An Overview of Subterranean Biodiversity Hotspots




1
Department of Environmental Science, American University, 4400 Massachusetts Ave. NW, Washington, DC 20016, USA
2
Institut de Systématique, Évolution, Biodiversité (ISYEB), UMR7205, CNRS, Muséum National d’Histoire Naturelle, Sorbonne Université, EPHE, 45 Rue Buffon, 75005 Paris, France
3
Research Center of the Slovenian Academy of Sciences and Arts, Karst Research Institute, Novi Trg 2, SI-1000 Ljubljana, Slovenia
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(10), 487; https://0-doi-org.brum.beds.ac.uk/10.3390/d13100487
Submission received: 22 August 2021
/
Accepted: 23 August 2021
/
Published: 6 October 2021
(This article belongs to the Special Issue Hotspots of Subterranean Biodiversity)
1. Introduction
Riding a wave of interest in biodiversity patterns in surface-dwelling communities, in 2000, Culver and Sket [1] published a paper listing 20 caves and karst wells with 20 or more known species. At the time of their study, it was widely recognized that because of the extremely narrow distribution of most cave specialists, the overall species richness of a region or country was the result of the accumulation of species from a number of different caves, and that average single-site species richness (α-diversity) was a minor component of overall species richness (γ-diversity) [2]. This has since been confirmed in a number of large-scale studies of subterranean species diversity, especially in Europe [3,4,5,6]. Although the number of studies measuring both α-diversity (local diversity) and β-diversity (between-site diversity) has grown in the intervening 20+ years, they have largely been confined to Europe and, to a lesser extent, North America [3,4,5,6,7], mainly because it requires extensive datasets for about 100 sites for these to be estimated [6,8]. However, it is also well known that considerable subterranean species α-diversity lies outside these regions, including in Australia [9,10], Brazil [11,12], China [13], South East Asia [14], and Mexico [15], in part as a result of a search for additions to the list of hotspot caves reported by Sket and Culver.
The collection of papers in this Special Issue of Diversity—“Hotspots of Subterranean Biodiversity”—expands and enriches previous hotspot lists [2,14,16,17]. These hotspots may not capture all regions of high subterranean species richness; all of these hotspot caves are worthy of study and protection in their own right, and any protection strategy should include these exceptional sites as well as regional areas of species richness. In total, papers in this Special Issue cover 13 of 22 sites known to harbor at least 25 obligate subterranean species. Culver and Sket’s original list of 20 has become too lengthy to be of much use; thus, we raised the bar to 25. One other cave is included in this Special Issue because of its special geographical position distant from any other hotspot site—Ganxiao Dong Cave in China [18]. However, we did not include it in our summary of hotspot caves.
2. Overview of Hotspot Subterranean Sites
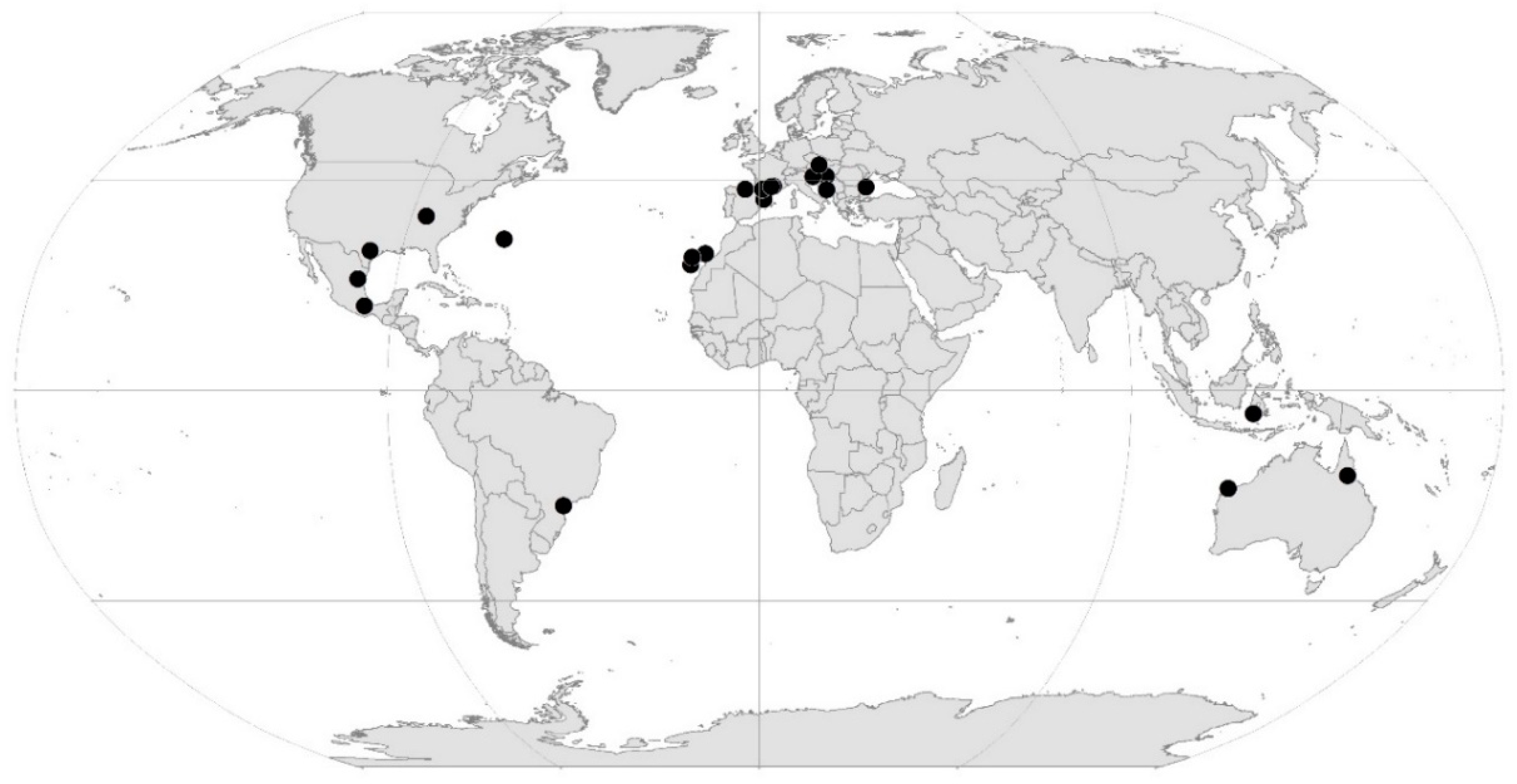
In Table 1, we list the 22 known sites with at least 25 obligate subterranean species, and they are also shown on the map in Figure 1. Two of these sites (Ojo Guareña in Spain and Sistema Huautla in Mexico) have not been previously noted in any list as hotspot caves. There may be additional non-cave subterranean sites, particularly in hyporheic habitats, but these data have for the most part not been assembled into species lists for individual sites. In addition, there is the difficulty in deciding what constitutes a single site—is it just one Bou-Rouch pump site or a stream reach (e.g., a riffle)?
