
Emerging Ecotone and Microbial Community of a Sulfidic Spring in the Reka River near Škocjanske Jame, Slovenia

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
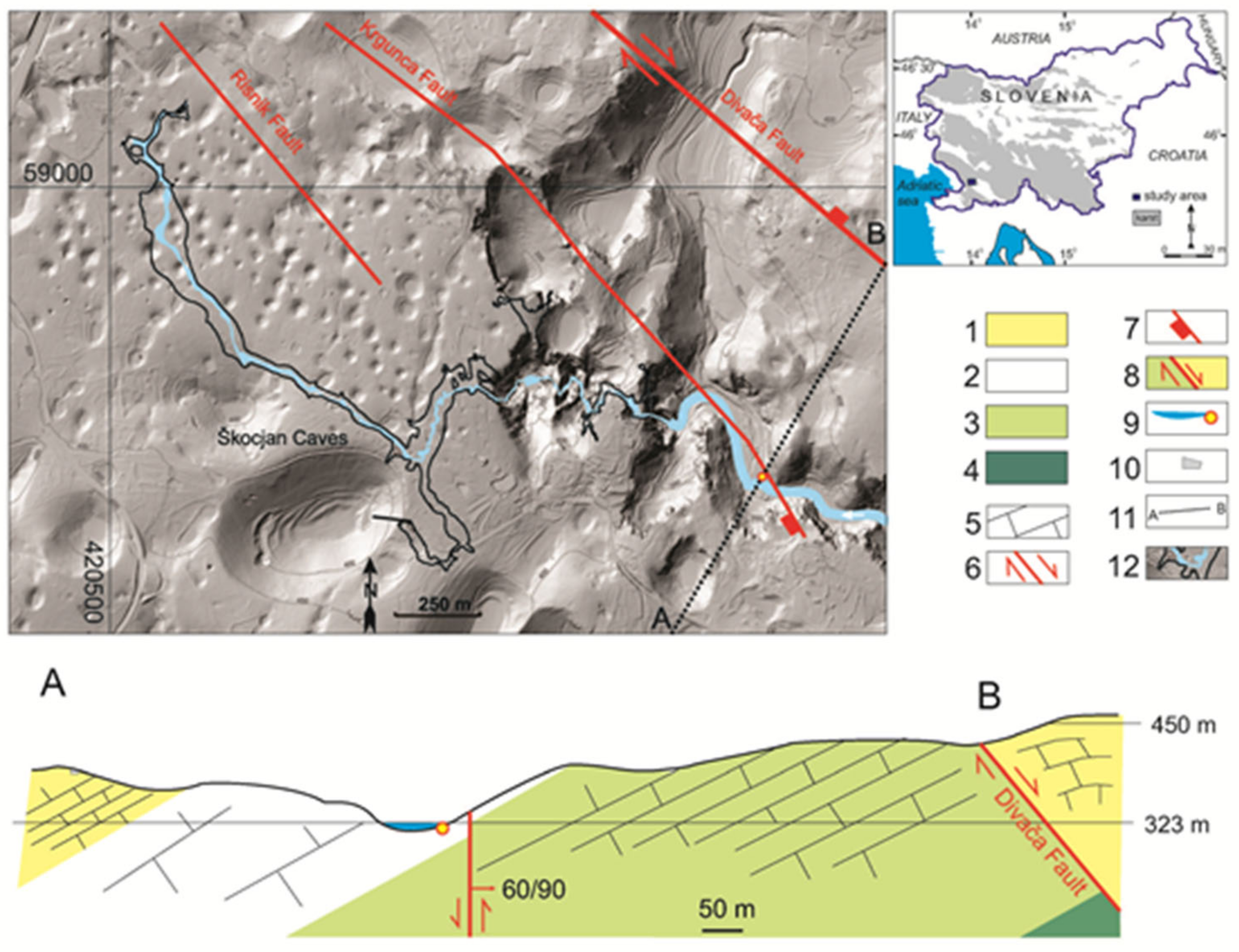
2.1. Discovery and Site Description
2.2. Geology of the Smrdljivec Spring Area

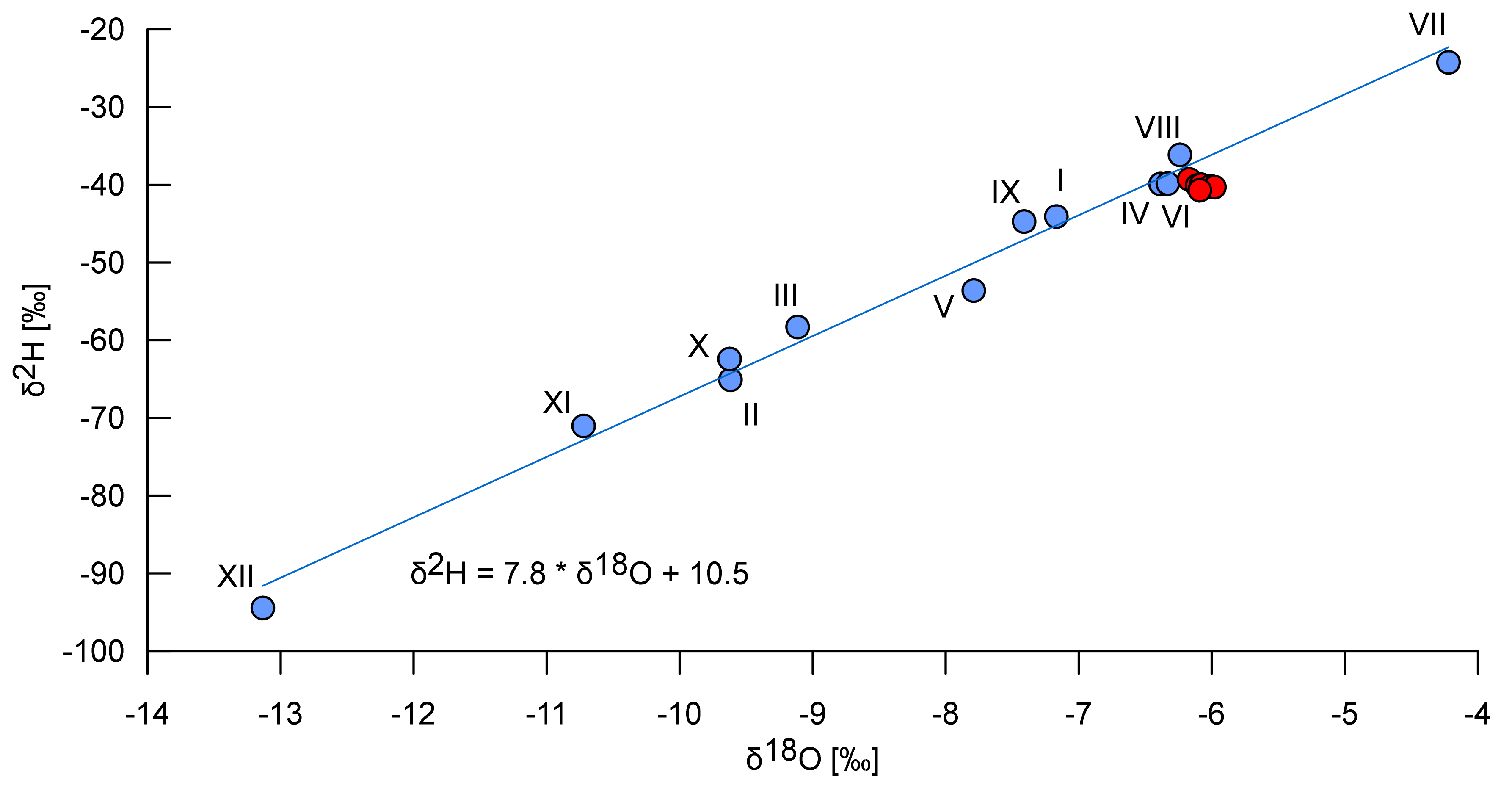
2.3. Physicochemical and Toxicity Analyses
2.4. Sampling and Microbiological Analyses
2.5. Amplicon Sequencing, and Bioinformatics Analysis
3. Results and Discussion
3.1. Environment of the Spring
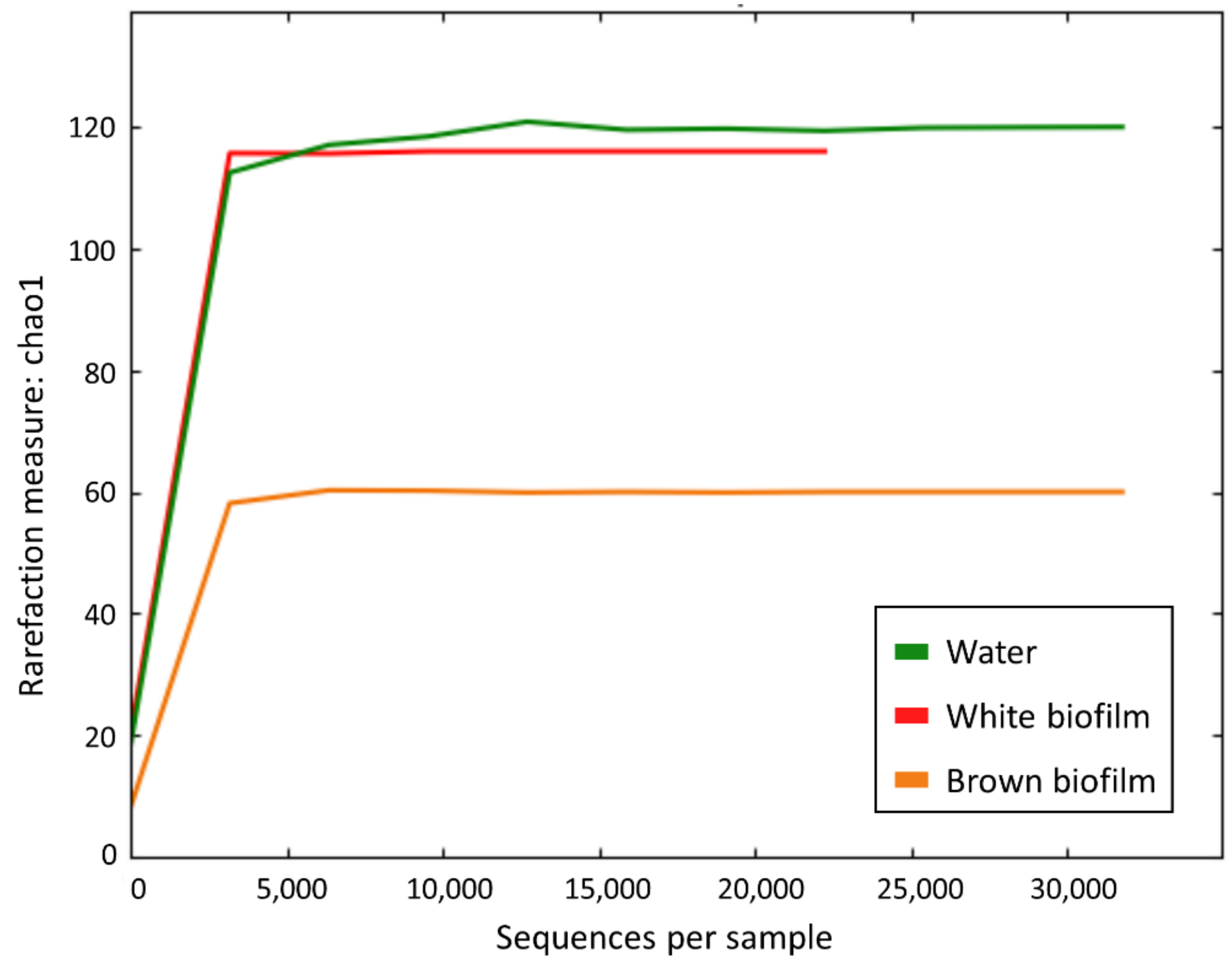
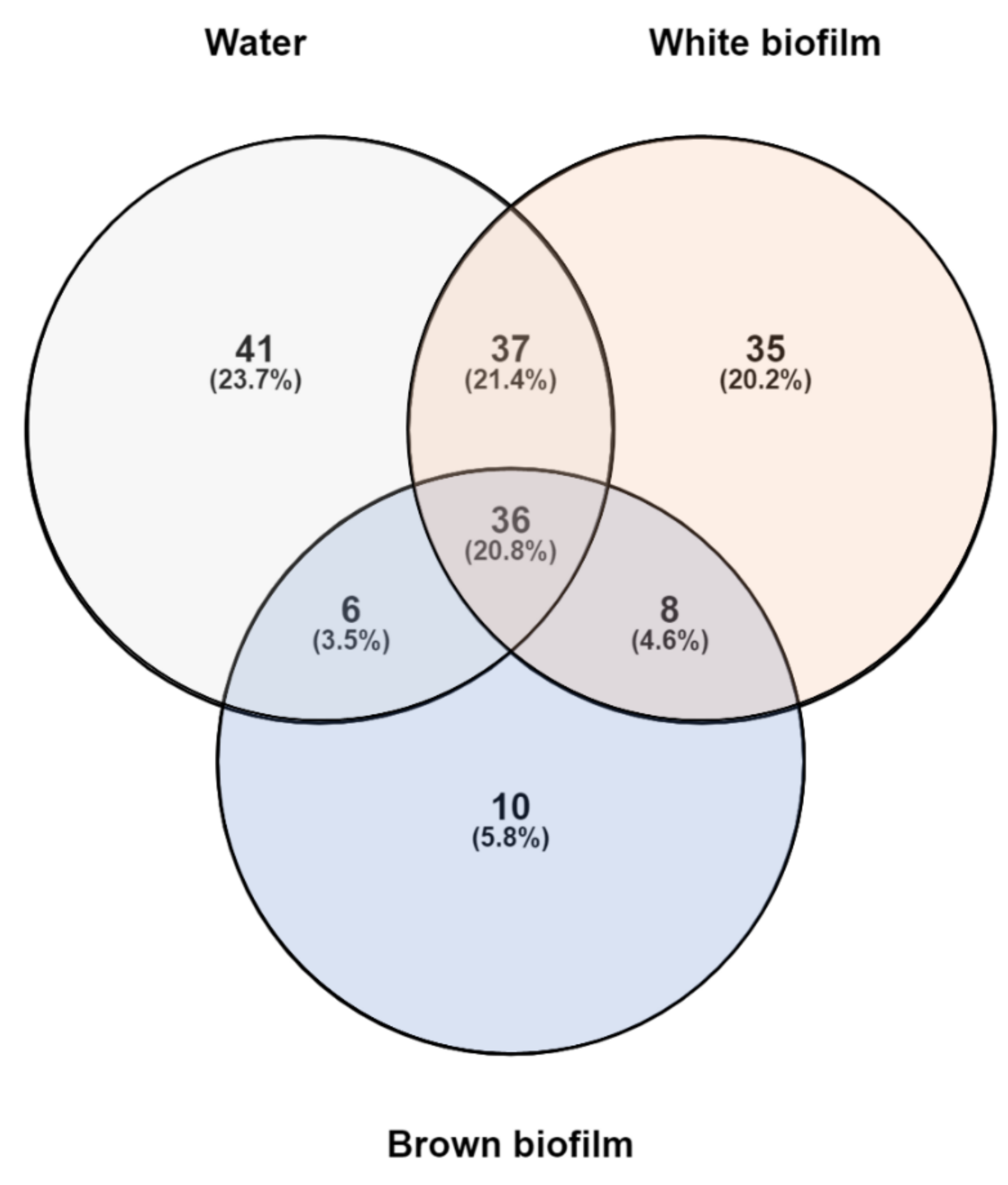
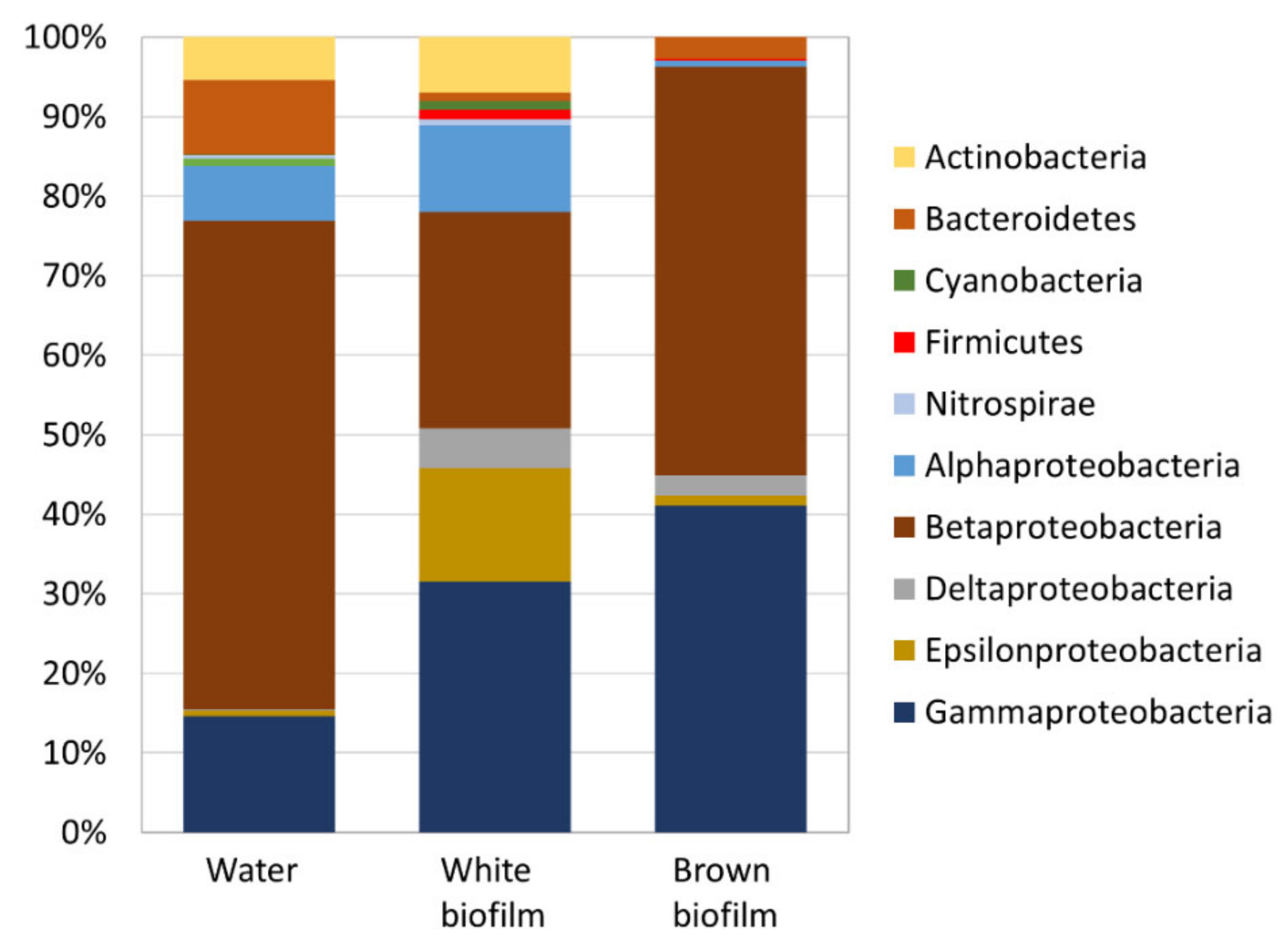
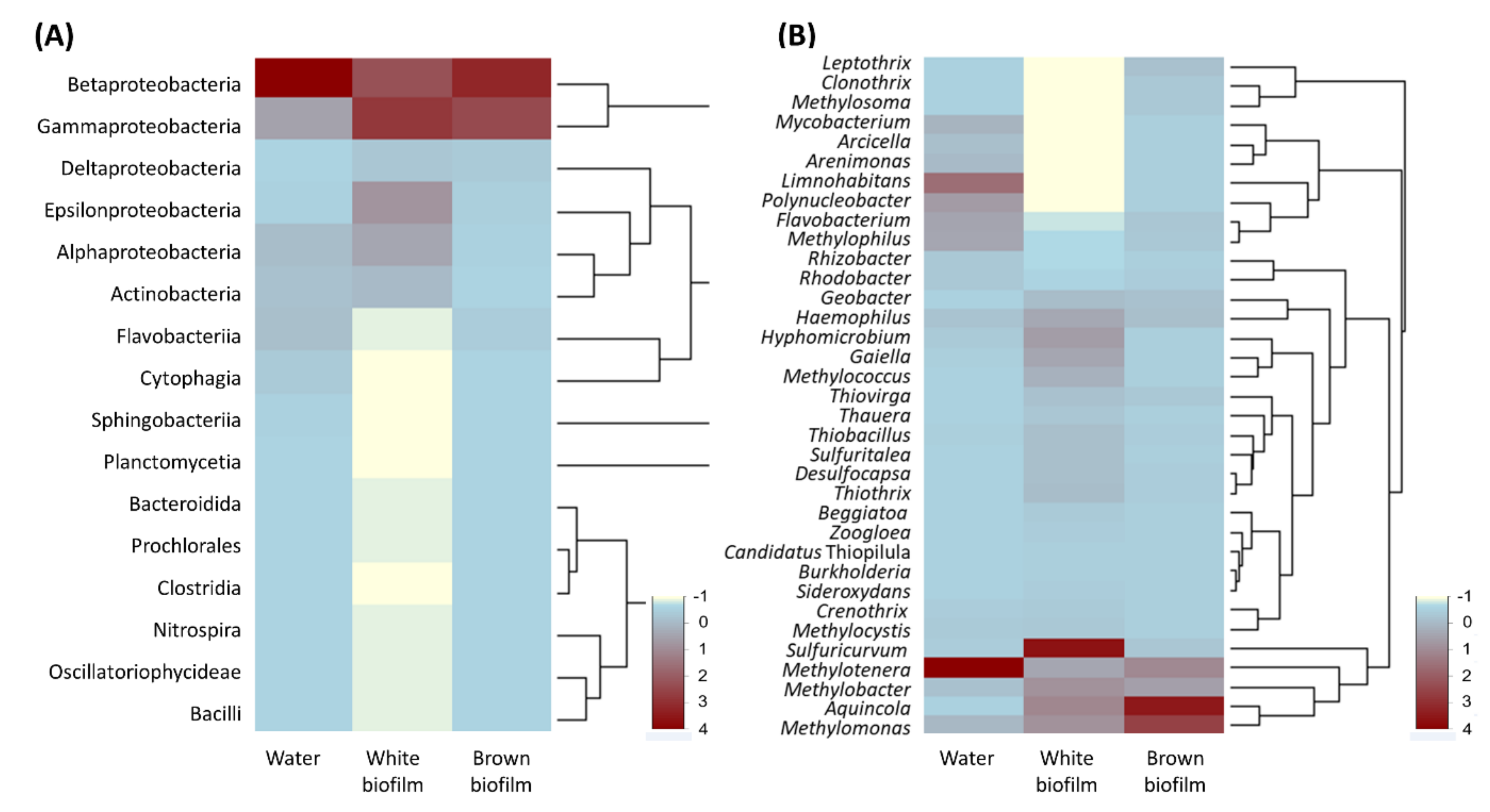
3.2. Microbial Diversity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qing, Y.; Wang, S.; Zhang, B.; Wang, Y. Ultra-high resolution regional climate projections for assessing changes in hydrological extremes and underlying uncertainties. Clim. Dynam. 2020, 55, 2031–2051. [Google Scholar] [CrossRef]
