.P.; Startzell, J.M. The leukemias. Oral Maxillofac. Surg. Clin. N. Am. 2008, 20, 597–608. [Google Scholar] [CrossRef]
- Castelli, G.; Pelosi, E.; Testa, U. Emerging therapies for acute myelogenus leukemia patients targeting apoptosis and mitochondrial metabolism. Cancers 2019, 11, 260. [Google Scholar] [CrossRef]
- Döhner, H.; Weisdorf, D.J.; Bloomfield, C.D. Acute myeloid leukemia. N. Engl. J. Med. 2015, 373, 1136–1152. [Google Scholar] [CrossRef]
- Brunetti, L.; Gundry, M.C.; Goodell, M.A. New insights into the biology of acute myeloid leukemia with mutated npm1. Int. J. Hematol. 2019, 110, 150–160. [Google Scholar] [CrossRef]
- Boulais, P.E.; Frenette, P.S. Making sense of hematopoietic stem cell niches. Blood 2015, 125, 2621–2629. [Google Scholar] [CrossRef]
- Reichert, C.O.; de Freitas, F.A.; Levy, D.; Bydlowski, S.P. Oxysterols and mesenchymal stem cell biology. Vitam. Horm. 2021, 116, 409–436. [Google Scholar]
- Frenette, P.S.; Pinho, S.; Lucas, D.; Scheiermann, C. Mesenchymal stem cell: Keystone of the hematopoietic stem cell niche and a stepping-stone for regenerative medicine. Annu. Rev. Immunol. 2013, 31, 285–316. [Google Scholar] [CrossRef]
- Méndez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; Macarthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef]
- Xia, C.; Wang, T.; Cheng, H.; Dong, Y.; Weng, Q.; Sun, G.; Zhou, P.; Wang, K.; Liu, X.; Geng, Y.; et al. Mesenchymal stem cells suppress leukemia via macrophage-mediated functional restoration of bone marrow microenvironment. Leukemia 2020, 34, 2375–2383. [Google Scholar] [CrossRef]
- Corradi, G.; Baldazzi, C.; Očadlíková, D.; Marconi, G.; Parisi, S.; Testoni, N.; Finelli, C.; Cavo, M.; Curti, A.; Ciciarello, M. Mesenchymal stromal cells from myelodysplastic and acute myeloid leukemia patients display in vitro reduced proliferative potential and similar capacity to support leukemia cell survival. Stem Cell Res. Ther. 2018, 9, 271. [Google Scholar] [CrossRef]
- de Alvarenga, E.C.; Silva, W.N.; Vasconcellos, R.; Paredes-Gamero, E.J.; Mintz, A.; Birbrair, A. Promyelocytic leukemia protein in mesenchymal stem cells is essential for leukemia progression. Ann. Hematol. 2018, 97, 1749–1755. [Google Scholar] [CrossRef]
- Yehudai-Resheff, S.; Attias-Turgeman, S.; Sabbah, R.; Gabay, T.; Musallam, R.; Fridman-Dror, A.; Zuckerman, T. Abnormal morphological and functional nature of bone marrow stromal cells provides preferential support for survival of acute myeloid leukemia cells. Int. J. Cancer 2019, 144, 2279–2289. [Google Scholar] [CrossRef]
- Raaijmakers, M.H.; Mukherjee, S.; Guo, S.; Zhang, S.; Kobayashi, T.; Schoonmaker, J.A.; Ebert, B.L.; Al-Shahrour, F.; Hasserjian, R.P.; Scadden, E.O.; et al. Bone progenitor dysfunction induces myelodysplasia and secondary leukaemia. Nature 2010, 464, 852–857. [Google Scholar] [CrossRef]
- Walkley, C.R.; Olsen, G.H.; Dworkin, S.; Fabb, S.A.; Swann, J.; McArthur, G.A.; Westmoreland, S.V.; Chambon, P.; Scadden, D.T.; Purton, L.E. A microenvironment-induced myeloproliferative syndrome caused by retinoic acid receptor gamma deficiency. Cell 2007, 129, 1097–1110. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, H.; Rodriguez, S.; Cao, L.; Parish, J.; Mumaw, C.; Zollman, A.; Kamoka, M.M.; Mu, J.; Chen, D.Z.; et al. Notch-dependent repression of mir-155 in the bone marrow niche regulates hematopoiesis in an nf-κb-dependent manner. Cell Stem Cell 2014, 15, 51–65. [Google Scholar] [CrossRef]