Several additional sites are worth mentioning because it is likely that if further collecting and identification is performed, they will have 25 stygobionts plus troglobionts. Trajano et al. [12] report a highly diverse troglobiotic fauna in a Brazilian karst region, with 29 troglobionts reported from 11 caves in a 25 km2 region. This record is noteworthy both for its geographic position and the fact that the caves are in limestone, even though no one cave system has 25 troglobionts. No hotspot caves are reported from Africa, but Deharveng and Bedos, in an ongoing study, have reported 30 troglobionts and 3 stygobionts known from an Algerian karst area—Djurdjura—in an area no more than 50 km in length. Especially noteworthy is that there are many species restricted to karstic holes with permanent snow above 2000 m, especially beetles and Collembola, which exhibit eye and pigment reduction, most living in caves as well. In Ganxiao Dong in China, a total of 19 troglobionts and 1 stygobiont have been reported [18], and it is likely that many more species will be found, especially because multi-taxa collection is in its infancy. The fauna is noteworthy because of the high level of morphological specialization, especially in Coleoptera and Myriapoda.
Outside of the Dinaric mountains, none of the sites were highly ranked (i.e., in the top ten) with respect to both their aquatic fauna and terrestrial fauna. Postojna Planina Cave system is by far the richest known subterranean site with respect to species richness; it ranks first with respect to both aquatic and terrestrial species richness [19]. Some sites had no terrestrial fauna (San Marcos well in Texas, Jameos del Agua in the Canary Islands, Walsingham Cave in Bermuda, Triadou Aquifer, and Robe Valley wells in Australia) and others had no aquatic fauna (Huautla in Mexico, Cueva de Felipe Reventón and Cueva del Viento-Sobrado in the Canary Islands). All continents are represented by at least one site, except for Africa.
When the aquatic and terrestrial fauna are considered separately, a different pattern emerges for the two habitats. For aquatic species, the top ten sites include six anchialine or deep phreatic sites, three sites in the Dinaric mountains, one site in the Cantabrian Mountains of Spain. The Dinaric and Cantabrian sites are within the ridge of high-diversity regions in Europe [4,6]. All of these sites are likely to be enhanced in available organic carbon and nutrients relative to other sites, but historical reasons in the case of the Dinaric sites may be important as well [4]. In particular, their proximity to the Mediterranean Sea, a source of colonists, especially during the Messinian salinity crisis, may be important.
For terrestrial species, the sites are more scattered outside the Dinaric mountains, which is the location of three of the five richest terrestrial caves (Table 1). The lava tubes on Tenerife in the Canary Islands are the site of two of the ten species-rich terrestrial caves, as are two caves in Mexico; the other top ten are scattered in Australia, Brazil, Sulawesi, and the United States. The mesas near the Robe Valley in Australia are very species rich, but the reported subterranean fauna includes the soil fauna [34], making comparisons impossible. It is difficult to determine what unites these terrestrial sites, because they are a combination of lava tubes and karst caves, with very different levels of organic carbon and nutrients. It is, however, noteworthy that tropical or subtropical sites are relatively more represented than for aquatic fauna.