- Schippers, P.; Abarca, E.L.; Verboom, J.; Wamelink, G.W.W.; Vos, C.C.; de Boer, W.F.; Harvey, J.A.; Essens, T.; Grashof-Bokdam, C.J.; WallisDeVries, M.F.; et al. Biodiversity conservation in climate change driven transient communities. Biodivers. Conserv. 2021, 30, 2885–2906. [Google Scholar] [CrossRef]
- Hoffmann, S.; Irl, S.D.H.; Beierkuhnlein, C. Predicted climate shifts within terrestrial protected areas worldwide. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPCC. Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P., Skea, J., Calvo Buendia, E., Masson-Delmotte, V., Pörtner, H., Roberts, D., Zhai, P., Slade, R., Connors, S., van Diemen, R., et al., Eds.; IPCC: Geneva, Switzerland, 2019; p. 864, In press. [Google Scholar]
- Root, T.L.; Price, J.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef]
- Roman-Palacios, C.; Wiens, J.J. Recent responses to climate change reveal the drivers of species extinction and survival. Proc. Natl. Acad. Sci. USA 2020, 117, 4211–4217. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.G.; Calder, W.J.; Cumming, G.S.; Hughes, T.P.; Jentsch, A.; LaDeau, S.L.; Lenton, T.M.; Shuman, B.N.; Turetsky, M.R.; Ratajczak, Z.; et al. Climate change, ecosystems and abrupt change: Science priorities. Philos. Trans. R. Soc. B. 