- Madel, R.J.; Börger, V.; Dittrich, R.; Bremer, M.; Tertel, T.; Phuong, N.N.T.; Baba, H.A.; Kordelas, L.; Staubach, S.; Stein, F.; et al. Independent human mesenchymal stromal cell-derived extracellular vesicle preparations differentially attenuate symptoms in an advanced murine graft-versus-host disease model. Cytotherapy 2023, 25, 821–836. [Google Scholar] [CrossRef]
- Yates, A.G.; Pink, R.C.; Erdbrügger, U.; Siljander, P.R.; Dellar, E.R.; Pantazi, P.; Akbar, N.; Cooke, W.R.; Vatish, M.; Dias-Neto, E.; et al. In sickness and in health: The functional role of extracellular vesicles in physiology and pathology in vivo: Part i: Health and normal physiology: Part i: Health and normal physiology. J. Extracell. Vesicles 2022, 11, e12151. [Google Scholar] [CrossRef]
- Yates, A.G.; Pink, R.C.; Erdbrügger, U.; Siljander, P.R.; Dellar, E.R.; Pantazi, P.; Akbar, N.; Cooke, W.R.; Vatish, M.; Dias-Neto, E.; et al. In sickness and in health: The functional role of extracellular vesicles in physiology and pathology in vivo: Part ii: Pathology: Part ii: Pathology. J. Extracell. Vesicles 2022, 11, e12190. [Google Scholar] [CrossRef]
- Zanetti, S.R.; Romecin, P.A.; Vinyoles, M.; Juan, M.; Fuster, J.L.; Cámos, M.; Querol, S.; Delgado, M.; Menendez, P. Bone marrow msc from pediatric patients with b-all highly immunosuppress t-cell responses but do not compromise cd19-car t-cell activity. J. Immunother. Cancer 2020, 8, e001419. [Google Scholar] [CrossRef]
- Liang, W.; Chen, X.; Zhang, S.; Fang, J.; Chen, M.; Xu, Y.; Chen, X. Mesenchymal stem cells as a double-edged sword in tumor growth: Focusing on msc-derived cytokines. Cell. Mol. Biol. Lett. 2021, 26, 3. [Google Scholar] [CrossRef]
- Baharaghdam, S.; Yousefi, M.; Movasaghpour, A.; Solali, S.; Talebi, M.; Ahani-Nahayati, M.; Lotfimehr, H.; Shamsasanjan, K. Effects of hypoxia on biology of human leukemia t-cell line (molt-4 cells) co-cultured with bone marrow mesenchymal stem cells. Avicenna J. Med. Biotechnol. 2018, 10, 62–68. [Google Scholar]
- Ruiz-Aparicio, P.F.; Uribe, G.I.; Linares-Ballesteros, A.; Vernot, J.P. Sensitization to drug treatment in precursor b-cell acute lymphoblastic leukemia is not achieved by stromal nf-κb inhibition of cell adhesion but by stromal pkc-dependent inhibition of abc transporters activity. Molecules 2021, 26, 5366. [Google Scholar] [CrossRef]
- Paz, J.L.; Levy, D.; Oliveira, B.A.; de Melo, T.C.; de Freitas, F.A.; Reichert, C.O.; Rodrigues, A.; Pereira, J.; Bydlowski, S.P. 7-ketocholesterol promotes oxiapoptophagy in bone marrow mesenchymal stem cell from patients with acute myeloid leukemia. Cells 2019, 8, 482. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Liu, X. Abc family transporters. Adv. Exp. Med. Biol. 2019, 1141, 13–100. [Google Scholar]
- Stutzbach, L. Perk Genetic Variation and Function in Progressive Supranuclear Palsy; University of Pennsylvania Penn Libraries: Philadelphia, PA, USA, 2015. [Google Scholar]
