
Animal Forest Mortality: Following the Consequences of a Gorgonian Coral Loss on a Mediterranean Coralligenous Assemblage

 , , , , and
, , , , and
Abstract
:1. Introduction
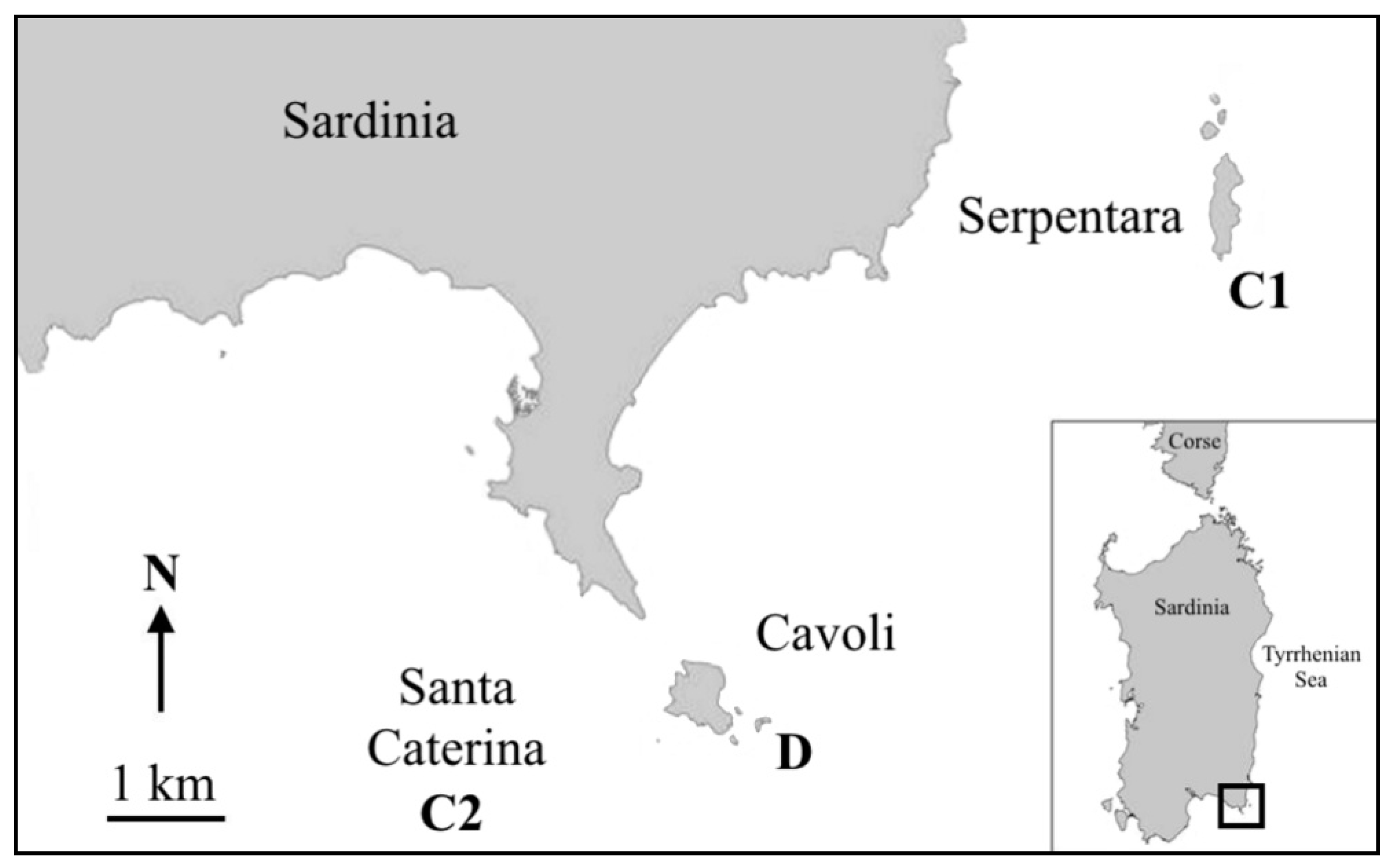
2. Material and Methods
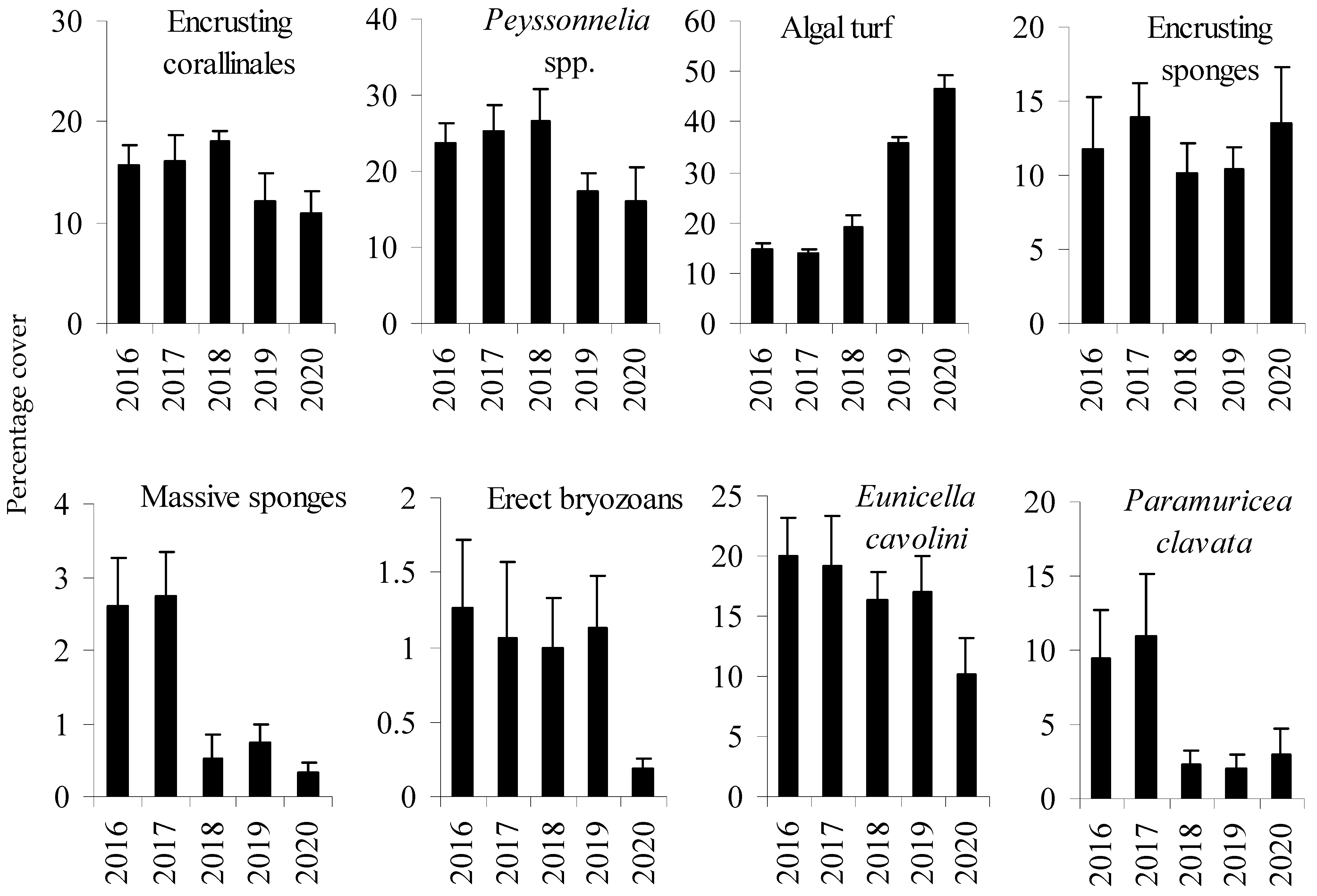
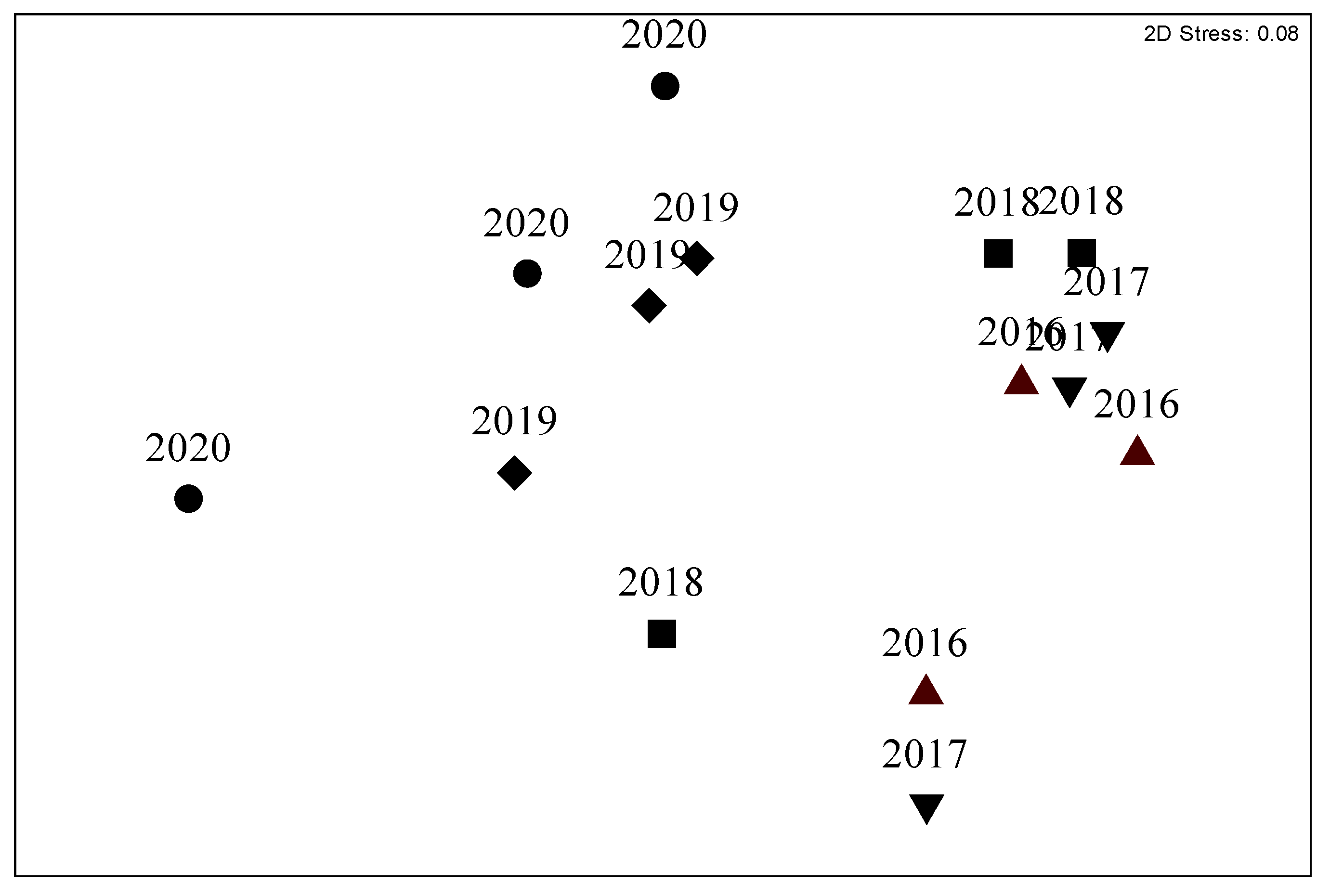
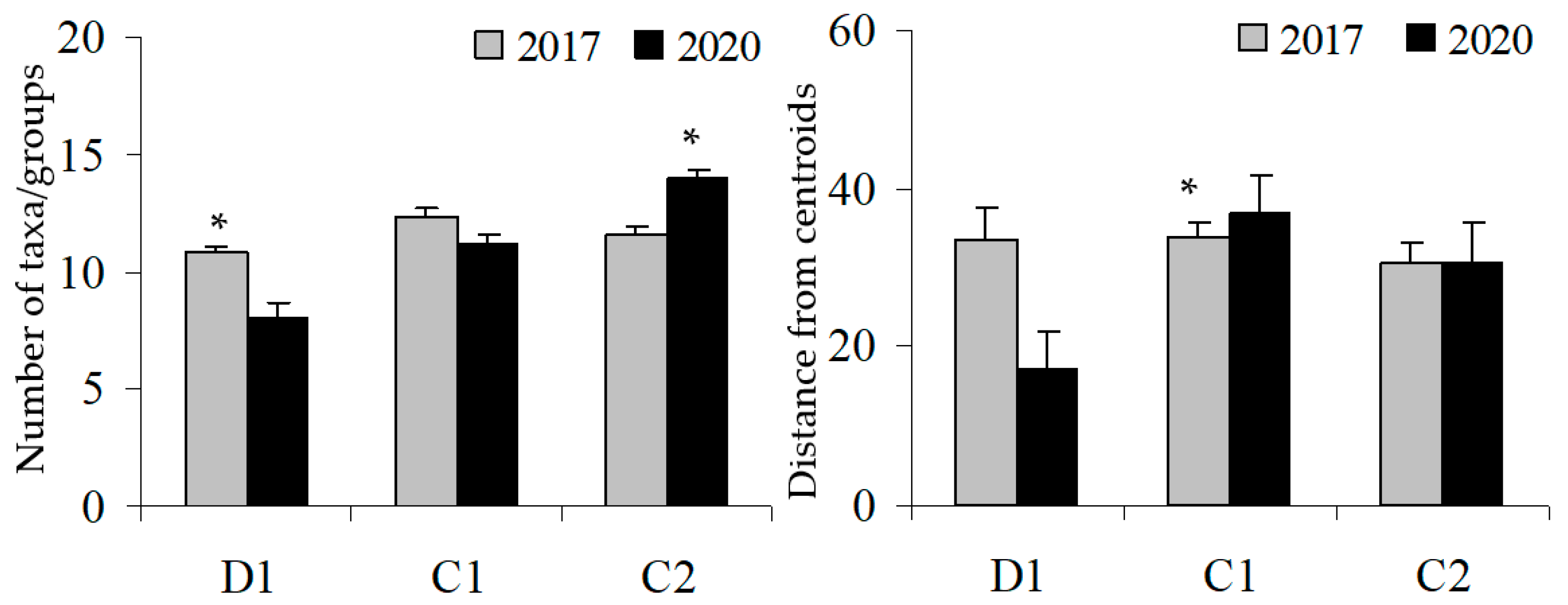
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crisci, C.; Bensoussan, N.; Romano, J.C.; Garrabou, J. Temperature anomalies and mortality events in marine communities: Insights on factors behind differential mortality impacts in the NW Mediterranean. PLoS ONE 2011, 6, e23814. [Google Scholar] [CrossRef] [Green Version]
- Eakin, M.; Liu, G.; Gomez, A.; De la Cour, J.; Heron, S.; Skirving, W. Global coral bleaching 2014–2017: Status and an appeal for observations. Reef Encount. 2016, 31, 20–26. [Google Scholar]
- Ereskovsky, A.; Ozerov, D.A.; Pantyulin, A.N.; Tzetlin, A.B. Mass mortality event of White Sea sponges as the result of high temperature in summer 2018. Polar Biol. 2019, 42, 2313–2318. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Sean, R.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Jones, N.P.; Figueiredo, J.; Gilliam, D.S. Thermal stress-related spatiotemporal variations in high-latitude coral reef benthic communities. Coral Reefs 2020, 39, 1661–1673. [Google Scholar] [CrossRef]
- Morri, C.; Montefalcone, M.; Lasagna, R.; Gatti, G.; Rovere, A.; Parravicini, V.; Baldelli, G.; Colantoni, P.; Bianchi, C.N. Through bleaching and tsunami: Coral reef recovery in the Maldives. Mar. Poll. Bull. 2015, 98, 188–200. [Google Scholar] [CrossRef]
- Montefalcone, M.; Morri, C.; Bianchi, C.N. Influence of local pressures on Maldivian coral reef resilience following repeated bleaching events, and recovery perspectives. Front. Mar. Sci. 2020, 7, 587. [Google Scholar] [CrossRef]
- Robinson, J.P.W.; Wilson, S.K.; Graham, N.A.J. Abiotic and biotic controls on coral recovery 16 years after mass bleaching. Coral Reefs 2019, 38, 1255–1265. [Google Scholar] [CrossRef] [Green Version]
- Vercelloni, J.; Mengersen, K.; Ruggeri, F.; Caley, M.J. Improved coral population estimation reveals trends at multiple scales on Australia’s Great Barrier Reef. Ecosystems 2017, 20, 1337–1350. [Google Scholar] [CrossRef]
