
Impact of Algicidal Bacillus mycoides on Diatom Ulnaria acus from Lake Baikal

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Diatom Culture
2.2. Isolation of Algicidal Bacteria
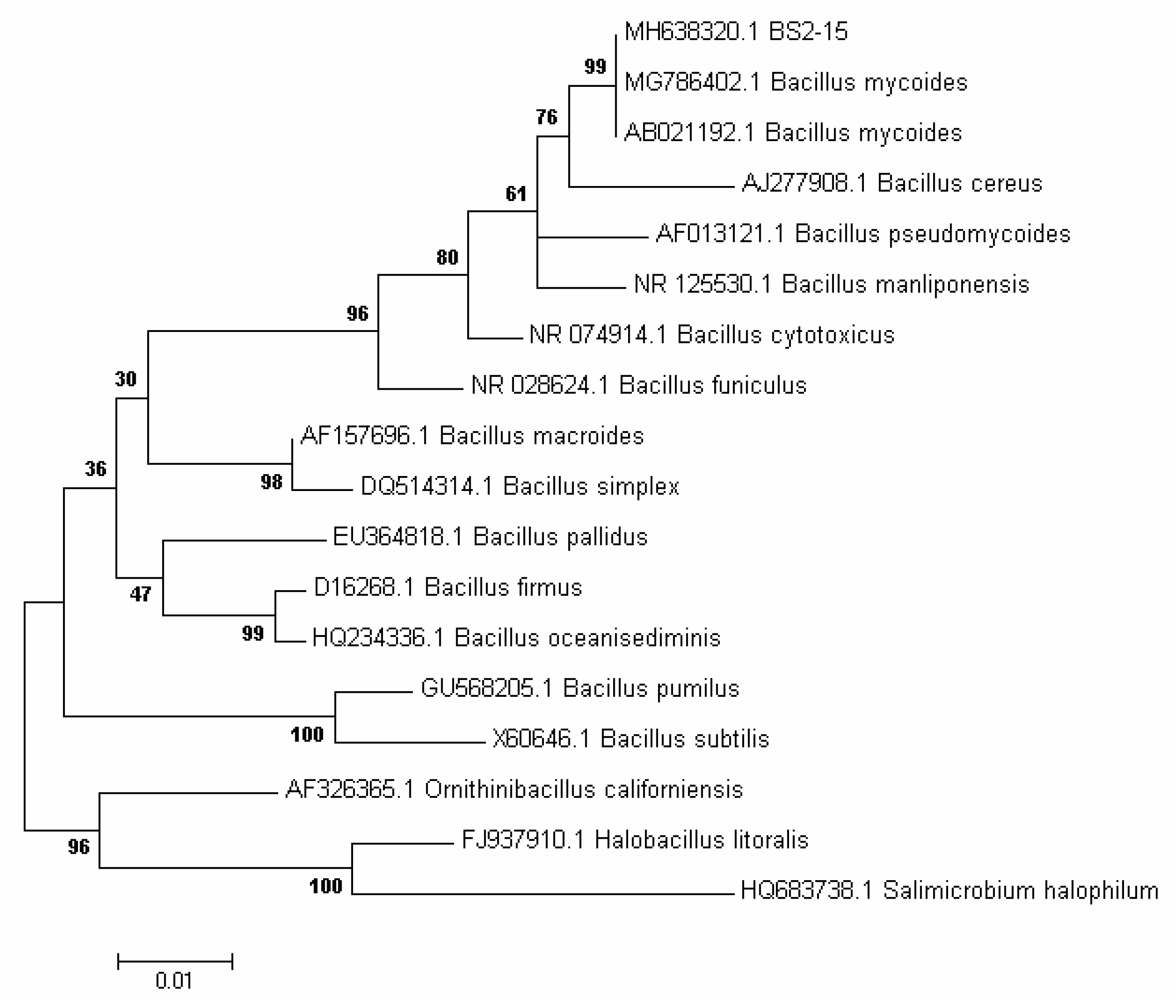
2.3. Identification of Selected Bacterium
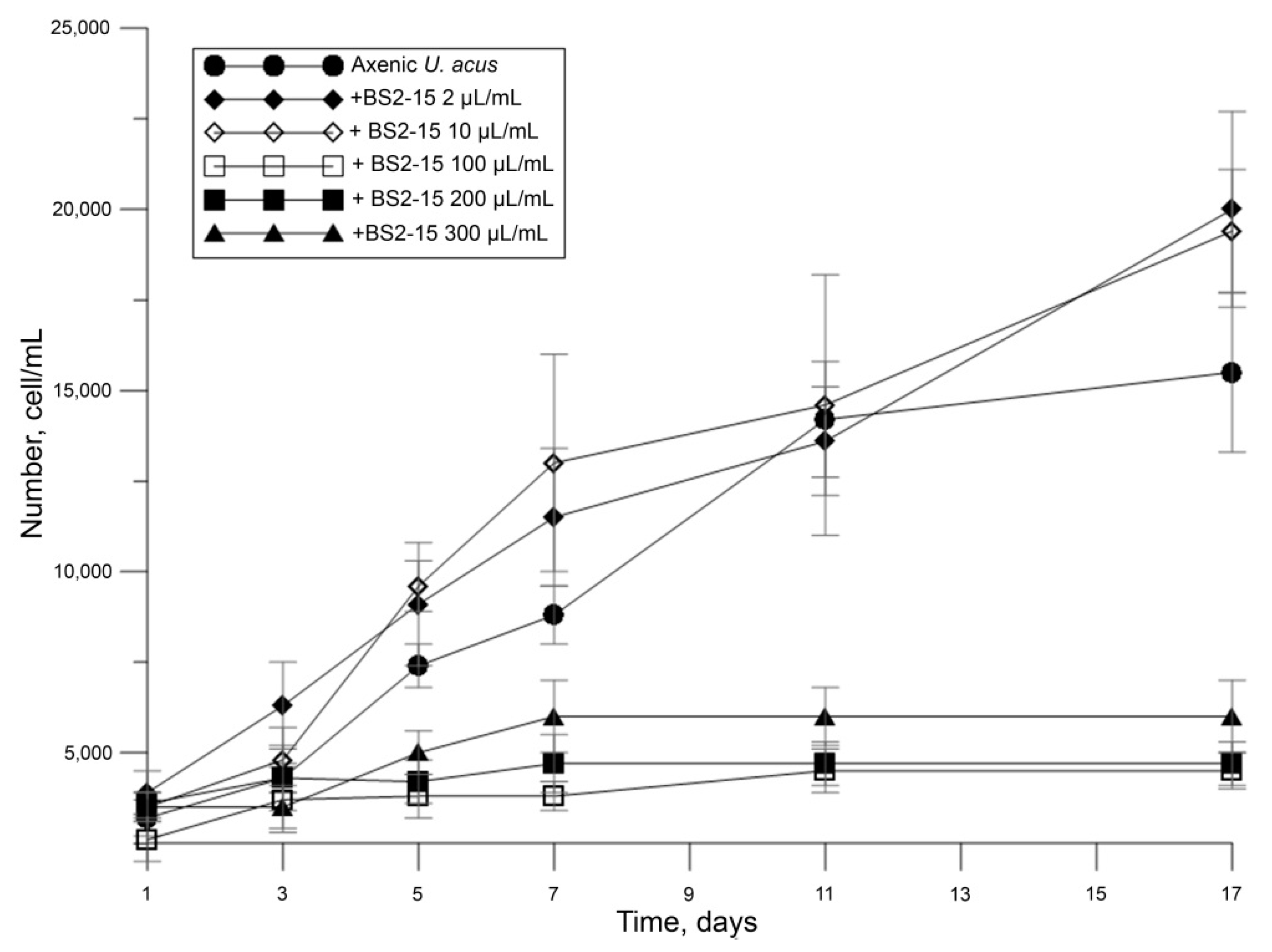
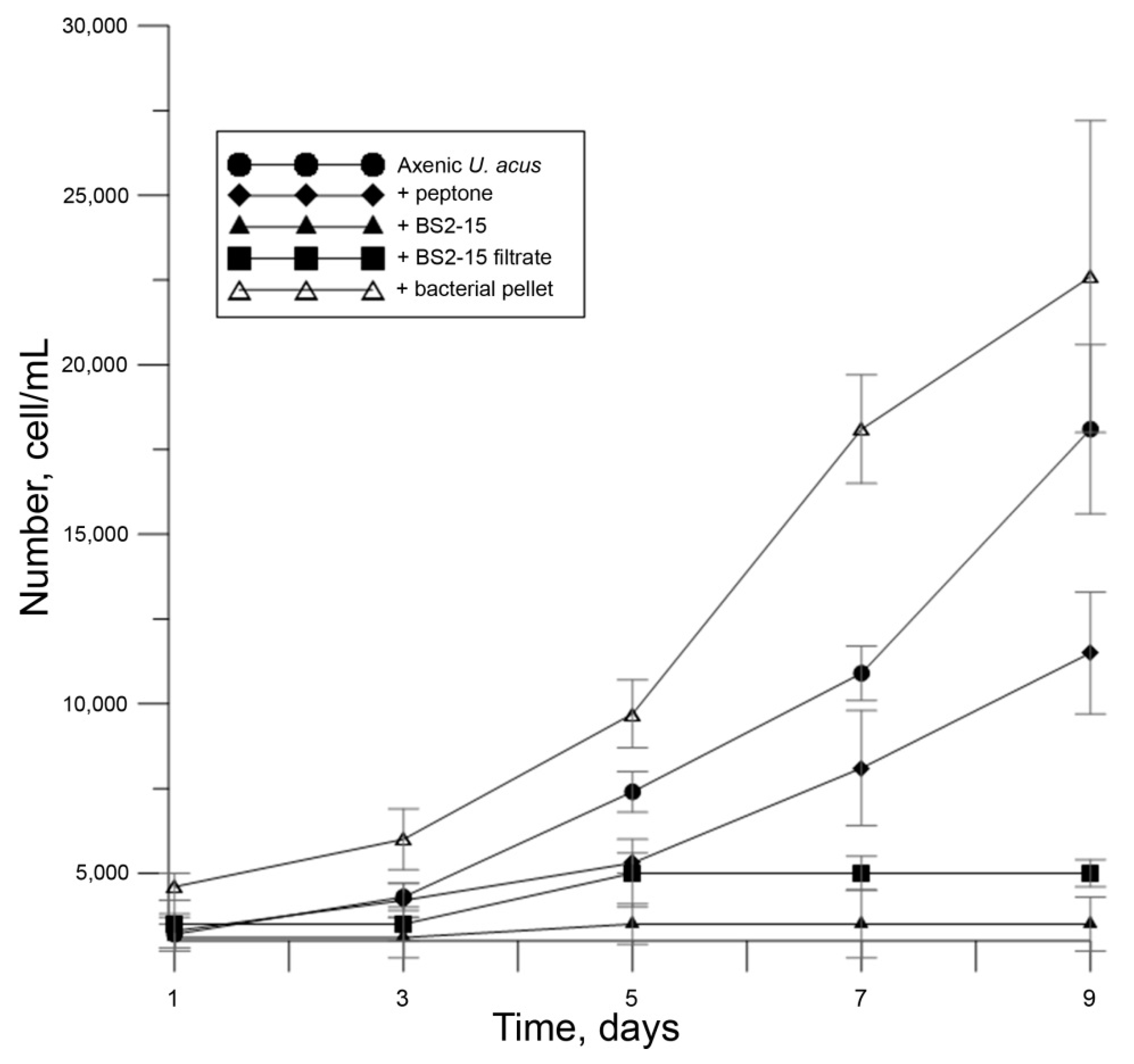
2.4. Determination of Algicidal Activity
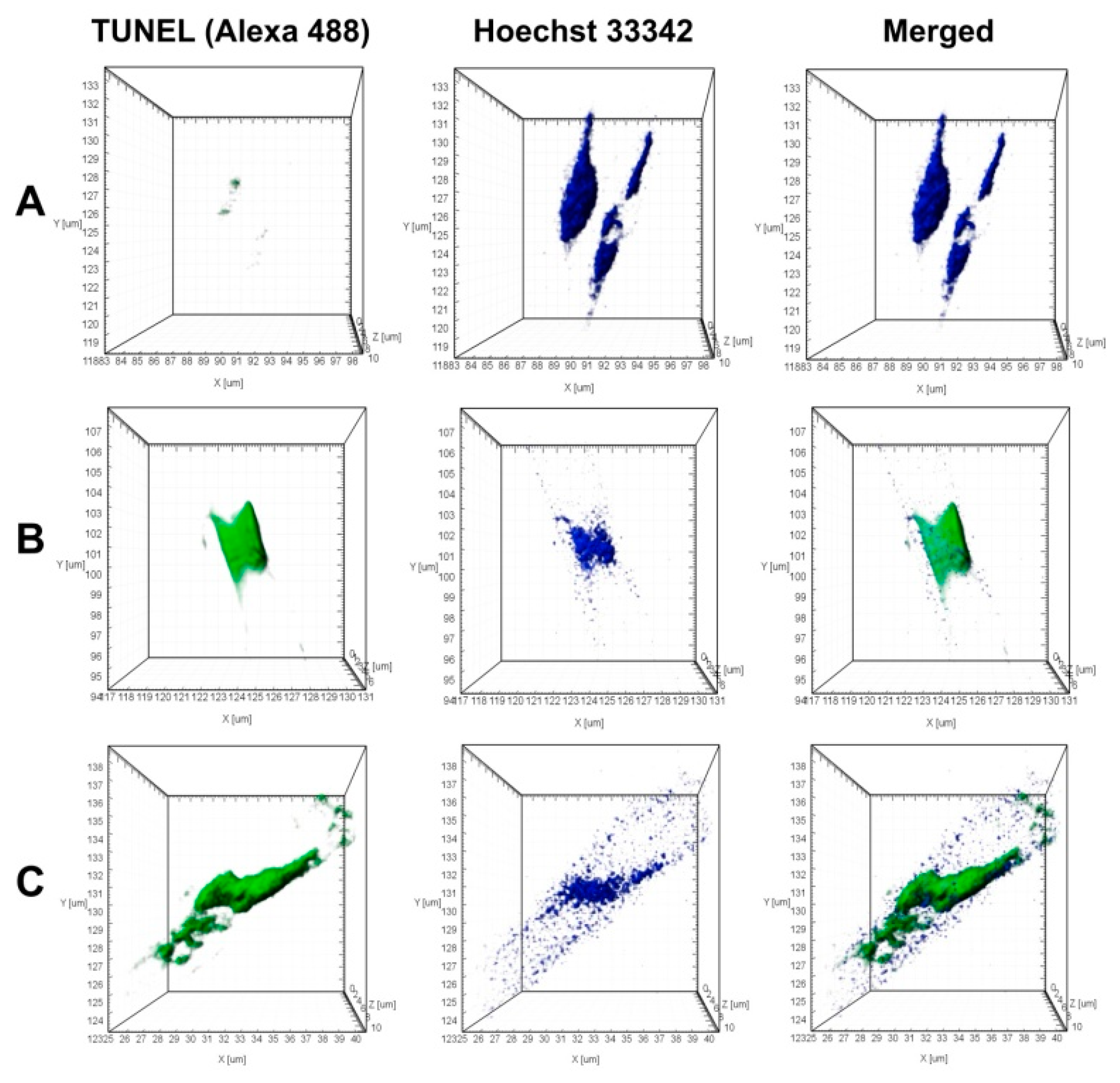
2.5. TUNNEL Assay
2.6. Lipid Droplets Analysis Using BODIPY
2.7. Fluorescent Microscopy
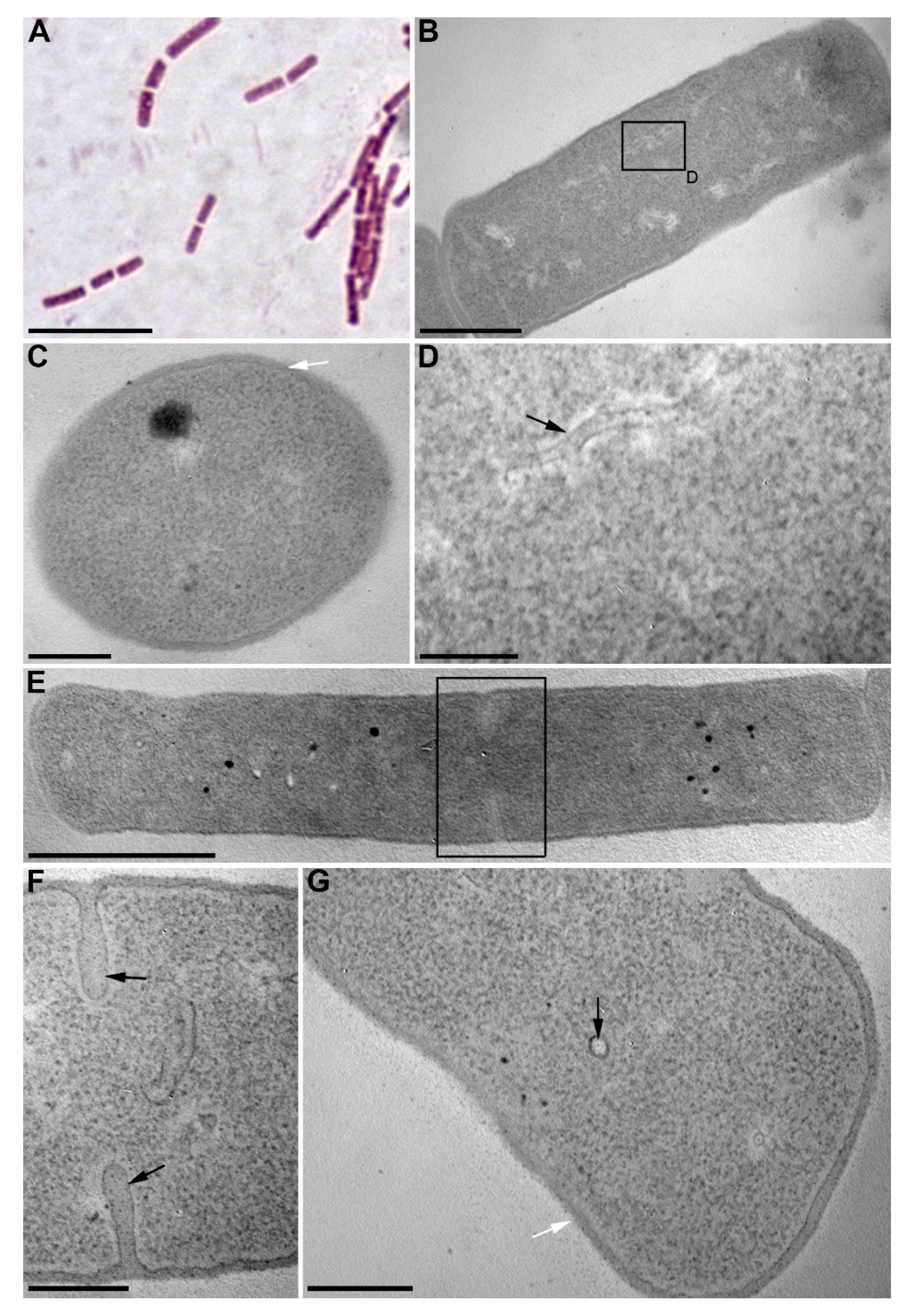
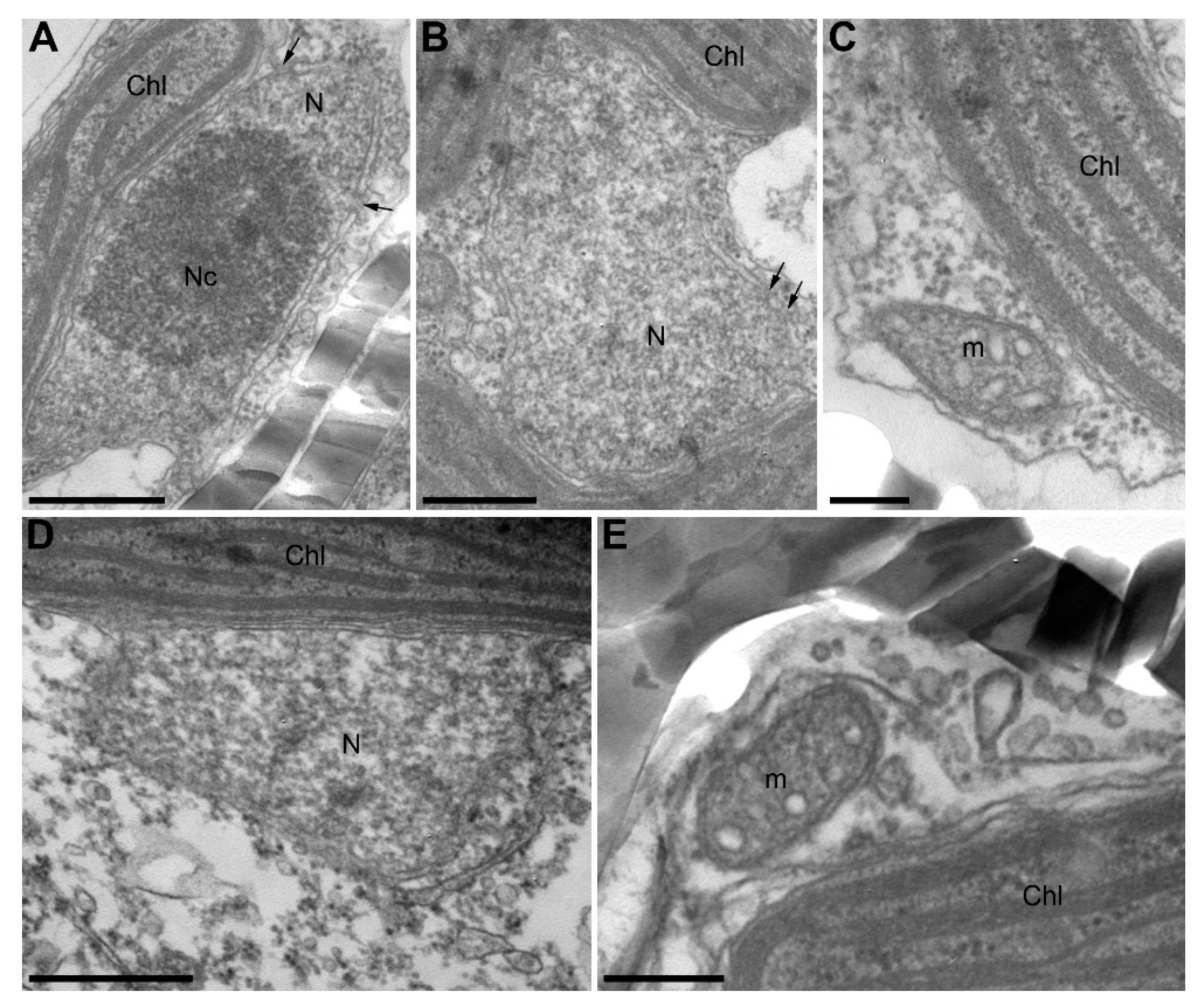
2.8. Transmission Electron Microscopy
2.9. Scanning Electron Microscopy
2.10. Statistical Analysis
3. Results and Discussion
3.1. Isolation and Identification of Algicidal Bacterium
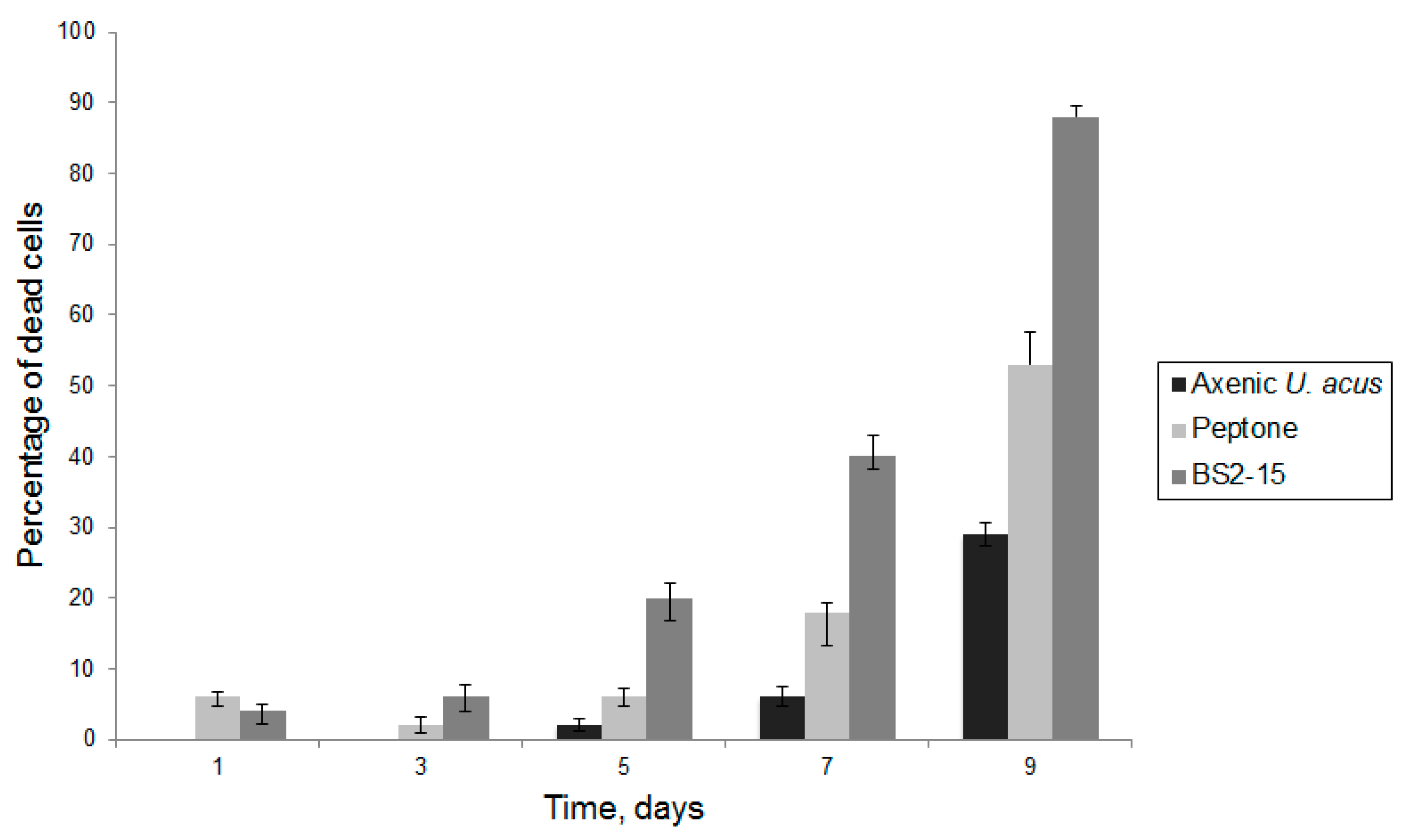
3.2. Algicidal Effect of B. mycoides BS2-15 on U. acus BK280
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Geider, R.J.; Delucia, E.H.; Falkowski, P.G.; Finzi, A.C.; Grime, J.P.; Grace, J. Primary productivity of planet earth: Biological determinants and physical constraints in terrestrial and aquatic habitats. Glob. Chang. Biol. 2001, 7, 849–882. [Google Scholar] [CrossRef]
- Nelson, D.M.; Tréguer, P.; Brzezinski, M.A.; Leynaert, A.; Quéguiner, B. Production and dissolution of biogenic silica in the ocean: Revised global estimates, comparison with regional data and relationship to biogenic sedimentation. Glob. Biogeochem. Cycles 1995, 9, 359–372. [Google Scholar] [CrossRef]