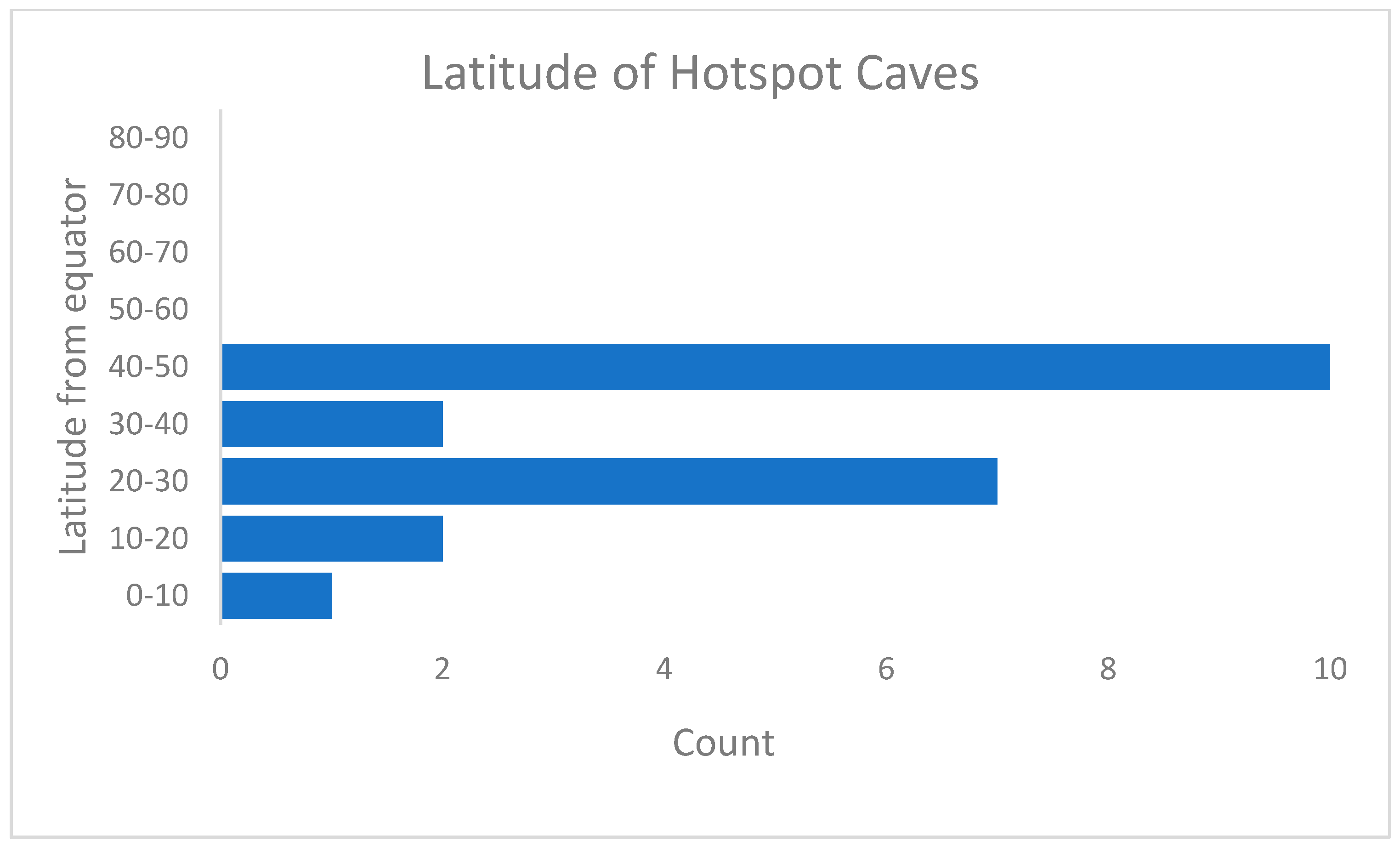
The latitudinal distribution of hotspot sites shows an unexpectedly bimodal distribution (Figure 2). The majority of sites are in the temperate zone, between 40 and 50 degrees north or south of the equator. This corresponds, at least in the northern hemisphere, to the region traditionally held to be the area of highest subterranean species richness, a result often attributed to the effects of repeated Pleistocene glaciations [36]. The second mode is in the sub-tropical and sub-temperate regions (between 20 and 30 degrees north and south of the equator). These sites are mostly lava tubes and wells connected to chemoautotrophic zones (Table 1). None of the sites are located in the arid tropics, also suggesting that food availability may be an important driver of species richness.
3. Caveats and Challenges
3.1. Differences in Working Definitions of Troglobionts and Stygobionts
Although there are many nuances [14,37,38,39], troglobionts and stygobionts are considered to be species exclusively found in subterranean habitats, which usually, but not always, show convergent morphological modification to subterranean life, often called troglomorphies [40]. Troglomorphies include, but are not limited to, reduced or absent eyes and pigment. For areas and groups with a well-studied surface fauna, it is possible to detect species occurring in subterranean habitats with well-developed eyes and pigment, but absent from the surface. In many, perhaps most cases, these are recent isolates in subterranean habitats. The difficulty is that in areas where the surface fauna is not well studied, such as most of the tropics, it is impossible to ascertain that such non-troglomorphic species are troglobiotic (or stygobiotic) species. Therefore, numbers of troglobiotic and stygobiotic species may be underestimated or overestimated in these sites. Such species may be common and the uncertainty of the ecological assignment (e.g., troglobiont or troglophile) is a bias. Deharveng and Bedos [41] report that soil-inhabiting species (especially Collembola and Diplura) are commonly listed as troglobionts.
