First Report on the Occurence of Dermatophytes of Microsporum Cookei Clade and Close Affinities to Paraphyton Cookei in the Harmanecká Cave (Veľká Fatra Mts., Slovakia)

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sample Collection
2.3. Isolation of Fungi from Sediment and Soil Samples
2.4. Identification of Fungi
2.5. Bioinformatics Data Analysis
2.6. The Assimilation of Various Carbon Sources
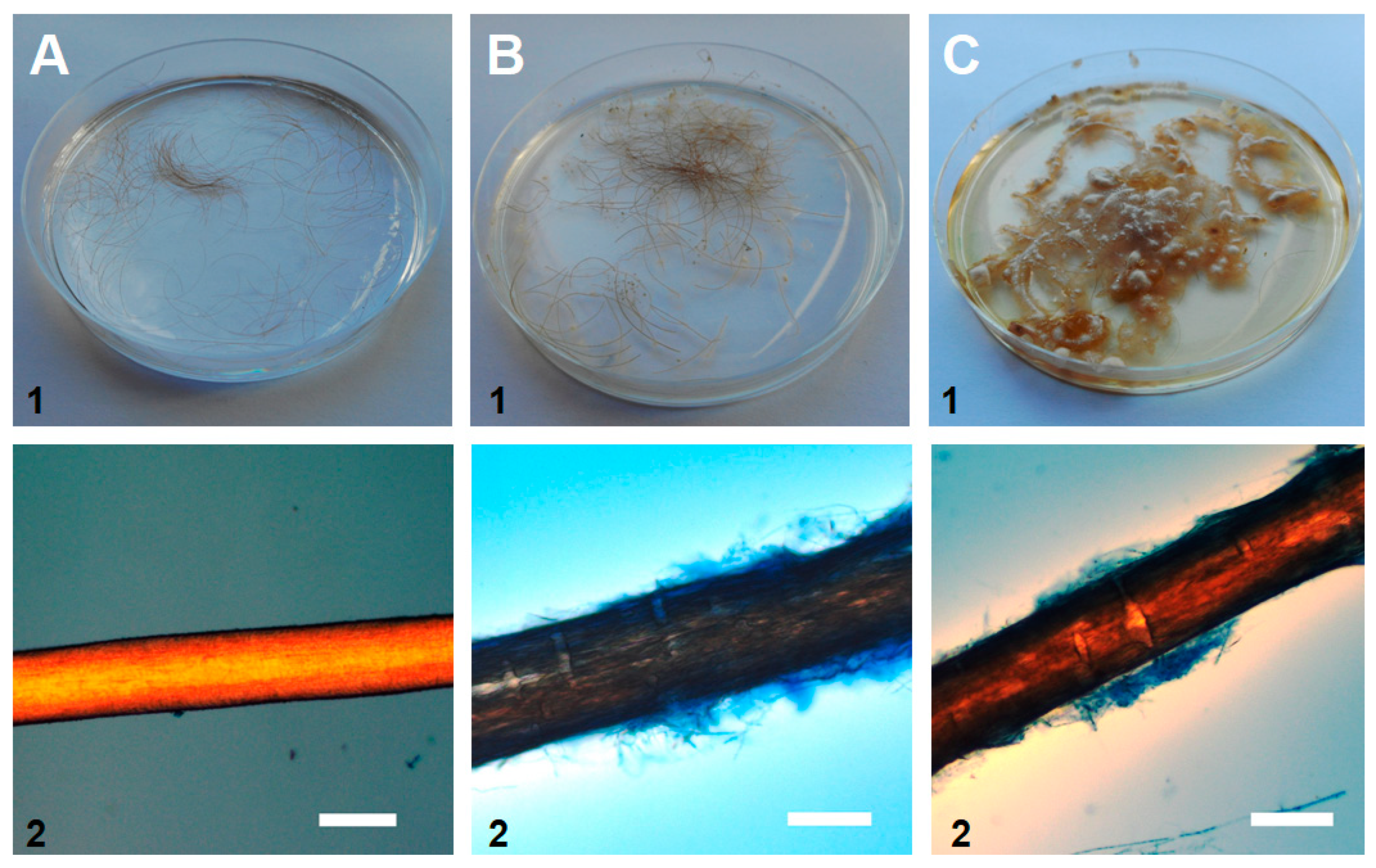
2.7. The in Vitro Hair Perforation Test
2.8. Survival at Different Temperatures
2.9. Growth at Different Temperatures
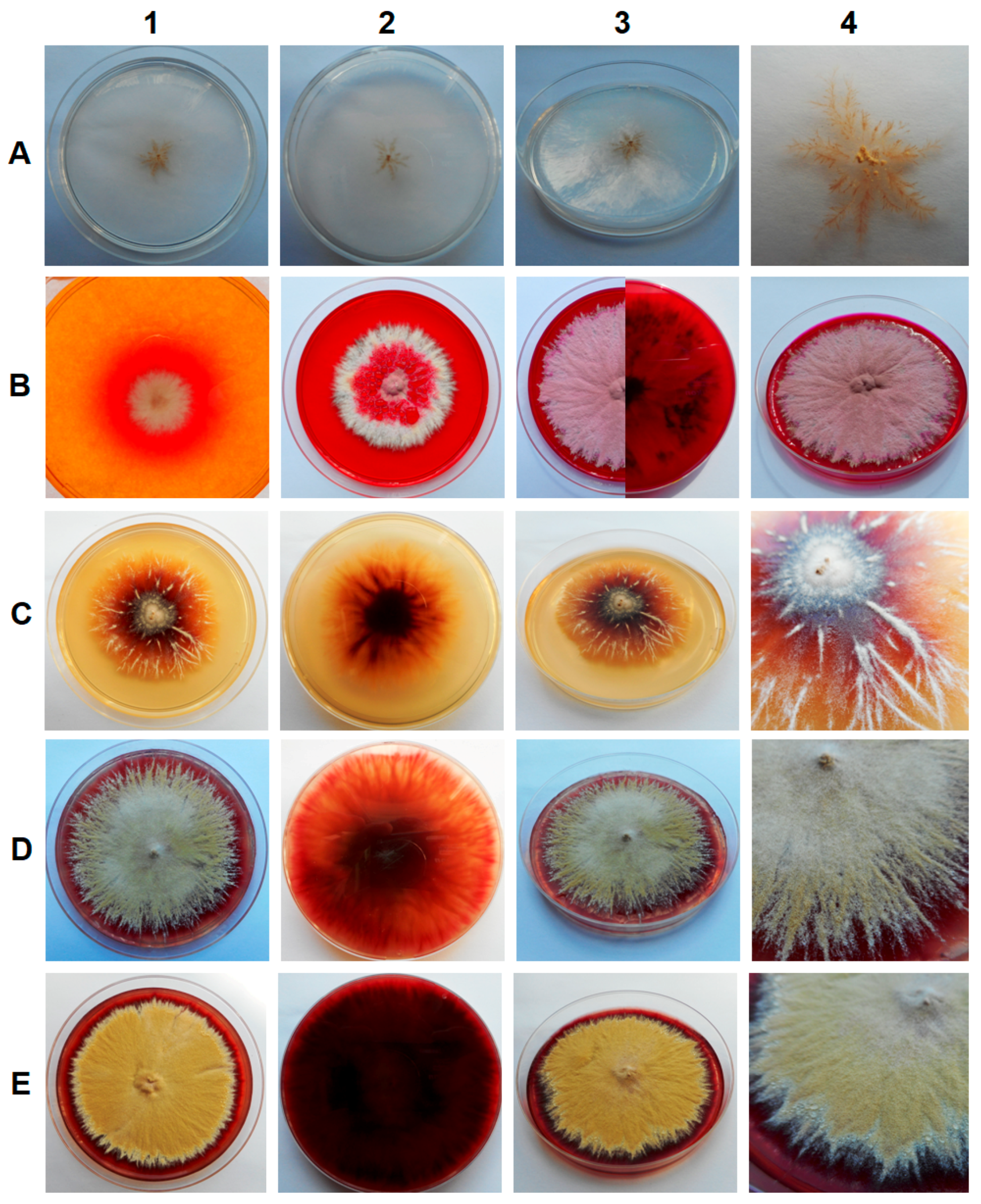
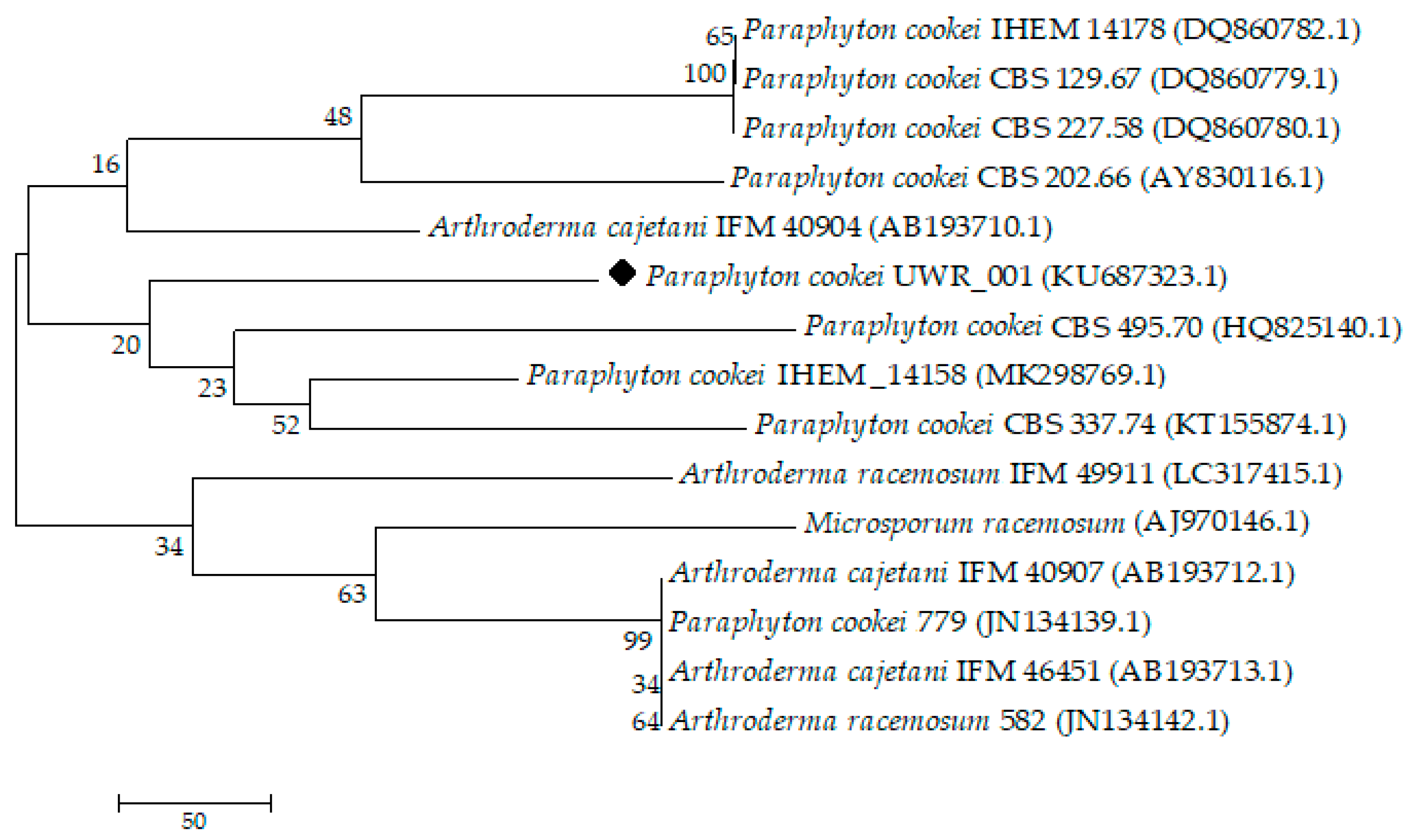
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharma, R.; Rajak, R.C. Keratinophilic fungi: nature’s keratin degrading machines! Resonance 2003, 8, 28–40. [Google Scholar] [CrossRef]
- Gherbawy, Y.A.M.H. Keratinolytic and keratinophilic fungi of mangrove’s soil and air in the city of Qena and their response to garlic extract and onion oil treatments. Acta Mycol. 1996, 31, 87–89. [Google Scholar] [CrossRef]
- Ulfig, K.; Płaza, G.; Terakowskip, M.; Janda-Ulfig, K. Sewage sludge open-air drying affects on keratinolytic, keratinophilic and actidione-resistant fungi. Roczniki PZH 2006, 57, 371–379. [Google Scholar]
- Kunert, J. Physiology of keratinophilic fungi. Rev. Iberoam Micol. 2000, 669, 77–85. [Google Scholar]
- Philpot, C.M. The use of nutritional tests for the differentiation of dermatophytes. Sabouraudia 1977, 15, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Vanderwolf, K.J.; Malloch, D.; McAlpine, D.F.; Forbes, G.J. A world review of fungi, yeasts, and slime molds in caves. Int. J. Speleol. 2013, 42, 77–96. [Google Scholar] [CrossRef]