2020, 375, 20190105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renaud, P.E.; Wallhead, P.; Kotta, J.; Wtodarska-Kowalczuk, M.; Bellerby, R.G.J.; Ratsep, M.; Slagstad, D.; Kuklinski, P. Arctic sensitivity? Suitable habitat for benthic taxa is surprisingly robust to climate change. Front. Mar. Sci. 2019, 6, 538. [Google Scholar] [CrossRef]
- Mammola, S.; Piano, E.; Cardoso, P.; Vernon, P.; Dominguez-Villar, D.; Culver, D.C.; Pipan, T.; Isaia, M. Climate change going deep: The effects of global climatic alterations on cave ecosystems. Anthr. Rev. 2019, 6, 98–116. [Google Scholar] [CrossRef]
- Kovačič, G.; Petrič, M.; Ravbar, N. Evaluation and quantification of the effects of climate and vegetation cover change on karst water sources: Case studies of two springs in South-Western Slovenia. Water 2020, 12, 3087. [Google Scholar] [CrossRef]
- Retter, A.; Karwautz, C.; Griebler, C. Groundwater microbial communities in times of climate change. Curr. Issues. Mol. Biol. 2021, 41, 509–538. [Google Scholar] [CrossRef]
- Lai, G.G.; Padedda, B.M.; Ector, L.; Wetzel, C.E.; Luglie, A.; Cantonati, M. Mediterranean karst springs: Diatom biodiversity hotspots under the pressure of hydrological fluctuation and nutrient enrichment. Plant. Biosyst. 2020, 154, 673–684. [Google Scholar] [CrossRef]
- Cantonati, M.; Fureder, L.; Gerecke, R.; Juttner, I.; Cox, E.J. Crenic habitats, hotspots for freshwater biodiversity conservation: Toward an understanding of their ecology. Freshw. Sci. 2012, 31, 463–480. [Google Scholar] [CrossRef]
- Headd, B.; Engel, A.S. Biogeographic congruency among bacterial communities from terrestrial sulfidic springs. Front. Microbiol. 2014, 5, 473. [Google Scholar] [CrossRef] [Green Version]
- Mihevc, A. Speleogeneza Divaškega Krasa; Založba ZRC: Ljubljana, Slovenia, 2001; p. 180. [Google Scholar]
- Kranjc, A. Škocjanske jame. In Kras, Pokrajina, Življenje, Ljudje; Kranjc, A., Ed.; Založba ZRC: Ljubljana, Slovenia, 1999; pp. 77–82. [Google Scholar]
- Gams, I. Kras v Sloveniji v Prostoru in Času; Založba ZRC: Ljubljana, Slovenia, 2004; p. 515. [Google Scholar]
- Programme for Protection and Development of the Škocjan Caves Park for the 2019–2023 Period; The Škocjan Caves Park Public Agency: Škocjan, Slovenia, 2018.