- Ragueneau, O.; Tréguer, P.; Leynaert, A.; Anderson, R.F.; Brzezinski, M.A.; DeMaster, D.J. A review of the Si cycle in the modern ocean: Recent progress and missing gaps in the application of biogenic opal as a paleoproductivity proxy. Glob. Planet. Chang. 2000, 26, 317–365. [Google Scholar] [CrossRef]
- Amin, S.A.; Parker, M.S.; Armbrust, E.V. Interactions between diatoms and bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 667–684. [Google Scholar] [CrossRef] [Green Version]
- Mayali, X.; Azam, F. Algicidal bacteria in the sea and their impact on algal blooms. J. Eukaryot. Microbiol. 2004, 51, 139–144. [Google Scholar] [CrossRef]
- Ramanan, R.; Kim, B.H.; Cho, D.H.; Oh, H.M.; Kim, H.S. Algae-bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Mitsutani, A.; Yamasaki, I.; Kitaguchi, H.; Kato, J.; Ueno, S.; Ishida, Y. Analysis of algicidal proteins of a diatom-lytic marine bacterium Pseudoalteromonas sp. strain A25 by two-dimensional electrophoresis. Phycologia 2001, 40, 286–291. [Google Scholar] [CrossRef]
- Kang, Y.H.; Kim, J.D.; Kim, B.H.; Kong, D.S.; Han, M.S. Isolation and characterization of a bio-agent antagonistic to diatom, Stephanodiscus hantzschii. J. Appl. Microbiol. 2005, 98, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Zakharova, Y.R.; Kurilkina, M.I.; Likhoshvay, Y.V.; Shishlyannikov, S.M.; Kalyuzhnaya, O.V.; Petrova, D.P. Effect of bacteria from the bottom water layer of Lake Baikal on degradation of diatoms. Paleontol. J. 2013, 47, 1030–1034. [Google Scholar] [CrossRef]