Some studies of poorly known fauna may be biased by the inclusion of too many species as troglobionts and stygobionts if the operational criteria for inclusion are eyelessness and lack of pigment. Many litter- and soil-dwelling species are eyeless and depigmented, and some major taxonomic groups, e.g., Symphyla or the highly diversified Onychiuridae (Collembola) species, are entirely eyeless and depigmented regardless of habitat [29,42]. Additionally, some troglophilic species have populations that have reduced eyes and pigment. Sometimes these are recognized as subspecies [43], but if not, they are usually not included in the lists.
A strong case could be made that the relevant communities to compare are the permanent, sustaining populations (troglobionts [stygobionts] and eutroglophiles [eustygophiles]) but such lists are rarely available. Eutroglophiles and eustygophiles (see [38] for definitions) may be hard to delineate from other non-troglobionts and non-stygobionts.
3.2. Differences in Taxonomic Groups, Habitats Sampled, and Sampling Techniques
Differences in taxonomic composition can be because of regional differences in the composition of the subterranean fauna or because of incomplete sampling and/or taxonomic description. The tropics and arid areas tend to be richer in Arachnida, with some minor arachnid orders entirely missing at higher latitudes [14,41]. Other cases are clearly ones of inadequate collection. The waters of epikarst are usually dominated by Harpacticoidea crustaceans, but they have not been sampled or inadequately sampled outside of Europe (but see [44]), and the fauna can be quite rich, with more than ten species in some Slovenian caves [45,46]. Among the species listed in Table 1, only Postojna Planina Cave System [19] and Ojo Guareña [23] have been thoroughly sampled for epikarst fauna. Other groups may be under-represented due to their small size. Among Collembola, the Neelidae have several species less than 0.5 mm long [41,47] and are probably much more common than existing records would indicate. Ostracoda have rarely been sampled, with only the Postojna Planina Cave System [19], Ojo Guareña [23] and San Marcos well [28] among the sites in Table 1 having been surveyed. Finally, there are often groups that are under-collected because they require special preservation techniques or there are no taxonomists to study them (the Racovitzan and Linnean shortfalls [48]). Planarians are a good example. Although many records of planarians are known, many, if not most, have not been identified to species, and few subterranean species have been described recently.
3.3. Described versus Undescribed Species
A particularly vexing problem in the analysis of species numbers in subterranean sites is the nearly universal problem of undescribed species. Except for Cueva de Felipe Reventón, Cueva del Viento-Sobrado, Walsingham Caves, and Mammoth Cave, all the hotspot caves listed in Table 1 include undescribed species, as do Bayliss Cave, Sistema Huautla, Towakkalak system and Ganxiao Dong included in this Special Issue. The percentage of undescribed species tends to be highest in tropical countries, with a rich fauna and a shorter tradition of taxonomic description, such as Brazil, but it is also high in countries such as Australia (Table 1).
The difficulty is that not all records reported as new species turn out to be new species. The West Virginia cave fauna provides an example of the problem. In 1976, Holsinger, Baroody, and Culver [49], listed 27 undescribed stygobionts and troglobionts. In a re-examination of the West Virginia cave fauna in 2007, Fong et al. [50] found that of the 27 undescribed species listed in 1976:
- 13 were described as new species;
- 6 were assigned to existing known stygobiotic and troglobiotic species;
- 8 remained unstudied.
This suggests a “discount rate” for undescribed species of 68% (13 of 19, because 6 were not new species after all), but this rate is likely to vary from region to region, and if it were applied to the data in Table 1, it would result in a bias against tropical regions and any other region with large numbers of undescribed species. Perhaps the best solution is to list both groups of species in any compilation of species richness but to indicate the number that remain undescribed, as we have done in Table 1.
Finally, there are questions about the quality of the taxonomic descriptions. This may be especially problematic for descriptions more than one century old, where the descriptions were very brief, and the range of diversity of different groups was largely unknown. This is, for example, a problem with mites described from Mammoth Cave in the 19th century, some of which have not been recollected [24].
4. Protection of Hotspot Caves
It is difficult to generalize, either about the threats or the level of protection for the hotspot caves listed in Table 1, at least beyond the existential threat that global warming poses for subterranean ecosystems [51,52]. In addition, a few generalities are possible. First, all sites are affected by surface processes. For caves, these inputs include infiltration through soil and epikarst, direct inputs from sinking streams, and direct human actions, including tourism.
Secondly, the area of the Earth’s surface that impacts a subterranean fauna is greater for the aquatic fauna than for the terrestrial fauna. At one extreme is the artesian well at San Marcos, Texas, which accesses groundwater in the Edwards Aquifer at a depth of 59.5 m, and the Edwards Aquifer itself, which covers an area of over 10,000 km2 [28]: water quality and quantity in the San Marcos well are potentially affected by events occurring through the Edwards Aquifer. At another extreme are the lava tubes in the Canary Islands. Most, if not all, of the organic input comes from tree roots as well as organic matter deposited from small cracks and crevices in the lava tube. In this case, the vulnerable zone is the projection of the cave onto the surface. In many cases, the limit of the vulnerable zone is unknown.