- Maacha, S.; Sidahmed, H.; Jacob, S.; Gentilcore, G.; Calzone, R.; Grivel, J.C.; Cugno, C. Paracrine mechanisms of mesenchymal stromal cells in angiogenesis. Stem Cells Int. 2020, 2020, 4356359. [Google Scholar] [CrossRef]
- Fonseka, M.; Ramasamy, R.; Tan, B.C.; Seow, H.F. Human umbilical cord blood-derived mesenchymal stem cells (hucb-msc) inhibit the proliferation of k562 (human erythromyeloblastoid leukaemic cell line). Cell Biol. Int. 2012, 36, 793–801. [Google Scholar] [CrossRef]
- Huwaikem, M.A.H.; Kalamegam, G.; Alrefaei, G.; Ahmed, F.; Kadam, R.; Qadah, T.; Sait, K.H.W.; Pushparaj, P.N. Human wharton’s jelly stem cell secretions inhibit human leukemic cell line k562 in vitro by inducing cell cycle arrest and apoptosis. Front. Cell Dev. Biol. 2021, 9, 614988. [Google Scholar] [CrossRef]
- Liu, Y.; Song, B.; Wei, Y.; Chen, F.; Chi, Y.; Fan, H.; Liu, N.; Li, Z.; Han, Z.; Ma, F. Exosomes from mesenchymal stromal cells enhance imatinib-induced apoptosis in human leukemia cells via activation of caspase signaling pathway. Cytotherapy 2018, 20, 181–188. [Google Scholar] [CrossRef]
- Tang, H.L.; Tang, H.M.; Mak, K.H.; Hu, S.; Wang, S.S.; Wong, K.M.; Wong, C.S.; Wu, H.Y.; Law, H.T.; Liu, K.; et al. Cell survival, DNA damage, and oncogenic transformation after a transient and reversible apoptotic response. Mol. Biol. Cell 2012, 23, 2240–2252. [Google Scholar] [CrossRef]
- Ding, A.X.; Sun, G.; Argaw, Y.G.; Wong, J.O.; Easwaran, S.; Montell, D.J. Casexpress reveals widespread and diverse patterns of cell survival of caspase-3 activation during development in vivo. eLife 2016, 5, e10936. [Google Scholar] [CrossRef]
- Alves Almeida, P.; Schmitz de Souza, L.F.; Franzoni Maioral, M.; Otto Walter, L.; Fischer Duarte, B.; Mattos Santos-Pirath, Í.; Bauer Speer, D.; Sens, L.; Tizziani, T.; Sena de Oliveira, A.; et al. Cell cycle arrest and apoptosis induction by a new 2,4-dinitrobenzenesulfonamide derivative in acute leukemia cells. J. Pharm. Pharm. Sci. A Publ. Can. Soc. Pharm. Sci. Soc. Can. Des Sci. Pharm. 2021, 24, 23–36. [Google Scholar] [CrossRef]
- Zhu, Y.; Wei, W.; Ye, T.; Liu, Z.; Liu, L.; Luo, Y.; Zhang, L.; Gao, C.; Wang, N.; Yu, L. Small molecule th-39 potentially targets hec1/nek2 interaction and exhibits antitumor efficacy in k562 cells via g0/g1 cell cycle arrest and apoptosis induction. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2016, 40, 297–308. [Google Scholar] [CrossRef]
- Liu, M.; Zhao, X.; Zhao, J.; Xiao, L.; Liu, H.; Wang, C.; Cheng, L.; Wu, N.; Lin, X. Induction of apoptosis, g₀/g₁ phase arrest and microtubule disassembly in k562 leukemia cells by mere15, a novel polypeptide from meretrix meretrix linnaeus. Mar. Drugs 2012, 10, 2596–2607. [Google Scholar] [CrossRef]
- Lee, E.J.; Min, H.Y.; Chung, H.J.; Park, E.J.; Shin, D.H.; Jeong, L.S.; Lee, S.K. A novel adenosine analog, thio-cl-ib-meca, induces g0/g1 cell cycle arrest and apoptosis in human promyelocytic leukemia hl-60 cells. Biochem. Pharmacol. 2005, 70, 918–924. [Google Scholar] [CrossRef]
- Qie, S.; Diehl, J.A. Cyclin d1, cancer progression, and opportunities in cancer treatment. J. Mol. Med. 2016, 94, 1313–1326. [Google Scholar] [CrossRef]