- Nicholson, W.L.; Munakata, N.; Horneck, G.; Melosh, H.J.; Setlow, P. Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments. Microbiol. Mol. Biol. Rev. 2000, 64, 548–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, D.; Dworkin, J. Recent progress in Bacillus subtilis sporulation. FEMS Microbiol. Rev. 2012, 36, 131–148. [Google Scholar] [CrossRef] [Green Version]
- Di Franco, C.; Beccari, E.; Santini, T.; Pisaneschi, G.; Tecce, G. Colony shape as a genetic trait in the pattern-forming Bacillus mycoides. BMC Microbiol. 2002, 2, 33. [Google Scholar] [CrossRef]
- Sugiura, H.; Nagase, A.; Oiki, S.; Mikami, B.; Watanabe, D.; Hashimoto, W. Bacterial inducible expression of plant cell wall-binding protein YesO through conflict between Glycine max and saprophytic Bacillus subtilis. Sci. Rep. 2020, 10, 18691. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, L.K.; Jackson, M.A. Clarification of the Taxonomy of Bacillus mycoides. Microbiol. Soc. 1995, 45, 46–49. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Zhang, J.; Zhao, X.; Wang, P.; Tong, M.; Glibert, P.M. Allelopathic inhibition by the bacteria Bacillus cereus BE23 on growth and photosynthesis of the Macroalga Ulva prolifera. J. Mar. Sci. Eng. 2020, 8, 718. [Google Scholar] [CrossRef]