- Bavestrello, G.; Boero, F. Necrosi e rigenerazione in Eunicella cavolinii (Anthozoa, Cnidaria) in Mar Ligure. Bollet. Mus. Ist. Biol. Univ. Genova 1986, 52, 295–300. [Google Scholar]
- Gaino, E.; Pronzato, R. Ultrastructural evidence of bacterial damage to Spongia officinalis fibres (Porifera, Demospongiae). Diseases Aquat. Organ. 1989, 6, 67–74. [Google Scholar] [CrossRef]
- Voultsiadou, E.; Dailianis, T.; Antoniadou, C.; Vafidis, D.; Dounas, C.; Chintiroglou, C.C. Aegean bath sponges: Historical data and current status. Rev. Fish. Sci. 2011, 19, 34–51. [Google Scholar] [CrossRef]
- Cerrano, C.; Bavestrello, G.; Bianchi, C.N.; Cattaneo-Vietti, R.; Bava, S.; Morganti, C.; Morri, C.; Pico, P.; Sarà, G.; Schiaparelli, S.; et al. A catastrophic mass-mortality episode of gorgonians and other organisms in the Ligurian Sea (Northwestern Mediterranean), summer 1999. Ecol. Lett. 2000, 3, 284–293. [Google Scholar] [CrossRef]
- Perez, T.; Garrabou, J.; Sartoretto, S.; Harmelin, J.G.; Francour, P.; Vacelet, J. Mass mortality of marine invertebrates: An unprecedent event in the North Occidental Mediterranean. CR Acad. Sci. Paris 2000, 323, 853–865. [Google Scholar] [CrossRef]
- Garrabou, J.; Coma, R.; Bensoussan, N.; Bally, M.; Chevaldonné, P.; Cigliano, M.; Diaz, D.; Harmelin, J.G.; Gambi, M.C.; Kersting, D.K.; et al. Mass mortality in Northwestern Mediterranean rocky benthic communities: Effects of the 2003 heat wave. Glob. Chang. Biol. 2009, 15, 1090–1103. [Google Scholar] [CrossRef]
- Rivetti, I.; Fraschetti, S.; Lionello, P.; Zambianchi, E.; Boero, F. Global warming and mass mortalities of benthic invertebrates in the Mediterranean Sea. PLoS ONE 2014, 9, e115655. [Google Scholar] [CrossRef] [Green Version]
- Marbà, N.; Gabriel, J.; Agusti, S.; Girard, C.; Duarte, C.M. Footprints of climate change on Mediterranean Sea biota. Front. Mar. Sci. 2015, 2, 56. [Google Scholar] [CrossRef]
- Kersting, D.K.; Bensoussan, N.; Linares, C. Long-term responses of the endemic reef-builder Cladocora caespitosa to Mediterranean warming. PLoS ONE 2013, 8, e70820. [Google Scholar] [CrossRef] [PubMed]
- Turicchia, E.; Abbiati, M.; Sweet, M.; Ponti, M. Mass mortality hits gorgonian forests at Montecristo Island. Dis. Aquat. Org. 2018, 131, 79–85. [Google Scholar] [CrossRef]