Thirdly, cave tourism is an important aspect of the state of affairs for a number of hotspot caves. Two caves in Table 1 have in excess of 100,000 visitors per year (Postojna Planina Cave System and Mammoth Cave). Others have smaller numbers of visitors, including wild cave tours (Walsingham Caves, Križna jama, Jameos del Agua, and Cueva del Viento Sobrado). Although cave tourism is often thought of in negative terms because of the attendant problems of light pollution, increases in CO2, structural modifications of the cave, etc., it can have a positive impact in that the economic value of a tourist cave makes major disruption unlikely. Additionally, the tourist part of the cave is typically a small fraction of the entire cave.
Furthermore, many of the hotspot caves are protected by some form of designation as park land, or protected area. Even if there is no official designation, there are often laws protecting caves in general [53,54]. However, the efficacy of these regulations and designations is never absolute, and at some level all sites face threats. For example, even Mammoth Cave, a U.S. National Park, faces a number of problems, ones due to actions taken outside the park, as well as past actions by the NPS itself [55]. On the obverse side, some caves are largely protected by their difficulty of access, by their length, as well as by the secrecy of some of their entrances, such as Sistema Huautla, one of the deepest caves (>1 km) in the world [56] or Coume Ouarnède [25].
Finally, there is not one template for threats or one prescription for the protection of subterranean hotspots. What works for a site in the United States probably will not work for a site in Brazil. Each site must be considered separately and carefully, and upon threat evaluation. Threat levels are the basic criteria to be used to derive protection measures. All the sites listed in Table 1 are worthy of protection as well as a source of regional and national heritage.
Author Contributions
Data compilation: D.C.C., L.D., T.P., and A.B.; Manuscript writing: D.C.C.; Editing: L.D., T.P., and A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Culver, D.C.; Sket, B. Hotspots of subterranean biodiversity in caves and wells. J. Cave Karst Stud. 2000, 62, 11–17. [Google Scholar]
- Gibert, J.; Deharveng, L. Subterranean ecosystems: A truncated functional diversity. Bioscience 2002, 52, 473–481. [Google Scholar] [CrossRef]
- Bregović, P.; Zagmajster, M. Understanding hotspots within a global hotspot—Identifying the drivers of regional species richness patterns in terrestrial subterranean habitats. Insect Conserv. Biodivers. 2016, 9, 268–281. [Google Scholar] [CrossRef]
- Zagmajster, M.; Eme, D.; Fišer, C.; Galassi, D.; Marmonier, P.; Stoch, F.; Cornu, J.; Malard, F. Geographic variation in range size and beta diversity of groundwater crustaceans: Insights from habitats with low thermal seasonality. Glob. Ecol. Biogeogr. 2014, 23, 1135–1145. [Google Scholar] [CrossRef]
- Malard, F.; Boutin, C.; Camacho, A.I.; Ferreira, D.; Michel, G.; Sket, B.; Stoch, F. Diversity patterns of stygobiotic crustaceans across multiple spatial scales in western Europe. Freshw. Biol. 2009, 54, 756–776. [Google Scholar] [CrossRef]
- Culver, D.C.; Deharveng, L.; Bedos, A.; Lewis, J.J.; Madden, M.; Reddell, J.R.; Sket, B.; Trontelj, P.; White, D. The mid-latitude biodiversity ridge in terrestrial cave fauna. Ecography 2006, 29, 120–128. [Google Scholar] [CrossRef]
- Christman, M.C.; Doctor, D.H.; Niemiller, M.L.; Weary, D.J.; Young, J.A.; Zigler, K.S.; Culver, D.C. Predicting the occurrence of cave-inhabiting fauna based on features of the Earth surface environment. PLoS ONE 2016, 11, e0160408. [Google Scholar] [CrossRef]
- Dole-Olivier, M.J.; Castellarini, F.; Coineau, N.; Galassi, D.M.P.; Martin, P.; Mori, N. Towards an optimal sampling strategy to assess groundwater biodiversity comparison across six European regions. Freshw. Biol. 2009, 54, 777–796. [Google Scholar] [CrossRef]
- Humphreys, W.H. Biodiversity patterns in Australia. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: New York, NY, USA, 2019; pp. 109–126. [Google Scholar]
- Guzick, M.T.; Austin, A.D.; Cooper, S.J.B.; Harvey, M.S.; Humphreys, W.F.; Bradford, T.; Eberhard, S.M.; King, R.A.; Leijs, R.; Muirhead, K.A.; et al. Is the Australian subterranean fauna uniquely diverse? Invert. Syst. 2010, 24, 407–418. [Google Scholar] [CrossRef] [Green Version]