- Wu, L.; Wu, H.; Chen, L.; Xie, S.; Zang, H.; Borriss, R.; Gao, X. Bacilysin from Bacillus amyloliquefaciens FZB42 has specific bactericidal activity against harmful algal bloom species. AEM 2014, 80, 7512–7520. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Lee, D.S.; Jeong, S.Y.; Lee, W.J.; Lee, M.S. Isolation and characterization of a marine algicidal bacterium against the harmful raphidophyceae Chattonella marina. J. Microbiol. 2009, 47, 9–18. [Google Scholar] [CrossRef]
- Hou, S.; Shu, W.; Tan, S.; Zhao, L.; Yin, P. Exploration of the antioxidant system and photosynthetic system of a marine algicidal Bacillus and its effect on four harmful algal bloom species. Can. J. Microbiol. 2015, 62, 49–59. [Google Scholar] [CrossRef]
- Huang, J.S.; Peng, Y.H.; Chung, K.R.; Huang, J.W. Suppressive efficacy of volatile compounds produced by Bacillus mycoides on damping-off pathogens of cabbage seedlings. J. Agric. Sci. 2018, 156, 795–809. [Google Scholar] [CrossRef]
- Zakharova, Y.R.; Galachyants, Y.P.; Kurilkina, M.I.; Likhoshvay, A.V.; Petrova, D.P.; Shishlyannikov, S.M.; Ravin, N.V.; Mardanov, A.V.; Beletsky, A.B.; Likhoshway, Y.V. The Structure of microbial community and degradation of diatoms in the deep near-bottom layer of Lake Baikal. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Zakharova, Y.R.; Adel’shin, R.V.; Parfenova, V.V.; Bedoshvili, Y.D.; Likhoshway, Y.V. Taxonomic characterization of the microorganisms associated with the cultivable diatom Synedra acus from Lake Baikal. Microbiology 2010, 79, 679–687. [Google Scholar] [CrossRef]