- Bensoussan, N.; Chiggiato, J.; Buongiorno Nardelli, B.; Pisano, A.; Garrabou, J. Insights on 2017 Marine Heat Waves in the Mediterranean Sea. In Copernicus Marine Service Ocean State, Report 3. J. Operat. Oceanogr. 2019. [Google Scholar] [CrossRef]
- Garrabou, J.; Gómez-Gras, D.; Ledoux, J.-B.; Linares, C.; Bensoussan, N.; López-Sendino, P.; Bazairi, H.; Espinosa, F.; Ramdani, M.; Grimes, S.; et al. Collaborative database to track mass mortality events in the Mediterranean Sea. Front. Mar. Sci. 2019, 6, 707. [Google Scholar] [CrossRef] [Green Version]
- Özalp, H.B.; Kersting, D.K. A pan-Mediterranean extinction? Pinna nobilis mass mortality has reached the Turkish straits system. Mar. Biodivers. 2020, 50, 81. [Google Scholar]
- Kruzic, P.; Rodic, P. Impact of climate changes on coralligenous community in the Adriatic Sea. In Proceedings of the Second Mediterranean Symposium of Coralligenous and Other Calcareous Bioconcretions, Portoroz, Slovenia, 29–30 October 2014; RACSPA: Tunis, Tunisia, 2014; pp. 100–105. [Google Scholar]
- Ballesteros, E. Mediterranean coralligenous assemblages: A synthesis of present knowledge. Oceanogr. Mar. Biol. 2006, 44, 123–195. [Google Scholar]
- Ezzat, L.; Merle, P.L.; Furla, P.; Buttler, A.; Ferrier-Pagès, C. The response of the Mediterranean gorgonian Eunicella singularis to thermal stress is independent of its nutritional regime. PLoS ONE 2013, 8, e64370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, S.; Cohu, S.; Vignot, C.; Zimmerman, G.; Gattuso, J.P. One-year experiment on the physiological response of the Mediterranean crustose coralline alga, Lithophyllum cabiochae, to elevated pCO2 and temperature. Ecol. Evol. 2013, 3, 676–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Prieto, C. Light and temperature requirements for survival, growth and reproduction of the crustose coralline Lithophyllum stictaeforme from the Mediterranean Sea. Bot. Mar. 2016, 59, 95–104. [Google Scholar] [CrossRef]
- Pagès-Escolà, M.; Hereu, B.; Garrabou, J.; Montero-Serra, I.; Gori, A.; Gómez-Gras, D.; Figuerola, B.; Linares, C. Divergent responses to warming of two common co-occurring Mediterranean bryozoans. Sci. Rep. 2018, 8, 17455. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Gras, D.; Linares, C.; de Caralt, S.; Cebrian, E.; Frleta-Valić, M.; Montero-Serra, I.; Pagès-Escolà, M.; López-Sendino, P.; Garrabou, J. Response diversity in Mediterranean coralligenous assemblages facing climate change: Insights from a multispecific thermotolerance experiment. Ecol. Evol. 2019, 9, 4168–4180. [Google Scholar] [CrossRef]