- Souza-Silva, M.; Ferreira, R.L. The first two hotspots of subterranean biodiversity in South America. Subterr. Biol. 2016, 19, 1–21. [Google Scholar] [CrossRef]
- Trajano, E.; Gallão, J.E.; Bichuette, M.E. Spots of high diversity of troglobites in Brazil: The challenge of measuring subterranean diversity. Biodivers. Conserv. 2016, 25, 1805–1828. [Google Scholar] [CrossRef]
- Latella, L. Biodiversity: China. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: London, UK, 2019; pp. 127–135. [Google Scholar]
- Deharveng, L.; Bedos, A. Biodiversity in the tropics. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: London, UK, 2019; pp. 146–162. [Google Scholar]
- Palacios-Vargas, J.G.; Juberthie, C.; Reddell, J.R. México. Mundos Subterráneos 2014–2015, 25–26, 1–101. [Google Scholar]
- Culver, D.C.; Pipan, T. Biology of Caves and Other Subterranean Habitats, 1st ed.; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Culver, D.C.; Pipan, T. Subterranean ecosystems. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: Waltham, MA, USA, 2013; Volume 7, pp. 49–62. [Google Scholar]
- Huang, S.; Wei, G.; Wang, H.; Liu, W.; Bedos, A.; Deharveng, L.; Tian, M. Ganxiao Dong: A hotspot of cave biodiversity in northern Guangxi, China. Diversity 2021, 13, 355. [Google Scholar] [CrossRef]
- Zagmajster, M.; Polak, S.; Fišer, C. Postojna Planina Cave System in Slovenia, a hotspot of subterranean biodiversity and a cradle of speleobiology. Diversity 2021, 13, 271. [Google Scholar] [CrossRef]
- Lučić, I.; Sket, B. Vjetrenica. Pogled U Dušu Zemlje; Savez Speleologa Bosne I Hercegovine and Hrvatsko Biospeleološko Društvo: Zagreb, Croatia, 2003. [Google Scholar]
- Ozimec, R.; Lučić, I. The Vjetrenica cave (Bosnia & Herzegovina)–one of the world’s most prominent biodiversity hotspots for cave-dwelling fauna. Subt. Biol. 2009, 7, 17–24. [Google Scholar]
- Polak, S.; Pipan, T. The subterranean fauna of Križna jama, Slovenia. Diversity 2021, 13, 210. [Google Scholar] [CrossRef]
- Camacho, A.I.; Puch, C. Ojo Guareña, a hotspot of subterranean biodiversity in Spain. Diversity 2021, 13, 199. [Google Scholar] [CrossRef]
- Niemiller, M.L.; Helf, K.; Toomey, R.S. Mammoth Cave: A hotspot of subterranean biodiversity in the United States. Diversity 2021, 13, 373. [Google Scholar] [CrossRef]
- Faille, A.; Deharveng, L. The Coume Ouarnède system, a hotspot of subterranean biodiversity in Pyrenees (France). Diversity 2021, 13, 419. [Google Scholar] [CrossRef]
- Brad, T.; Iepure, S.; Sarbu, S.M. The chemoautotrophically based Movile Cave groundwater ecosystem, a hotspot of subterranean biodiversity. Diversity 2021, 13, 128. [Google Scholar] [CrossRef]
- Olivier, M.J.; Martin, D.; Bou, C.; Prié, V. Interprétation du suivi hydrobiologique de la faune stygobie, réalisé sur le système karstique des Cents Fonts lors du pompage d’essai. In Système Karstique des Cent Fonts. Simulation de Scénarios d’Exploitation et de Gestion de la Resource; Ladouche, B., Maréchal, J.C., Dörfliger, N., Lachassagne, P., Eds.; Bureau de Recherches Géologiques et Minières: Montpelier, France, 2006; pp. 127–142. [Google Scholar]
- Iliffe, T.M.; Calderón Gutiérrez. Bermuda’s Walsingham Caves: A global hotspot for anchialine stygobionts. Diversity 2021, 13, 352. [Google Scholar] [CrossRef]
- Hutchins, B.T.; Gibson, J.R.; Diaz, P.H.; Schwartz, B.F. Stygobiont diversity in the San Marcos Artesian Well and Edwards Aquifer groundwater ecosystem, Texas, USA. Diversity 2021, 13, 234. [Google Scholar] [CrossRef]
- Martínez, A.; Gonzalez, B.C. Volcanic anchialine habitats of Lanzarote. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 399–414. [Google Scholar]
- Oromí, P.; Socorro, S. Biodiversity in the Cueva del Vientzo lava tube system (Tenerife, Canary Islands). Diversity 2021, 13, 226. [Google Scholar] [CrossRef]
- Francke, O.F.; Monjaraz-Ruedas, R.; Cruz-López, J. Biodiversity of the Huautla Cave System, Oaxaca, Mexico. Diversity 2021, 13, 429. [Google Scholar] [CrossRef]
- Eberhard, S.M.; Howarth, F.G. Undara basalt flow, a hotspot of subterranean biodiversity, Queensland, Australia. Diversity 2021, 13, 326. [Google Scholar] [CrossRef]
- Clark, H.L.; Buzatto, B.A.; Halse, S.A. A hotspot of arid zone subterranean biodiversity: The Robe Valley in Western Australia. Diversity 2021, 13, 482. [Google Scholar] [CrossRef]
- Deharveng, L.; Rahmadi, C.; Suhardjono, Y.R.; Bedos, A. The Towakkalak System, a hotspot of subterranean biodiversity in Sulawesi, Indonesia. Diversity 2021, 13, 392. [Google Scholar] [CrossRef]