- Galachyants, Y.P.; Zakharova, Y.R.; Petrova, D.P.; Morozov, A.A.; Sidorov, I.A.; Marchenkov, A.M.; Logacheva, M.D.; Markelov, M.L.; Khabudaev, K.V.; Likhoshway, Y.V.; et al. Sequencing of the complete genome of an araphid pennate diatom Synedra acus subsp. radians from Lake Baikal. Dokl. Biochem. Biophys. 2015, 461, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Galachyants, Y.P.; Morozov, A.; Mardanov, A.V.; Beletsky, A.V.; Ravin, N.V.; Petrova, D.P.; Likhoshway, Y.V. Complete chloroplast genome sequence of freshwater araphid pennate diatom alga Synedra acus from Lake Baikal. Int. J. Biol. 2012, 4, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Galachyants, Y.P.; Zakharova, Y.R.; Volokitina, N.A.; Morozov, A.A.; Likhoshway, Y.V.; Grachev, M.A. De novo transcriptome assembly and analysis of the freshwater araphid diatom Fragilaria radians, Lake Baikal. Sci. Data 2019, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharitonenko, K.V.; Bedoshvili, Y.D.; Likhoshway, Y.V. Changes in the micro- and nanostructure of siliceous frustule valves in the diatom Synedra acus under the effect of colchicine treatment at different stages of the cell cycle. J. Struct. Biol. 2015, 190, 73–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, D.P.; Bedoshvili, Y.D.; Zakharova, Y.R.; Volokitina, N.A.; Likhoshway, Y.V.; Grachev, M.A. Changes in valve morphology of two pennate diatom species during long-term culture. Acta Biol. Sib. 2020, 6, 669–678. [Google Scholar] [CrossRef]