- Garrabou, J.; Perez, T.; Sartoretto, S.; Harmelin, J.G. Mass mortality event in red coral Corallium rubrum populations in the Provence Region (France, NW Mediterranean). Mar. Ecol. Prog. Ser. 2001, 217, 263–272. [Google Scholar] [CrossRef]
- Cebrian, E.; Uriz, M.J.; Garrabou, J.; Ballesteros, E. Sponge mass mortalities in a warming Mediterranean Sea: Are Cyanobacteria harboring species worse off? PLoS ONE 2011, 6, e20211. [Google Scholar]
- Cocito, S.; Sgorbini, S. Long-term trend in substratum occupation by a clonal, carbonate bryozoan in a temperate rocky reef in times of thermal anomalies. Mar. Biol. 2013, 161, 17–27. [Google Scholar] [CrossRef]
- Hereu, B.; Kersting, D.K. Diseases of coralline algae in the Mediterranean Sea. Coral Reefs 2016, 35, 713. [Google Scholar] [CrossRef] [Green Version]
- Cerrano, C.; Bavestrello, G. Medium-term effects of die-off of rocky benthos in the Ligurian Sea. What can we learn from gorgonians? Chem. Ecol. 2008, 24, 73–82. [Google Scholar] [CrossRef]
- Coma, R.; Linares, C.; Ribes, M.; Diaz, D.; Garrabou, J.; Ballesteros, E. Consequences of a mass mortality in populations of Eunicella singularis (Cnidaria:Octocorallia) in Menorca (NW Mediterranean). Mar. Ecol. Prog. Ser. 2006, 327, 51–60. [Google Scholar] [CrossRef]
- Huete-Stauffer, C.; Vielmini, I.; Palma, M.; Navone, A.; Panzalis, P.; Vezzulli, L.; Misic, C.; Cerrano, C. Paramuricea clavata (Anthozoa, Octocorallia) loss in the Marine Protected Area of Tavolara (Sardinia, Italy) due to a mass mortality event. Mar. Ecol. Evol. Persp. 2011, 32, 107–116. [Google Scholar] [CrossRef]
- Teixido, N.; Casas, E.; Cebrian, E.; Linares, C.; Garrabou, J. Impacts on coralligenous outcrop biodiversity of a dramatic coastal storm. PLoS ONE 2013, 8, e53742. [Google Scholar] [CrossRef] [Green Version]
- Bavestrello, G.; Cerrano, C.; Zanzi, D.; Cattaneo-Vietti, R. Damage by fishing activities in the gorgonian coral Paramuricea clavata in the Ligurian Sea. Aquat. Conserv. 1997, 7, 253–262. [Google Scholar] [CrossRef]