- Poon, R.Y. Cell cycle control: A system of interlinking oscillators. Methods Mol. Biol. 2016, 1342, 3–19. [Google Scholar]
- Wilhide, C.C.; Van Dang, C.; Dipersio, J.; Kenedy, A.A.; Bray, P.F. Overexpression of cyclin d1 in the dami megakaryocytic cell line causes growth arrest. Blood 1995, 86, 294–304. [Google Scholar] [CrossRef]
- Song, J.M.; Xu, D.; Fan, E.J.; Xu, S.R.; Li, D.; Zhao, C.H. cyclin d2 expression in chronic myelogenous leukemia. Zhonghua Xue Ye Xue Za Zhi Zhonghua Xueyexue Zazhi 2004, 25, 103–105. [Google Scholar]
- Chen, B.B.; Glasser, J.R.; Coon, T.A.; Zou, C.; Miller, H.L.; Fenton, M.; McDyer, J.F.; Boyiadzis, M.; Mallampalli, R.K. F-box protein fbxl2 targets cyclin d2 for ubiquitination and degradation to inhibit leukemic cell proliferation. Blood 2012, 119, 3132–3141. [Google Scholar] [CrossRef]
- Luciani, M.F.; Denizot, F.; Savary, S.; Mattei, M.G.; Chimini, G. Cloning of two novel abc transporters mapping on human chromosome 9. Genomics 1994, 21, 150–159. [Google Scholar] [CrossRef]
- Schmitz, G.; Langmann, T. Structure, function and regulation of the abc1 gene product. Curr. Opin. Lipidol. 2001, 12, 129–140. [Google Scholar] [CrossRef]
- Stieger, B.; Meier, Y.; Meier, P.J. The bile salt export pump. Pflug. Arch. Eur. J. Physiol. 2007, 453, 611–620. [Google Scholar] [CrossRef]
- Tian, Y.; Tian, X.; Han, X.; Chen, Y.; Song, C.Y.; Jiang, W.J.; Tian, D.L. Abce1 plays an essential role in lung cancer progression and metastasis. Tumour Biol. J. Int. Soc. Oncodevelopmental Biol. Med. 2016, 37, 8375–8382. [Google Scholar] [CrossRef]
- Hirohashi, T.; Suzuki, H.; Takikawa, H.; Sugiyama, Y. Atp-dependent transport of bile salts by rat multidrug resistance-associated protein 3 (mrp3). J. Biol. Chem. 2000, 275, 2905–2910. [Google Scholar] [CrossRef]
- Balaji, S.A.; Udupa, N.; Chamallamudi, M.R.; Gupta, V.; Rangarajan, A. Role of the drug transporter abcc3 in breast cancer chemoresistance. PLoS ONE 2016, 11, e0155013. [Google Scholar] [CrossRef]
- Lagas, J.S.; Fan, L.; Wagenaar, E.; Vlaming, M.L.; van Tellingen, O.; Beijnen, J.H.; Schinkel, A.H. P-glycoprotein (p-gp/abcb1), abcc2, and abcc3 determine the pharmacokinetics of etoposide. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2010, 16, 130–140. [Google Scholar] [CrossRef]
- Bryan, J.; Muñoz, A.; Zhang, X.; Düfer, M.; Drews, G.; Krippeit-Drews, P.; Aguilar-Bryan, L. Abcc8 and abcc9: Abc transporters that regulate k+ channels. Pflug. Arch. Eur. J. Physiol. 2007, 453, 703–718. [Google Scholar] [CrossRef]
- Kawaguchi, K.; Morita, M. Abc transporter subfamily d: Distinct differences in behavior between abcd1-3 and abcd4 in subcellular localization, function, and human disease. BioMed Res. Int. 2016, 2016, 6786245. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Wang, J.; Yang, J.; Yang, G. Abnormal expression of abcd3 is an independent prognostic factor for colorectal cancer. Oncol. Lett. 2020, 19, 3567–3577. [Google Scholar] [CrossRef]
- Yang, A.; Alrosan, A.Z.; Sharpe, L.J.; Brown, A.J.; Callaghan, R.; Gelissen, I.C. Regulation of abcg4 transporter expression by sterols and lxr ligands. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2021, 1865, 129769. [Google Scholar] [CrossRef] [PubMed]