- Weitzman, I.; Summerbell, R.C. The Dermatophytes. CMR 1995, 8, 240–259. [Google Scholar] [CrossRef]
- Simpanya, M.F. Dermatophytes: Their taxonomy, ecology and pathogenicity. Rev. Iberoam Micol. 2000, 669, 1–12. [Google Scholar]
- de Hoog, G.S.; Dukik, K.; Monod, M.; Packeu, A.; Stubbe, D.; Hendrickx, M.; Kupsch, C.; Stielow, J.B.; Freeke, J.; Göker, M.; et al. Toward a novel multilocus phylogenetic taxonomy for the dermatophytes. Mycopathologia 2017, 182, 5–31. [Google Scholar] [CrossRef]
- Lurie, H.I.; Borok, R. Trichophyton mentagrophytes isolated from the soil of caves. Mycologia 1955, 47, 506–510. [Google Scholar] [CrossRef]
- Lurie, H.I.; Way, M. The isolation of dermatophytes from the atmosphere of caves. Mycologia 1957, 49, 178–180. [Google Scholar] [CrossRef]
- Kajihiro, E.S. Occurrence of dermatophytes in fresh bat guano. Appl. Microbiol. 1965, 13, 720–724. [Google Scholar] [PubMed]
- Balabanoff, V.A. Comparative studies of dermatophytes isolated from caves and stables in Bulgaria. Mycopathol. Mycol. Appl. 1967, 32, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Bella, P.; Hlavac, J.; Gazik, P. Protection and Management of Show Caves in Slovakia. In Proceedings of the 13th International Congress of Speleology 4th Speleological Congress of Latin América and Caribbean 26th Brazilian Congress of Speleology Brasília, Brasília, Brazil, 15–22 July 2001; pp. 287–290. [Google Scholar]
- Bella, P. Slovakia Show Caves; Grafon: Liptovský Mikuláš, Slovakia, 2003; pp. 21–55. [Google Scholar]
- Nudziková, Ľ. Course of show caves attendance in Slovakia since 2009. Aragonit 2014, 19, 35–38. (In Slovak) [Google Scholar]