- Coma, R.; Pola, E.; Ribes, M.; Zabala, M. Long-term assessment of temperate octocoral mortality patterns, protected vs. unprotected areas. Ecol. Appl. 2004, 14, 1466–1478. [Google Scholar] [CrossRef] [Green Version]
- Cebrian, E.; Linares, C.; Marschal, C.; Garrabou, J. Exploring the effects of invasive algae on the persistence of gorgonian populations. Biol. Invasions 2012, 14, 2647–2656. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Azzola, A.; Bertolino, M.; Betti, F.; Bo, M.; CattaneoVietti, R.; Cocito, S.; Montefalcone, M.; Morri, C.; Oprandi, A.; et al. Consequences of the marine climate and ecosystem shift of the 1980–90s on the Ligurian Sea biodiversity (NW Mediterranean). Eur. Zool. J. 2019, 86, 458–487. [Google Scholar] [CrossRef]
- Verdura, J.; Linares, C.; Ballesteros, E.; Coma, R.; Uriz, M.J.; Bensoussan, N.; Cebrian, E. Biodiversity loss in a Mediterranean ecosystem due to an extreme warming event unveils the role of an engineering gorgonian species. Sci. Rep. 2019, 9, 5911. [Google Scholar] [CrossRef] [PubMed]
- Mistri, M.; Ceccherelli, V.U. Effects of a mucilage event on the Mediterranean gorgonian Paramuricea clavata. 1. Short term impacts at the population and colony levels. It. J. Zool. 1996, 63, 221–230. [Google Scholar] [CrossRef]
- Giuliani, S.; Virno Lamberti, C.; Sonni, C.; Pellegrini, D. Mucilage impact on gorgonians in the Tyrrhenian Sea. Sci. Total Environ. 2005, 353, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Piazzi, L.; Atzori, F.; Cadoni, N.; Cinti, M.F.; Frau, F.; Ceccherelli, G. Benthic mucilage blooms threaten coralligenous reefs. Mar. Environ. Res. 2018, 140, 145–151. [Google Scholar] [CrossRef]
- Martin, Y.; Bonnefort, J.L.; Chancerelle, L. Gorgonians mass mortality during the 1999 late summer in french Mediterranean coastal waters: The bacterial hypothesis. Water Res. 2002, 36, 779–782. [Google Scholar] [CrossRef]
- Vezzulli, L.; Previati, M.; Pruzzo, C.; Marchese, A.; Bourne, D.G.; Cerrano, C. Vibrio infections triggering mass mortality events in a warming Mediterranean Sea. Environ. Microbiol. 2010, 12, 2007–2019. [Google Scholar] [CrossRef]
- Vezzulli, L.; Pezzati, E.; Huete-Stauffer, C.; Pruzzo, C.; Cerrano, C. 16SrDNA pyrosequencing of the Mediterranean gorgonian Paramuricea clavata reveals a link among alterations in bacterial holobiont members, anthropogenic influence and disease outbreaks. PLoS ONE 2013, 8, e67745. [Google Scholar] [CrossRef] [Green Version]