- Thompson, A.S.; Rhodes, J.C.; Pettman, I. Culture Collection of Algae and Protozoa: Catalogue of Strains; Titus Wilson and Son: Kendal, UK, 1988. [Google Scholar]
- Shishlyannikov, S.M.; Zakharova, Y.R.; Volokitina, N.A.; Mikhailov, I.S.; Petrova, D.P.; Likhoshway, Y.V. A procedure for establishing an axenic culture of the diatom Synedra acus subsp. radians (Kütz.) Skabibitsch. From Lake Baikal. Limnol. Oceanogr. Methods 2011, 9, 478–484. [Google Scholar] [CrossRef]
- Marmur, J. A procedure for the isolation of deoxyribonucleic acid from micro-organisms. J. Mol. Biol. 1961, 3, 208–218. [Google Scholar] [CrossRef]
- Johnson, M.; Zaretskaya, I.; Raytselis, Y.; Merezhuk, Y.; McGinnis, S.; Madden, T.L. NCBI BLAST: A better web interface. Nucleic Acids Res. 2008, 36, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Bruckner, C.G.; Rehm, C.; Grossart, H.P.; Kroth, P.G. Growth and release of extracellular organic compounds by benthic diatoms depend on interactions with bacteria. Environ. Microbiol. 2011, 13, 1052–1063. [Google Scholar] [CrossRef] [PubMed]
- Van Tol, H.M.; Amin, S.A.; Armbrust, E.V. Ubiquitous marine bacterium inhibits diatom cell division. ISME J. 2017, 11, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Bedoshvili, Y.D.; Popkova, T.P.; Likhoshway, Y.V. Chloroplast structure of diatoms of different classes. Cell Tissue Biol. 2009, 3, 297–310. [Google Scholar] [CrossRef]