- Ogórek, R.; Dyląg, M.; Kozak, B. Dark stains on rock surfaces in Driny Cave (Little Carpathian Mountains, Slovakia). Extremophiles 2016, 20, 641–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehotská, B.; Lehotský, R. 15 years of winter monitoring of bats in the Driny Cave. Aragonit 2009, 14, 171–172. (In Slovak) [Google Scholar]
- Vanbreuseghem, R. Technique biologique pour I’ isolement des dermat ophytes dusol. Ann. Soc. Belge. Med. Trop. 1952, 32, 173–178. [Google Scholar]
- Choi, J.S.; Gräser, Y.; Walther, G.; Peano, A.; Symoens, F.; de Hoog, S. Microsporum mirabile and its teleomorph Arthroderma mirabile, a new dermatophyte species in the M. cookei clade. Med. Mycol. 2012, 50, 161–169. [Google Scholar] [CrossRef]
- Metin, B.; Heitman, J. Sexual reproduction in dermatophytes. Mycopathol 2017, 182, 45–55. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytoch. Bull. 1987, 19, 11–15. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Ogórek, R.; Lejman, A.; Matkowski, K. Fungi isolated from the Niedźwiedzia Cave in Kletno (Lower Silesia, Poland). Int. J. Speleol. 2013, 42, 161–166. [Google Scholar] [CrossRef]
- Nováková, A.; Hubka, V.; Valinová, Š.; Kolařík, M.; Hillebrand-Voiculescu, A.M. Cultivable microscopic fungi from an underground chemosynthesis-based ecosystem: A preliminary study. Folia Microbiol. 2018, 63, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Poulson, T.L.; White, W.B. The cave environment. Science 1969, 165, 971–981. [Google Scholar] [CrossRef]