- Mistri, M.; Ceccherelli, V.U. Effects of a mucilage event on the Mediterranean gorgonian Paramuricea clavata. 2. Population recovery after two years. It. J. Zool. 1996, 63, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Linares, C.; Coma, R.; Diaz, D.; Zabala, M.; Hereu, B.; Dantart, L. Immediate and delayed effects of a mass mortality event on gorgonian population dynamics and benthic community structure in the NW Mediterranean Sea. Mar. Ecol. Prog. Ser. 2005, 305, 127–137. [Google Scholar] [CrossRef]
- Cupido, R.; Cocito, S.; Sgorbini, S.; Bordone, A.; Santangelo, G. Response of a gorgonian (Paramuricea clavata) population to mortality events: Recovery or loss? Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 984–992. [Google Scholar] [CrossRef]
- Bramanti, L.; Bendetti, M.C.; Cupido, S.; Priori, C.; Erra, F.; Iannelli, M.; Santangelo, G. Demography of animal forests: The example of Mediterranean gorgonians. In Marine Animal Forests; Rossi, S., Bramanti, L., Gori, A., Orejas Saco del Valle, C., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Cerrano, C.; Danovaro, R.; Gambi, C.; Pusceddu, A.; Riva, A.; Schiaparelli, S. Gold coral (Savalia savaglia) and gorgonian forests enhance benthic biodiversity and ecosystem functioning in the mesophotic zone. Biodivers. Conserv. 2010, 19, 153–167. [Google Scholar] [CrossRef] [Green Version]
- Casas-Güell, E.; Teixidó, N.; Garrabou, J.; Cebrian, E. Structure and biodiversity of coralligenous assemblages over broad spatial and temporal scales. Mar. Biol. 2015, 162, 901–912. [Google Scholar] [CrossRef]
- Ponti, M.; Grech, D.; Mori, M.; Perlini, R.A.; Ventra, V.; Panzalis, P.A.; Cerrano, C. The role of gorgonians on the diversity of vagile benthic fauna in Mediterranean rocky habitats. Mar. Biol. 2016, 163, 1–14. [Google Scholar] [CrossRef]
- Ponti, M.; Turicchia, E.; Ferro, F.; Cerrano, C.; Abbiati, M. The understorey of gorgonian forests in mesophotic temperate reefs. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 1153–1166. [Google Scholar] [CrossRef]