- Viaud, M.; Abdel-Wahab, O.; Gall, J.; Ivanov, S.; Guinamard, R.; Sore, S.; Merlin, J.; Ayrault, M.; Guilbaud, E.; Jacquel, A.; et al. Abca1 exerts tumor-suppressor function in myeloproliferative neoplasms. Cell Rep. 2020, 30, 3397–3410.e3395. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Luo, Q.; Zeng, S.; Lou, Y.; Li, X.; Hu, M.; Lu, L.; Liu, Z. Disordered farnesoid x receptor signaling is associated with liver carcinogenesis in abcb11-deficient mice. J. Pathol. 2021, 255, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Anglada, M.; Molina-Arcas, M.; Casado, F.J.; Bellosillo, B.; Colomer, D.; Gil, J. Nucleoside transporters in chronic lymphocytic leukaemia. Leukemia 2004, 18, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Kotova, E.; Chen, Z.S.; Lee, K.; Hopper-Borge, E.; Belinsky, M.G.; Kruh, G.D. Mrp8, atp-binding cassette c11 (abcc11), is a cyclic nucleotide efflux pump and a resistance factor for fluoropyrimidines 2′,3′-dideoxycytidine and 9′-(2′-phosphonylmethoxyethyl)adenine. J. Biol. Chem. 2003, 278, 29509–29514. [Google Scholar] [CrossRef]
- Wuxiao, Z.; Wang, H.; Su, Q.; Zhou, H.; Hu, M.; Tao, S.; Xu, L.; Chen, Y.; Hao, X. Microrna-145 promotes the apoptosis of leukemic stem cells and enhances drug-resistant k562/adm cell sensitivity to adriamycin via the regulation of abce1. Int. J. Mol. Med. 2020, 46, 1289–1300. [Google Scholar] [CrossRef]
- Radich, J.P.; Dai, H.; Mao, M.; Oehler, V.; Schelter, J.; Druker, B.; Sawyers, C.; Shah, N.; Stock, W.; Willman, C.L.; et al. Gene expression changes associated with progression and response in chronic myeloid leukemia. Proc. Natl. Acad. Sci. USA 2006, 103, 2794–2799. [Google Scholar] [CrossRef] [PubMed]
- Giannoudis, A.; Davies, A.; Harris, R.J.; Lucas, C.M.; Pirmohamed, M.; Clark, R.E. The clinical significance of abcc3 as an imatinib transporter in chronic myeloid leukaemia. Leukemia 2014, 28, 1360–1363. [Google Scholar] [CrossRef]
- Seborova, K.; Kloudova-Spalenkova, A.; Koucka, K.; Holy, P.; Ehrlichova, M.; Wang, C.; Ojima, I.; Voleska, I.; Daniel, P.; Balusikova, K.; et al. The role of trip6, abcc3 and cps1 expression in resistance of ovarian cancer to taxanes. Int. J. Mol. Sci. 2021, 23, 73. [Google Scholar] [CrossRef]
- De Grouw, E.P.; Raaijmakers, M.H.; Boezeman, J.B.; van der Reijden, B.A.; van de Locht, L.T.; de Witte, T.J.; Jansen, J.H.; Raymakers, R.A. Preferential expression of a high number of atp binding cassette transporters in both normal and leukemic cd34+cd38- cells. Leukemia 2006, 20, 750–754. [Google Scholar] [CrossRef]
- Trojani, A.; Pungolino, E.; Dal Molin, A.; Lodola, M.; Rossi, G.; D’Adda, M.; Perego, A.; Elena, C.; Turrini, M.; Borin, L.; et al. Nilotinib interferes with cell cycle, abc transporters and jak-stat signaling pathway in cd34+/lin- cells of patients with chronic phase chronic myeloid leukemia after 12 months of treatment. PLoS ONE 2019, 14, e0218444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, X.; Wang, X.; Zhou, Y.; Xu, H.; Wang, J.; Huang, L.; Tian, Y.; Cheng, Q. The expression of abcg4, v-atpase and clinic significance of their correlation with nsclc. Zhongguo Fei Ai Za Zhi Chin. J. Lung Cancer 2008, 11, 691–695. [Google Scholar]