- Nováková, A. Microscopic fungi isolated from the Domica Cave system (Slovak Karst National Park, Slovakia). A review. Int. J. Speleol. 2009, 38, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Pusz, W.; Ogórek, R.; Uklańska-Pusz, C.; Zagożdżon, P. Speleomycological research in underground Osówka Complex in Sowie Mountains (Lower Silesia, Poland). Int. J. Speleol. 2014, 43, 27–34. [Google Scholar] [CrossRef]
- Mulec, J.; Vaupotic, J.; Walochnik, J. Prokaryotic and eukaryotic airborne microorganisms as tracers of microclimatic changes in the underground (Postojna Cave, Slovenia). Environ. Microbiol. 2012, 64, 654–667. [Google Scholar] [CrossRef]
- Wang, W.; Ma, X.; Ma, Y.; Mao, L.; Wu, F.; Ma, X.; An, L.; Feng, H. Seasonal dynamics of airborne fungi in different caves of the Mogao Grottoes, Dunhuang, China. Int. Biodeter. Biodeger. 2010, 64, 461–466. [Google Scholar] [CrossRef]
- Borelli, D. Microsporum racemosum nova species. Acta Med. Venez. 1965, 12, 148–151. [Google Scholar]
- Gianfreda, L.; Rao, M. Potential of extra cellular enzymes in remediation of polluted soils: A review. Enzyme Microb. Technol. 2004, 35, 339–354. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Agrawal, S.C. In vitro degradation of human hair by some keratinophilic fungi. Mykosen 1982, 25, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Morita, R.Y. Psychrophilic bacteria. Bacteriol. Rev. 1975, 39, 144–167. [Google Scholar] [PubMed]
- Al-Fakih, A.A. Overview on the fungal metabolites involved in mycopathy. Open J. Med. Microbiol. 2014, 4, 38–63. [Google Scholar] [CrossRef]
- Schabereiter-Gurtner, C.; Saiz-Jimenez, C.; Piñar, G.; Lubitz, W.; Rolleke, S. Phylogenetic diversity of bacteria associated with Paleolithic paintings and surrounding rock walls in two Spanish caves (Llonin and La Garma). FEMS Microb. Ecol. 2004, 47, 1–13. [Google Scholar] [CrossRef]