- Gori, A.; Bavestrello, G.; Grinyó, J.; Dominguez-Carrió, C.; Ambroso, S.; Bo, M. Animal forests in deep coastal bottoms and continental shelf of the Mediterranean Sea. In Marine Animal Forests; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Rossi, S.; Bramanti, L.; Gori, A.; Orejas, C. An overview of the animal forests of the world. In Marine Animal Forests; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Ponti, M.; Perlini, R.A.; Ventra, V.; Grech, D.; Abbiati, M.; Cerrano, C. Ecological shifts in Mediterranean coralligenous assemblages related to gorgonian forest loss. PLoS ONE 2014, 9, e102782. [Google Scholar] [CrossRef] [PubMed]
- Cerrano, C.; Arillo, A.; Azzini, F.; Calcinai, B.; Castellano, L.; Muti, C.; Valisano, L.; Zega, G.; Bavestrello, G. Gorgonian population recovery after a mass mortality event. Aquat. Conserv. 2005, 15, 147–157. [Google Scholar] [CrossRef]
- Santangelo, G.; Cupido, R.; Cocito, S.; Bramanti, L.; Priori, C.; Erra, F.; Iannelli, M. Effects of increased mortality on gorgonian corals (Cnidaria, Octocorallia): Different demographic features may lead affected populations to unexpected recovery and new equilibrium points. Hydrobiologia 2015, 759, 171–187. [Google Scholar] [CrossRef]
- Underwood, A.J. On beyond BACI: Sampling designs that might reliably detect environmental disturbances. Ecol. Appl. 1994, 4, 3–15. [Google Scholar] [CrossRef]
- Benedetti-Cecchi, L. Beyond BACI: Optimization of environmental sampling designs through monitoring and simulation. Ecol. Appl. 2001, 11, 783–799. [Google Scholar] [CrossRef]
- Piazzi, L.; Gennaro, P.; Montefalcone, M.; Bianchi, C.N.; Cecchi, E.; Morri, C.; Serena, F. STAR: An integrated and standardized procedure to evaluate the ecological status of coralligenous reefs. Aquat. Conserv. Mar. Freshw. Ecosys. 2019, 29, 189–201. [Google Scholar] [CrossRef]
- Cecchi, E.; Gennaro, P.; Piazzi, L.; Ricevuto, E.; Serena, F. Development of a new biotic index for ecological status assessment of Italian coastal waters based on coralligenous macroalgal assemblages. Eur. J. Phycol. 2014, 49, 298–312. [Google Scholar] [CrossRef]
- Piazzi, L.; Gennaro, P.; Cecchi, E.; Serena, F.; Bianchi, C.N.; Morri, C.; Montefalcone, M. Integration of ESCA index through the use of sessile invertebrates. Sci. Mar. 2017, 81, 1–8. [Google Scholar]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate dispersion as a measure of beta diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Benedetti-Cecchi, L.; Pannacciulli, F.; Bulleri, F.; Moschella, P.; Airoldi, L.; Relini, G.; Cinelli, F. Predicting the consequences of anthropogenic disturbance: Large-scale effects of loss of canopy algae on rocky shores. Mar. Ecol. Progr. Ser. 2001, 214, 137–150. [Google Scholar] [CrossRef] [Green Version]