- Jurkovšek, B.; Toman, M.; Ogorelec, B.; Šribar, L.; Drobne, K.; Poljak, M.; Šribar, L. Geological Map of the Southern Part of the Trieste-Komen Plateau, Cretaceous and Paleogene Carbonate Rocks 1:50,000; Inštitut za Geologijo, Geotehniko in Geofiziko: Ljubljana, Slovenia, 1996. [Google Scholar]
- Jurkovšek, B. Geološka Karta Krasa 1:100,000; Geološki Zavod Slovenije: Ljubljana, Slovenia, 2013.
- Šebela, S. Structural geology of the Škocjan Caves. Acta Carsol. 2009, 38, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Hamrla, M. O pogojih nastanka premogišč na Krasu. Geologija 1959, 5, 180–264. [Google Scholar]
- Hamrla, M. Light reflectance of some Slovenian coals. Geologija 1987, 28–29, 293–317. [Google Scholar]
- Geocaching.com. Opuščen Rudnik Črnega Premoga Vremski Britof. Available online: https://www.geocaching.com/geocache/GC8B52A_opuscen-rudnik-crnega-premoga-vremski-britof?guid=1d18a9da-4d74-4f70-b803-5adef69c815c (accessed on 10 September 2021).
- Malec, T. Rudnik v Vremski Dolini. Available online: http://www.tatjana-malec.si/default.asp?Id=420 (accessed on 10 September 2021).
- Poljak, M. Structural-Tectonic Map of Slovenia, Based on Basic Geological Map of SFR Yugoslavia 1:100000; Mladinska Knjiga: Ljubljana, Slovenia, 2000. [Google Scholar]
- Placer, L. Poenostavljena strukturno-geološka karta Krasa. Geologija 2015, 58, 89–93. [Google Scholar] [CrossRef]
- Žvab Rožič, P.; Čar, J.; Rožič, B. Geological structure of the Divača area and its influence on speleogenesis and hydrogeology of Kačna jama. Acta Carsol. 2015, 44, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Knez, M. Vpliv Lezik na Razvoj Kraških Jam: Primer Velike Doline, Škocjanske Jame; Založba ZRC: Ljubljana, Slovenia, 1996; p. 186. [Google Scholar]
- Knez, M. The influence of bedding-planes on the development of Karst caves (a study of Velika Dolina at Škocjanske Jame caves, Slovenia). Carbonate. Evaporite. 1998, 13, 121–131. [Google Scholar] [CrossRef]
- Gospodarič, R. Cave sediments and Škocjanske jame speleogenesis. Acta Carsol. 1984, 12, 27–48. [Google Scholar]
- Slovenian Environment Agency (ARSO). Lidar Data. Available online: http://gis.arso.gov.si/evode/profile.aspx?id=atlas_voda_Lidar@Arso&culture=en-US (accessed on 10 September 2021).
- Clesceri, L.S.; Greenberg, A.E.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 1998. [Google Scholar]
- Mulec, J.; Oarga-Mulec, A.; Holko, L.; Pašić, L.; Kopitar, A.N.; Eleršek, T.; Mihevc, A. Microbiota entrapped in recently-formed ice: Paradana Ice Cave, Slovenia. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Holko, L. Syringe life and memory effects in isotopic analyses performed by liquid water isotopic analysers—A case study for natural waters from central Europe. Isot. Environ. Health Stud. 2016, 52, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Coplen, T.B.; Wassenaar, L.I. LIMS for Lasers 2015 for achieving long-term accuracy and precision of δ2 H, δ17 O, and δ18 O of waters using laser absorption spectrometry. Rapid Commun. Mass Spectom. 2015, 29, 2122–2130. [Google Scholar] [CrossRef] [PubMed]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 1. Teil: Chroococcales; Spectrum Akademischer: Heidelberg/Berlin, Germany, 2000; p. 548. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Cyanoprokaryota 2. Teil: Oscillatoriales; Elsevier, Spektrum Akademischer: München, Germany, 2005; p. 759. [Google Scholar]
- Komárek, J. Cyanoprokaryota 3. Teil: Heterocytous genera; Springer Spektrum: Berlin, Heidelberg, 2013; p. 1130. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 1. Teil: Naviculaceae; Gustav Fischer: Stuttgart, Germany; New York, NY, USA, 1986; p. 876. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae; Gustav Fischer: Stuttgart, Germany; New York, NY, USA, 1988; p. 596. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 3. Teil: Centrales, Fragilariaceae, Eunotiaceae; Gustav Fischer: Stuttgart, Germany; New York, NY, USA, 1991; p. 576. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Achnanthes s.l., Navicula s. str., Gomphonema; Gustav Fischer: Stuttgart, Germany; New York, NY, USA, 1991; p. 468. [Google Scholar]
- Caporaso, J.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.; Costello, E.; Fierer, N.; Pena, A.; Goodrich, J.; Gordon, J.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’hara, R.; Simpson, G.; Solymos, P.; Stevens, M.; Wagner, H. Vegan: Community ecology package. R Package Version 2019, 2, 5–6. [Google Scholar]
- Huber, W.; Carey, V.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.; Bravo, H.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Team RC. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- OECD. Test No. 202: Daphnia sp. Acute Immobilisation Test., OECD Guidelines for the Testing of Chemicals, Section 2; OECD Publishing: Paris, France, 2004. [Google Scholar]
- Foudhaili, T.; Jaidi, R.; Neculita, C.M.; Rosa, E.; Triffault-Bouchet, G.; Veilleux, E.; Coudert, L.; Lefebvre, O. Effect of the electrocoagulation process on the toxicity of gold mine effluents: A comparative assessment of Daphnia magna and Daphnia pulex. Sci. Total. Environ. 2020, 708, 134739. [Google Scholar] [CrossRef]
- Medunić, G.; Fiket, Ž.; Ivanić, M. Arsenic contamination status in Europe, Australia, and other parts of the world. In Arsenic in Drinking Water and Food; Srivastava, S., Ed.; Springer: Singapore, 2020; pp. 183–233. [Google Scholar]
- Van Elsas, J.D.; Semenov, A.V.; Costa, R.; Trevors, J.T. Survival of Escherichia coli in the environment: Fundamental and public health aspects. ISME J. 2011, 5, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.; Hur, H.G.; Sadowsky, M.J.; Byappanahalli, M.N.; Yan, T.; Ishii, S. Environmental Escherichia coli: Ecology and public health implications—A review. J. Appl. Microbiol. 2017, 123, 570–581. [Google Scholar] [CrossRef] [Green Version]