- Jeannel, R. Les Fossiles Vivants des Cavernes; Gallimard: Paris, France, 1944. [Google Scholar]
- Trajano, E.; de Carvalho, M.R. Towards a biologically meaningful classification of subterranean organisms: A critical analysis of the Schiner-Racovitza system from a historical perspective, difficulties of its application and implications for conservation. Subterr. Biol. 2017, 22, 1–26. [Google Scholar] [CrossRef]
- Sket, B. Can we agree on an ecological classification of subterranean animals. J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- Culver, D.C.; Pipan, T. Ecological and evolutionary classifications of subterranean organisms. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: London, UK, 2019; pp. 376–379. [Google Scholar]
- Christiansen, K.A. Proposition pour la classification des animaux cavernicoles. Spelunca 2019, 2, 75–78. [Google Scholar]
- Deharveng, L.; Bedos, A. Diversity of terrestrial invertebrates in subterranean environments. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 107–172. [Google Scholar]
- Peck, S.B. Eyeless arthropods of the Galapagos Islands, Ecuador: Composition and origin of the cryptozoic fauna of a young, tropical, oceanic archipelago. Biotropica 1990, 22, 366–381. [Google Scholar] [CrossRef]
- Schultz, G.A. Descriptions of new subspecies of Ligidium elrodii (Packard) comb. nov. with notes on other isopod crustaceans from caves in North America (Oniscoidea). Am. Midl. Nat. 1970, 84, 36–45. [Google Scholar] [CrossRef]
- Pipan, T.; Christman, M.C.; Culver, D.C. Dynamics of epikarst communities: Microgeographic pattern and environmental determinants of epikarst copepods in Organ Cave, West Virginia. Am. Midl. Nat. 2006, 156, 75–87. [Google Scholar] [CrossRef]
- Pipan, T. Epikarst–A Promising Habitat; Založba ZRC: Ljubljana, Slovenia, 2005. [Google Scholar]
- Pipan, T.; Culver, D.C.; Papi, F.; Kozel, P. Partitioning diversity in subterranean invertebrates: The epikarst fauna of Slovenia. PLoS ONE 2018, 13, e0185991. [Google Scholar] [CrossRef]
- Schneider, C.; Deharveng, L. First record of the genus Spinaethorax Papáč and Palacios-Vargas, 2016 (Collembola, Neelipleona, Neelidae) in Asia, with a new species from a Vietnamese cave. Eur. J. Taxon. 2017, 363, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Ficetola, C.F.; Canadoli, C.; Stoch, F. The Racovitzan impediment and the hidden biodiversity of unexplored environments. Conserv. Biol. 2019, 33, 214–216. [Google Scholar] [CrossRef]
- Holsinger, J.R.; Baroody, R.A.; Culver, D.C. The Invertebrate Cave Fauna of West Virginia; West Virginia Speleological Survey: Barrackville, WV, USA, 1976; No. 7. [Google Scholar]
- Fong, D.W.; Culver, D.C.; Hobbs, H.H., III; Pipan, T. The Invertebrate Cave Fauna of West Virginia, 2nd ed.; West Virginia Speleological Survey: Barrackville, WV, USA, 2007; No. 16. [Google Scholar]
- Mammola, S.; Cardoso, P.; Culver, D.C.; Deharveng, L.; Ferreira, R.L.; Fišer, C.; Galassi, D.M.P.; Griebler, C.; Halse, S.; Humphreys, W.F.; et al. Scientists’ warning on the conservation of subterranean ecosystems. BioScience 2019, 69, 641–650. [Google Scholar] [CrossRef] [Green Version]
- Mammola, S.; Piano, E.; Cardoso, P.; Vernon, P.; Domìnguez-Viller, D.; Culver, D.C.; Pipan, T.; Isaia, M. Climate change going deep: The effects of global climatic alterations on cave ecosystems. Anthr. Rev. 2019, 6, 98–116. [Google Scholar] [CrossRef]
- Niemiller, M.L.; Taylor, S.J.; Bichuette, M.E. Conservation of cave fauna, with emphasis on Europe and the Americas. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 451–478. [Google Scholar]
- Halse, S.J. Conservation and impact assessment of subterranean fauna in Australia. In Cave Ecology; Moldovan, O.T., Kováč, L., Halse, S., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 479–496. [Google Scholar]
- Olson, R.A. Environmental issues relevant to the Mammoth Cave area. In Mammoth Cave. A Human and Natural History; Hobbs, H.H., III, Olson, R.A., Winkler, E.G., Culver, D.C., Eds.; Springer Nature: Cham, Switzerland, 2017; pp. 265–275. [Google Scholar]
- Steele, C.W.; Shifflett, T.E. Huautla cave system (Sistema Huautla), Mexico. In Encyclopedia of Caves, 3rd ed.; White, W.B., Culver, D.C., Pipan, T., Eds.; Academic Press: New York, NY, USA, 2019; pp. 527–536. [Google Scholar]
Figure 1.
Map of hotspot sites listed in Table 1. Map prepared by Magda Aljančič, used with permission.
Figure 1.
Map of hotspot sites listed in Table 1. Map prepared by Magda Aljančič, used with permission.

Figure 2.