- Pavlova, O.N.; Bukin, S.V.; Kostyreva, E.A.; Moskvin, V.I.; Manakov, A.Y.; Morozov, I.V.; Galachyants, Y.P.; Khabuev, A.V.; Zemskaya, T.I. Experimental transformation of organic matter by the microbial community from the bottom sediments of Akademichesky Ridge (Lake Baikal). Russ. Geol. Geophys. 2019, 60, 926–937. [Google Scholar] [CrossRef]
- Zulu, N.N.; Zienkiewicz, K.; Vollheyde, K.; Feussner, I. Current trends to comprehend lipid metabolism in diatoms. Prog. Lipid Res. 2018, 70. [Google Scholar] [CrossRef]
- Abida, H.; Dolch, L.-J.; Mei, C.; Villanova, V.; Conte, M.; Block, M.A.; Bastien, G.F.O.; Tirichine, L.; Bowler, C.; Rébeillé, F.; et al. Membrane glycerolipid remodeling triggered by nitrogen and phosphorus starvation in Phaeodactylum tricornutum. Plant Physiol. 2015, 167, 118–136. [Google Scholar] [CrossRef] [Green Version]
- Conte, M.; Lupette, J.; Seddiki, K.; Mei, C.; Dolch, L.-J.; Gros, V.; Barette, C.; Rébeillé, F.; Jouhet, J.; Maréchal, E. Screening for biologically annotated drugs that trigger triacylglycerol accumulation in the diatom Phaeodactylum. Plant Physiol. 2018, 177, 532–552. [Google Scholar] [CrossRef] [Green Version]
- Opute, F.I. Studies on fat accumulation in Nitzschia palea Kütz. Ann. Bot. 1974, 38, 889–902. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | BS2-15 | B. mycoides |
|---|---|---|
| Motility | − | − |
| Cell morphology | rod-shaped | rod-shaped |
| Cell diameter, µm | 0.75–1.3 | >1 µm |
| Cell length, µm | 1.8–4.5 | nd |
| Gram status | + | + |
| Endospores | + | + |