- Zhi, S.; Banting, G.; Stothard, P.; Ashbolt, N.J.; Checkley, S.; Meyer, K.; Otto, S.; Neumann, N.F. Evidence for the evolution, clonal expansion and global dissemination of water treatment-resistant naturalized strains of Escherichia coli in wastewater. Water Res. 2019, 156, 208–222. [Google Scholar] [CrossRef]
- Ingle, D.J.; Clermont, O.; Skurnik, D.; Denamur, E.; Walk, S.T.; Gordon, D.M. Biofilm formation by and thermal niche and virulence characteristics of Escherichia spp. Appl. Environ. Microb. 2011, 77, 2695–2700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vital, M.; Hammes, F.; Egli, T. Escherichia coli O157 can grow in natural freshwater at low carbon concentrations. Environ. Microbiol. 2008, 10, 2387–2396. [Google Scholar] [CrossRef]
- Guo, H.; Liu, R.; Yu, Z.; Zhang, H.; Yun, J.; Li, Y.; Liu, X.; Pan, J. Pyrosequencing reveals the dominance of methylotrophic methanogenesis in a coal bed methane reservoir associated with Eastern Ordos Basin in China. Int. J. Coal Geol. 2012, 93, 56–61. [Google Scholar] [CrossRef]
- Wolinska, A.; Pytlak, A.; Stepniewska, Z.; Kuzniar, A.; Piasecki, C. Identification of methanotrophic bacteria community in the Jastrzebie-Moszczenica coal mine by fluorescence in situ hybridization and PCR techniques. Pol. J. Environ. Stud. 2013, 22, 275–282. [Google Scholar]
- Bowman, J. Methylomonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W., Rainey, F., Kampfer, P., Trujillo, M., Chun, J., DeVos, P., Hedlund, B., Dedysh, S., Eds.; Wiley Online Library: Hoboken, NJ, USA, 2015; pp. 1–10. [Google Scholar]
- Giri, D.D.; Kumar, A.; Shukla, P.N.; Singh, R.; Singh, P.K.; Pandey, K.D. Salt stress tolerance of methylotrophic bacteria Methylophilus sp. and Methylobacterium sp. isolated from coal mine spoils. Pol. J. Environ. Stud. 2013, 62, 273–280. [Google Scholar] [CrossRef]
- Wang, B.; Wang, Y.; Cui, X.; Zhang, Y.; Yu, Z. Bioconversion of coal to methane by microbial communities from soil and from an opencast mine in the Xilingol grassland of northeast China. Biotechnol. Biofuels 2019, 12, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadnikov, V.V.; Mardanov, A.V.; Beletsky, A.V.; Antsiferov, D.V.; Kovalyova, A.A.; Karnachuk, O.V.; Ravin, N.V. Sulfur-oxidizing bacteria dominate in the water from a flooded coal mine shaft in Kuzbass. Microbiology 2019, 88, 120–123. [Google Scholar] [CrossRef]
- Van Grinsven, S.; Damste, J.S.S.; Harrison, J.; Polerecky, L.; Villanueva, L. Nitrate promotes the transfer of methane-derived carbon from the methanotroph Methylobacter sp. to the methylotroph Methylotenera sp. in eutrophic lake water. Limnol. Oceanogr. 2021, 66, 878–891. [Google Scholar] [CrossRef]
- Hahn, M.W.; Kasalicky, V.; Jezbera, J.; Brandt, U.; Jezberova, J.; Simek, K. Limnohabitans curvus gen. nov., sp nov., a planktonic bacterium isolated from a freshwater lake. Int. J. Syst. Evol. Micr. 2010, 60, 1358–1365. [Google Scholar] [CrossRef]
- Han, B.; Chen, Y.; Abell, G.; Jiang, H.; Bodrossy, L.; Zhao, J.; Murrell, J.C.; Xing, X.H. Diversity and activity of methanotrophs in alkaline soil from a Chinese coal mine. Fems. Microbiol. Ecol. 2009, 70, 196–207. [Google Scholar] [CrossRef] [Green Version]
- Nuy, J.K.; Hoetzinger, M.; Hahn, M.W.; Beisser, D.; Boenigk, J. Ecological differentiation in two major freshwater bacterial taxa along environmental gradients. Front. Microbiol. 2020, 11, 154. [Google Scholar] [CrossRef]
- Boenigk, J.; Stadler, P.; Wiedlroither, A.; Hahn, M.W. Strain-specific differences in the grazing sensitivities of closely related ultramicrobacteria affiliated with the Polynucleobacter cluster. Appl. Environ. Microb. 2004, 70, 5787–5793. [Google Scholar] [CrossRef] [Green Version]
- Millar, W. Heterotrophic bacterial population in acid coal mine water: Flavobacterium acidurans sp. n. Int. J. Syst. Bacteriol. 1973, 23, 142–150. [Google Scholar] [CrossRef] [Green Version]
- Kodama, Y.; Watanabe, K. Sulfuricurvum kujiense gen. nov., sp nov., a facultatively anaerobic, chemolithoautotrophic, sulfur-oxidizing bacterium isolated from an underground crude-oil storage cavity. Int. J. Syst. Evol. Micr. 2004, 54, 2297–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macalady, J.L.; Dattagupta, S.; Schaperdoth, I.; Jones, D.S.; Druschel, G.K.; Eastman, D. Niche differentiation among sulfur-oxidizing bacterial populations in cave waters. ISME J. 2008, 2, 590–601. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, T.L.; Jones, D.S.; Schaperdoth, I.; Macalady, J.L. Metagenomic insights into S(0) precipitation in a terrestrial subsurface lithoautotrophic ecosystem. Front. Microbiol. 2015, 5, 756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossmassler, K.; Hanson, T.E.; Campbell, B.J. Diverse sulfur metabolisms from two subterranean sulfidic spring systems. FEMS Microbiol. Lett. 2016, 363, fnw162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reigstad, L.J.; Jorgensen, S.L.; Lauritzen, S.E.; Schleper, C.; Urich, T. Sulfur-oxidizing chemolithotrophic proteobacteria dominate the microbiota in high Arctic thermal springs on Svalbard. Astrobiology 2011, 11, 665–678. [Google Scholar] [CrossRef]
- Lechner, U.; Brodkorb, D.; Geyer, R.; Hause, G.; Hartig, C.; Auling, G.; Fayolle-Guichard, F.; Piveteau, P.; Muller, R.H.; Rohwerder, T. Aquincola tertiaricarbonis gen. nov., sp. nov., a tertiary butyl moiety-degrading bacterium. Int. J. Syst. Evol. Micr. 2007, 57, 1295–1303. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Xia, M.; Zhang, L.; Song, Y.; Duan, Y.; Yuan, T.; Yao, M.; Wu, L.; Tian, C.; Wu, Z.; et al. RpoN (σ54) is required for floc formation but not for extracellular polysaccharide biosynthesis in a floc-forming Aquincola tertiaricarbonis strain. Appl. Environ. Microb. 2017, 83, e00709-17. [Google Scholar] [CrossRef] [Green Version]
- Eleršek, T.; Mulec, J. The algal community at an ecocline of a cold sulphidic spring (Sovra artesian borehole, Slovenia). Environ. Earth. Sci. 2014, 71, 5255–5261. [Google Scholar] [CrossRef]
- AlgaeBase. Available online: https://www.algaebase.org/ (accessed on 9 October 2021).