- McKenzie, R.L.; Bjorn, L.O.; Bais, A.; Iayis, M. Changes in biologically active ultraviolet radiation reaching the Earth’s Surface. Photochem. Photobiol. Sci. 2003, 2, 5–15. [Google Scholar] [CrossRef]
- Kidd, S.; Halliday, C.; Alexiou, H.; Ellis, D. Descriptions of Medical Fungi, 3rd ed.; Newstyle Printing: Adelaide, South Australia, 2006; p. 149. [Google Scholar]
- Bastian, F.; Jurado, V.; Nováková, A.; Alabouvette, C.; Saiz-Jimenez, C. The microbiology of Lascaux Cave. Microbiology 2010, 156, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Griffin, D.W.; Gray, M.A.; Lyles, M.B.; Northup, D.E. The transport of nonindigenous microorganisms into caves by human visitation: A case study at Carlsbad Caverns National Park. Geomicrob. J. 2014, 31, 175–185. [Google Scholar] [CrossRef]
- Martin-Sanchez, P.M.; Nováková, A.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Two new species of the genus Ochroconis, O. lascauxensis and O. anomala isolated from black stains in Lascaux Cave, France. Fungal Biol. 2012, 116, 574–589. [Google Scholar] [CrossRef] [PubMed]
- Ogórek, R.; Višňovská, Z.; Tančinová, D. Mycobiota of underground habitats: Case study of Harmanecká Cave in Slovakia. Microb. Ecol. 2016, 71, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Kokurewicz, T.; Ogórek, R.; Pusz, W.; Matkowski, K. Bats increase the number of cultivable airborne fungi in the “Nietoperek’’ bat reserve in Western Poland. Microb. Ecol. 2016, 72, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Victor, T.R.; Rajkumar, S.S.; Li, X.; Okoniewski, J.C.; Hicks, A.C.; Davis, A.D.; Broussard, K.; LaDeau, S.L.; Chaturvedi, S.; et al. Mycobiome of the bat White Nose Syndrome affected caves and mines reveals diversity of fungi and local adaptation by the fungal pathogen Pseudogymnoascus (Geomyces) destructans. PLoS ONE 2014, 9, e116149. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth at Different Temperatures | |||||||