| Elliptic | + | + |
| Cylindrical | − | − |
| Spherical | − | − |
| Enzymatic activity | ||
| Catalase | + | + |
| Caseinase | − | + |
| Gelatinase | + | + |
| Lecithinase | + | + |
| Lypase | + | nd |
| Amylase | − | + |
| Acid production from carbohydrates: | ||
| Glucose | + | + |
| Galactose | − | d |
| Maltose | + | + |
| Rafinose | − | − |
| Lactose | − | − |
| Sucrose | + | d |
| Fructose | + | d |
| Rhamnose | − | − |
| Xylose | − | − |
| Arabinose | − | − |
| Dulcite | − | − |
| Inose | − | − |
| Sorbite | − | − |
| Mannite | − | − |
| β-galactosidase | − | d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bedoshvili, Y.; Bayramova, E.; Sudakov, N.; Klimenkov, I.; Kurilkina, M.; Likhoshway, Y.; Zakharova, Y. Impact of Algicidal Bacillus mycoides on Diatom Ulnaria acus from Lake Baikal. Diversity 2021, 13, 469. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100469
Bedoshvili Y, Bayramova E, Sudakov N, Klimenkov I, Kurilkina M, Likhoshway Y, Zakharova Y. Impact of Algicidal Bacillus mycoides on Diatom Ulnaria acus from Lake Baikal. Diversity. 2021; 13(10):469. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100469
Chicago/Turabian StyleBedoshvili, Yekaterina, Elvira Bayramova, Nikolay Sudakov, Igor Klimenkov, Maria Kurilkina, Yelena Likhoshway, and Yulia Zakharova. 2021. "Impact of Algicidal Bacillus mycoides on Diatom Ulnaria acus from Lake Baikal" Diversity 13, no. 10: 469. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100469