Distribution distances from the equator of the subterranean hotspot sites listed in Table 1.
Figure 2.
Distribution distances from the equator of the subterranean hotspot sites listed in Table 1.


{kind=link}
{kind=link}
Table 1.
All sites with 25 or more stygobionts plus troglobionts. Caves assessed in this special issue are marked with an asterisk.
Table 1.
All sites with 25 or more stygobionts plus troglobionts. Caves assessed in this special issue are marked with an asterisk.
| Stygo- bionts | Rank | Troglo-bionts | Rank | Total | Rank | No. Unde-scribed | Terrain and Hydrogeology | Latitude | YearAssessed | Reference | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperate | |||||||||||
| Postojna Planina System (SLO) * | 72 | 1 | 45 | 1 | 117 | 1 | 8 | karst | 45.7 | 2021 | [19] |
| Vjetrenica (B&H) | 42 | 7 | 39 | 2 | 81 | 2 | 3 | karst | 42.9 | 2009 | [20,21] |
| Križna Jama (SLO) * | 32 | 10 | 28 | 6 | 60 | 4 | 5 | karst | 45.7 | 2021 | [22] |
| Ojo Guareña (ESP) * | 46 | 5 | 8 | 54 | 6 | 23 | karst | 43.0 | 2021 | [23] | |
| Logarček (SLO) | 28 | 15 | 43 | 9 | ? | karst | 43.5 | 2000 | [1] | ||
| Mammoth Cave (USA) * | 17 | 32 | 4 | 49 | 7 | 0 | karst | 45.9 | 2021 | [24] | |
| Coume Ouarnède (FRA) * | 21 | 17 | 38 | 3 | karst | 43.0 | 2021 | [25] | |||
| Pestera de la Movile (RO) * | 13 | 25 | 38 | 4 | karst, chemoautotrophic | 37.2 | 2021 | [26] | |||
| Cent fonts (FRA) | 43 | 6 | 0 | 43 | 9 | 7 | karst, phreatic | 43.6 | 2006 | [27] | |
| Triadou aquifer (FRA) | 34 | 9 | 0 | 34 | ? | karst, phreatic | 43.7 | 2000 | [1] | ||
| Baget system (FRA) 1 | 24 | 9 | 33 | ? | karst, phreatic | 43.0 | 2000 | [1] | |||
| Subtemperate | |||||||||||
| Walsingham Cave (BER) * | 65 | 2 | 0 | 65 | 3 | 0 | karst, anchialine | 32.3 | 2021 | [28] | |
| San Marcos Artesian Well (USA) * | 55 | 3 | 0 | 55 | 5 | 16 | karst, phreatic, chemoautotrophic | 29.9 | 2021 | [29] | |
| Jameos del Agua (ESP) | 40 | 8 | 0 | 40 | 4 | lava tube, anchialine | 28.4 | 2018 | [30] | ||
| Cueva de Felipe Reventon (ESP) * | 0 | 38 | 3 | 38 | 0 | lava tube | 28.4 | 2021 | [31] | ||
| Cueva del Viento-Sobrado (ESP) * | 0 | 28 | 6 | 28 | 0 | lava tube | 28.4 | 2021 | [31] | ||
| Sub-tropical | |||||||||||
| Sistema Huautla (MEX) * | 0 | 27 | 9 | 27 | 10 | deep karst | 18.1 | 2021 | [32] | ||
| Undara Basalt Flow (AUS) *2 | 1 | 30 | 5 | 31 | 27 | lava tube | −18.2 | 2021 | [33] | ||
| Robe Valley wells (AUS) *3 | 48 | 4 | 0 | 48 | 8 | 18 | karst, phreatic | −21.6 | 2021 | [34] | |
| Sistema Purificacion (MEX) | 3 | 28 | 6 | 31 | ? | karst | 23.8 | 2019 | [14] | ||
| Areias system (BRA) | 6 | 22 | 28 | 14 | karst | −24.6 | 2016 | [11] | |||
| Tropical | |||||||||||
| Towakkalak (SUL) * | 10 | 26 | 10 | 36 | 17 | karst | −5.0 | 2021 | [35] |
1 Unpublished records of L. Deharveng show 30 stygobionts and 11 troglobionts from the Baget Basin, including Grotte de Sainte-Catherine. Two species are undescribed. 2 Numbers are for the Undara Basalt Flow as a whole. Bayliss Cave, the largest cave, has 23 troglobionts and 1 stygobiont. 3 Clark et al. [34] describe a nearby terrestrial area but it includes both deep soil and MSS (milieu souterrain superficiel) components, and is not directly comparable to the other sites.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Culver, D.C.; Deharveng, L.; Pipan, T.; Bedos, A. An Overview of Subterranean Biodiversity Hotspots. Diversity 2021, 13, 487. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100487
AMA Style
Culver DC, Deharveng L, Pipan T, Bedos A. An Overview of Subterranean Biodiversity Hotspots. Diversity. 2021; 13(10):487. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100487
Chicago/Turabian StyleCulver, David C., Louis Deharveng, Tanja Pipan, and Anne Bedos. 2021. "An Overview of Subterranean Biodiversity Hotspots" Diversity 13, no. 10: 487. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100487
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.