| Incubation Period [day] | 5.0 °C | 10.0 °C | 15.0 °C | 20.0 °C | 25.0 °C | 30.0 °C | 37.0 °C |
| 28 | ― 1 | ― | + | ++ | ++++ | ++ | + |
| 56 | + | + | ND | ND | ND | ND | ND |
| Survival at Different Temperatures | |||||||
| Incubation period [day] | 5.0 °C | −10.0 °C | −25.0 °C | −72.0 °C | |||
| 14 | + 2 | + | + | + | |||
| 28 | + | + | + | + | |||
| 42 | + | + | + | + | |||
| 56 | + | + | + | + | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogórek, R.; Piecuch, A.; Višňovská, Z.; Cal, M.; Niedźwiecka, K. First Report on the Occurence of Dermatophytes of Microsporum Cookei Clade and Close Affinities to Paraphyton Cookei in the Harmanecká Cave (Veľká Fatra Mts., Slovakia). Diversity 2019, 11, 191. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100191
Ogórek R, Piecuch A, Višňovská Z, Cal M, Niedźwiecka K. First Report on the Occurence of Dermatophytes of Microsporum Cookei Clade and Close Affinities to Paraphyton Cookei in the Harmanecká Cave (Veľká Fatra Mts., Slovakia). Diversity. 2019; 11(10):191. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100191
Chicago/Turabian StyleOgórek, Rafał, Agata Piecuch, Zuzana Višňovská, Magdalena Cal, and Katarzyna Niedźwiecka. 2019. "First Report on the Occurence of Dermatophytes of Microsporum Cookei Clade and Close Affinities to Paraphyton Cookei in the Harmanecká Cave (Veľká Fatra Mts., Slovakia)" Diversity 11, no. 10: 191. https://0-doi-org.brum.beds.ac.uk/10.3390/d11100191