- Leira, M.; Meijide-Failde, R.; Torres, E. Diatom communities in thermo-mineral springs of Galicia (NW Spain). Diatom. Res. 2017, 32, 29–42. [Google Scholar] [CrossRef]
- Luis, A.T.; Teixeira, P.; Almeida, S.F.P.; Ector, L.; Matos, J.X.; Ferreira da Silva, E.A. Impact of acid mine drainage (AMD) on water quality, stream sediments and periphytic diatom communities in the surrounding streams of Aljustrel mining area (Portugal). Water Air Soil Poll. 2009, 200, 147–167. [Google Scholar] [CrossRef]
- Yanagawa, K.; Haraguchi, A.; Yoshitake, K.; Asamatsu, K.; Harano, M.; Yamashita, K.; Ishibashi, J. Ubiquity of Euglena mutabilis population in three ecologically distinct acidic habitats in Southwestern Japan. Water 2021, 13, 1570. [Google Scholar] [CrossRef]
- Danilova, E.; Barkhutova, D.; Bryanskaya, A.; Namsaraev, Z.; Namsaraev, B. Effect of environmental conditions on the distribution of functional groups of microorganisms in the Khoito-Gol mineral springs (East Sayan). Contemp. Probl. Ecol. 2009, 2, 33–39. [Google Scholar] [CrossRef]
- Lumian, J.E.; Jungblut, A.D.; Dillion, M.L.; Hawes, I.; Doran, P.T.; Mackey, T.J.; Dick, G.J.; Grettenberger, C.L.; Sumner, D.Y. Metabolic capacity of the Antarctic cyanobacterium Phormidium pseudopriestleyi that sustains oxygenic photosynthesis in the presence of hydrogen sulfide. Genes 2021, 12, 426. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Zheng, Y. Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef] [PubMed]
- Barranguet, C.; Veuger, B.; Van Beusekom, S.A.M.; Marvan, P.; Sinke, J.J.; Admiraal, W. Divergent composition of algal-bacterial biofilms developing under various external factors. Eur. J. Phycol. 2005, 40, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mulec, J.; Oarga, A.; Schiller, E.; Persoiu, A.; Holko, L.; Sebela, S. Assessment of the physical environment of epigean invertebrates in a unique habitat: The case of a karst sulfidic spring, Slovenia. Ecohydrology 2015, 8, 1326–1334. [Google Scholar] [CrossRef]
- Mulec, J.; Summers Engel, A. Karst spring microbial diversity differs across an oxygen-sulphide ecocline and reveals potential for novel taxa discovery. Acta Carsol. 2019, 48, 129–143. [Google Scholar] [CrossRef] [Green Version]
- Kark, S. Ecotones and ecological gradients. In Ecological Systems: Selected Entries from the Encyclopedia of Sustainability Science and Technology; Leemans, R., Ed.; Springer: New York, NY, USA, 2013; pp. 147–160. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Body | Date | d18O (‰) | d2H (‰) | Deuterium Excess |
|---|---|---|---|---|
| Reka | 18 July 2017 | −6.17 | −39.3 | 10.1 |
| Smrdljivec | −6.18 | −39.3 | 10.1 | |
| Reka | 30 July 2017 | −6.11 | −40.1 | 8.8 |
| Smrdljivec | −6.08 | −39.8 | 8.8 | |
| Reka | 1 August 2017 | −6.08 | −40 | 8.6 |
| Smrdljivec | −6.09 | −39.9 | 8.8 | |
| Reka | 9 August 2017 | −6.01 | −40.2 | 7.9 |
| Smrdljivec | −6.03 | −40.4 | 7.8 | |
| Reka | 24 August 2017 | −5.98 | −40.3 | 7.5 |
| Smrdljivec | −6.02 | −40.6 | 7.6 | |
| Reka | 31 August 2017 | −6.09 | −40.7 | 8.0 |
| Smrdljivec | −6.09 | −40.9 | 7.8 |
| Water Body Date | |||||
|---|---|---|---|---|---|
| Parameter | Smrdljivec 18 July 2017 | Reka 18 July 2017 | Smrdljivec 24 August 2017 | Smrdljivec 28 July 2021 | Reka 28 July 2021 |
| Temperature (°C) | 21.5 | 21.8 | 19.7 | 21.2 | 21.4 |
| pH (≥6.5 and ≤9.5) | 7.28 | 8.00 | 7.28 | 7.57 | 8.26 |
| EC (μS/cm) (2500 μS/cm) | 371 | 363 | 370 | 334 | 330 |
| O2 (mg/L) | 0.17 | 8.91 | 0.39 | 0.59 | 8.74 |
| O2 (%) | 1.8 | 105.0 | 4.4 | 6.9 | 102.8 |
| Hardness (CaCO3 mg/L) | 183 | 187 | nd | 160 | 130 |
| Alkalinity (CaCO3 mg/L) | 189 | 184 | nd | 165 | 150 |
| Ca (mg/L) | 66.5 | 67.3 | 99.0 | 61.3 | 58.0 |
| Cl (mg/L) (250 mg/L) | 6.2 | 6.2 | 10.0 | 4.7 | 4.7 |
| Fe (mg/L) (200 µg/L) | 1.7 | 0.1 | 11.0 | 0.1 | 0.0 |
| H2S (mg/L) | Nd | 0.0 | 0.15 | 0.23 | 0.0 |
| K (mg/L) | 2.1 | 1.7 | 1.6 | 1.3 | 1.3 |
| Mg (mg/L) | 4.0 | 4.5 | 5.0 | 4.3 | 4.2 |
| NH4 (mg/L) | 0.4 | <0.2 | 0.2 | <0.2 | <0.2 |