- Cole, S.P.; Bhardwaj, G.; Gerlach, J.H.; Mackie, J.E.; Grant, C.E.; Almquist, K.C.; Stewart, A.J.; Kurz, E.U.; Duncan, A.M.; Deeley, R.G. Overexpression of a transporter gene in a multidrug-resistant human lung cancer cell line. Science 1992, 258, 1650–1654. [Google Scholar] [CrossRef] [PubMed]
- Andersson, L.C.; Nilsson, K.; Gahmberg, C.G. K562—A human erythroleukemic cell line. Int. J. Cancer 1979, 23, 143–147. [Google Scholar] [CrossRef]
- Rumjanek, V.M.; Trindade, G.S.; Wagner-Souza, K.; de-Oliveira, M.C.; Marques-Santos, L.F.; Maia, R.C.; Capella, M.A. Multidrug resistance in tumour cells: Characterization of the multidrug resistant cell line k562-lucena 1. An. Da Acad. Bras. De Cienc. 2001, 73, 57–69. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokine | K562 | Lucena | MSC-AML | MSC-H |
|---|---|---|---|---|
| CCL2/MCP-1 | - | - | 24.03 ± 0.44 | 4.41 ± 0.16 |
| CXCL12/SDF-1 | - | - | 4.16 ± 3.09 | 4.15 ± 0.71 |
| IL-6 | - | - | 7.87 ± 4.73 | 20.60 ± 0.54 |
| IL-8 | 4.45 ± 0.09 | 3.30 ± 0.50 | 8.67 ± 0.36 | 3.23 ± 0.21 |
| MIF | - | - | 2.93 ± 1.22 | 2.72 ± 0.45 |
| Serpin E1/PAI-1 | - | - | 39.28 ± 5.27 | 39.11 ± 2.71 |
| Cytokine | K562 | Lucena | ||||
|---|---|---|---|---|---|---|
| MSC-AML | MSC-H | p | MSC-AML | MSC-H | p | |
| CCL2/MCP-1 | 22.8 ± 2.6 | 113.5 ± 11.4 | 0.016 | 17.5 ± 0.7 | 78.5 ± 5.0 | 0.007 |
| CXCL12/SDF-1 | 5.1 ± 1.1 | 8.5 ± 0.1 | 0.082 | 4.4 ± 0.1 | 3.1 ± 0.1 | 0.012 |
| IL-6 | 83.1 ± 2.8 | 65.4 ± 1.1 | 0.027 | 84.5 ± 6.3 | 128.5 ± 1.5 | 0.021 |
| IL-8 | 28.7 ± 2.4 | 15.8 ± 0.1 | 0.033 | 23.9 ± 2.3 | 43.0 ± 2.2 | 0.027 |
| MIF | 27.5 ± 1.8 | 32.8 ± 1.8 | 0.167 | 22.6 ± 1.1 | 20.8 ± 1.1 | 0.369 |
| Serpin E1/PAI-1 | 172.9 ± 5.1 | 188.7 ± 20.9 | 0.540 | 136.8 ± 2.6 | 182.1 ± 2.3 | 0.006 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Freitas, F.A.; Levy, D.; Reichert, C.O.; Sampaio-Silva, J.; Giglio, P.N.; de Pádua Covas Lage, L.A.; Demange, M.K.; Pereira, J.; Bydlowski, S.P. Influence of Human Bone Marrow Mesenchymal Stem Cells Secretome from Acute Myeloid Leukemia Patients on the Proliferation and Death of K562 and K562-Lucena Leukemia Cell Lineages. Int. J. Mol. Sci. 2024, 25, 4748. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094748
de Freitas FA, Levy D, Reichert CO, Sampaio-Silva J, Giglio PN, de Pádua Covas Lage LA, Demange MK, Pereira J, Bydlowski SP. Influence of Human Bone Marrow Mesenchymal Stem Cells Secretome from Acute Myeloid Leukemia Patients on the Proliferation and Death of K562 and K562-Lucena Leukemia Cell Lineages. International Journal of Molecular Sciences. 2024; 25(9):4748. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094748
Chicago/Turabian Stylede Freitas, Fábio Alessandro, Débora Levy, Cadiele Oliana Reichert, Juliana Sampaio-Silva, Pedro Nogueira Giglio, Luís Alberto de Pádua Covas Lage, Marco Kawamura Demange, Juliana Pereira, and Sérgio Paulo Bydlowski. 2024. "Influence of Human Bone Marrow Mesenchymal Stem Cells Secretome from Acute Myeloid Leukemia Patients on the Proliferation and Death of K562 and K562-Lucena Leukemia Cell Lineages" International Journal of Molecular Sciences 25, no. 9: 4748. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms25094748