| NO2 (mg/L) (0.50 mg/L) | <0.01 | 0.01 | <0.01 | <0.01 | 0.08 |
| NO3 (mg/L) (50 mg/L) | 0.4 | 0.5 | <2.2 | <0.1 | 1.5 |
| SiO2 (mg/L) | 3.4 | 1.8 | 3.4 | 5.1 | 2.8 |
| SO4 (mg/L) (250 mg/L) | 2.4 | 4.3 | 2.4 | 6.6 | 7.7 |
| PO4 (mg/L) | 0.16 | <0.05 | nd | <0.05 | <0.05 |
| Al (µg/L) (200 µg/L) | 4100 | ||||
| Cu (µg/L) (2000 µg/L) | 10 | ||||
| Cr (µg/L) (50 µg/L) | 5.7 | ||||
| Mn (µg/L) (50 µg/L) | 660 | ||||
| Ni (µg/L) (20 µg/L) | 22 | ||||
| Pb (µg/L) (10 µg/L) | 9.9 | ||||
| Zn (µg/L) | 38 | ||||
| TOC (mg/L) | 1.46 | ||||
| D. magna toxicity test (% immobilization) | 10 | ||||
| ATP (RLU) | 85 | 1160 | 217 | 157 | 757 |
| Bacteria 22 °C (CFU/mL) (100 CFU/mL) | 8 | 333 | nd | 87 | 287 |
| Bacteria 37 °C (CFU/mL) (20 CFU/mL) | 11 | 263 | 28 | 68 | 349 |
| E. coli (CFU/mL) (0 CFU/250 mL) | 0 | 0 | 0 | 0.05 * | 0.17 * |
| Coliforms (CFU/mL) (0 CFU/100 mL) | 3 | 208 | 8 | 10 | 208 |
| Enterococci (CFU/mL) (0/250 mL) | 1 | 0 | 0 | 2.56 * | 2.30 * |
| Diversity | ||||
|---|---|---|---|---|
| Sample | Number of Reads | Total OTUs | ACE | Shannon |
| Water | 143,271 | 120 | 120.0 ± 2.4 | 1.71 |
| White biofilm | 83,156 | 116 | 116.0 ± 1.7 | 2.98 |
| Brown biofilm | 98,578 | 60 | 60.0 ± 2.3 | 1.91 |
| Taxon | Class | Biofilm Type: Date |
|---|---|---|
| Achnanthidium sp. | Bacillariophyceae | B: 24 August 2017, W: 17 July 2017 |
| Closterium sp. | Zygnematophyceae | B: 28 July 2021 |
| Cocconeis sp. | Bacillariophyceae | B: 28 July 2021 |
| Coelastrum sp. | Chlorophyceae | B: 28 July 2021 |
| Cosmarium sp. | Zygnematophyceae | B: 28 July 2021 |
| Craticula cuspidata (Navicula cuspidata) | Bacillariophyceae | B: 17 July 2017, B: 24 August 2017, W: 17 July 2017 |
| Cymatopleura elliptica | Bacillariophyceae | W: 17 July 2017 |
| Cymatopleura solea | Bacillariophyceae | W: 17 July 2017 |
| Dinobryon divergens | Chrysophyceae | B: 24 August 2017 |
| Euglena sp. | Euglenoidea | B: 28 July 2021 |
| Gyrosigma sp. | Bacillariophyceae | B: 28 July 2021 |
| Melosira varians | Bacillariophyceae | W: 17 July 2017, W: 24 August 2017 |
| Merismopedia sp. | Cyanophyceae | B: 28 July 2021 |
| Navicula sp. | Bacillariophyceae | B: 28 July 2021, W: 17 July 2017 |
| Nitzschia linearis | Bacillariophyceae | B: 28 July 2021 |
| Nitzschia palea | Bacillariophyceae | B: 28 July 2021 |
| Nitzschia sp. | Bacillariophyceae | B: 24 August 2017, B: 28 July 2021, W: 24 August 2017, W: 17 July 2017 |
| Oscillatoria limosa | Cyanophyceae | B: 24 August 2017, W: 24 August 2017 |
| Pediastrum duplex | Chlorophyceae | B: 24 August 2017 |
| Phacus sp. (Lepocinclis sp.) | Euglenoidea | B: 28 July 2021 |
| Phormidium sp. | Cyanophyceae | B: 24 August 2017, B: 28 July 2021, W: 24 August 2017 |
| Rhizosolenia longiseta | Bacillariophyceae | W: 24 August 2017 |
| Scenedesmus opoliensis | Chlorophyceae | B: 24 August 2017, B: 28 July 2021 |
| Selephora pupula (Navicula pupula) | Bacillariophyceae | B: 24 August 2017 |
| Spirogyra sp. | Zygnematophyceae | B: 28 July 2021 |
| Surirella sp. | Bacillariophyceae | B: 28 July 2021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulec, J.; Oarga-Mulec, A.; Skok, S.; Šebela, S.; Cerkvenik, R.; Zorman, T.; Holko, L.; Eleršek, T.; Pašić, L. Emerging Ecotone and Microbial Community of a Sulfidic Spring in the Reka River near Škocjanske Jame, Slovenia. Diversity 2021, 13, 655. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120655
Mulec J, Oarga-Mulec A, Skok S, Šebela S, Cerkvenik R, Zorman T, Holko L, Eleršek T, Pašić L. Emerging Ecotone and Microbial Community of a Sulfidic Spring in the Reka River near Škocjanske Jame, Slovenia. Diversity. 2021; 13(12):655. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120655
Chicago/Turabian StyleMulec, Janez, Andreea Oarga-Mulec, Sara Skok, Stanka Šebela, Rosana Cerkvenik, Tomaž Zorman, Ladislav Holko, Tina Eleršek, and Lejla Pašić. 2021. "Emerging Ecotone and Microbial Community of a Sulfidic Spring in the Reka River near Škocjanske Jame, Slovenia" Diversity 13, no. 12: 655. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120655