- Montefalcone, M.; Vassallo, P.; Gatti, G.; Parravicini, V.; Paoli, C.; Morri, C.; Bianchi, C.N. The exergy of a phase shift: Ecosystem functioning loss in seagrass meadows of the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2015, 156, 186–194. [Google Scholar] [CrossRef]
- Carugati, L.; Gatto, B.; Rastelli, E.; Lo Martire, M.; Coral, C.; Greco, S.; Danovaro, R. Impact of mangrove forests degradation on biodiversity and ecosystem functioning. Sci. Rep. 2018, 8, 13298. [Google Scholar] [CrossRef] [Green Version]
- Valisano, L.; Notari, F.; Mori, M.; Cerrano, C. Temporal variability of sedimentation rates and mobile fauna inside and outside a gorgonian garden. Mar. Ecol. 2016, 37, 1303–1314. [Google Scholar] [CrossRef]
- Linares, C.; Cebrian, E.; Coma, R. Effects of turf algae on recruitment and juvenile survival of gorgonian corals. Mar. Ecol. Prog. Ser. 2012, 452, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Birrell, C.L.; McCook, L.J.; Willis, B.L. Effects of algal turfs and sediment on coral settlement. Mar. Pollut. Bull. 2005, 51, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.N.; Steneck, R.S.; Mumby, P.J. Running the gauntlet: Inhibitory effects of algal turfs on the processes of coral recruitment. Mar. Ecol. Prog. Ser. 2010, 414, 91–105. [Google Scholar] [CrossRef]
- Piazzi, L.; Atzori, F.; Cadoni, N.; Cinti, M.F.; Frau, F.; Ceccherelli, G. Monitoring non-indigenous macroalgae in a Mediterranean MPA: Lessons from a short-temporal variability of pristine habitats invasion. Ocean. Coast. Manag. 2021, in press. [Google Scholar] [CrossRef]
- Piazzi, L.; Gennaro, P.; Cecchi, E.; Bianchi, C.N.; Cinti, F.; Gatti, G.; Guala, I.; Morri, C.; Sartoretto, F.; Serena, F.; et al. Ecological Status of Coralligenous Assemblages: Ten years of application of the ESCA index from local to wide scale validation. Ecol. Ind. 2021, 121, 107077. [Google Scholar] [CrossRef]
- Rossi, S. The destruction of the ‘animal forests’ in the oceans: Towards an over-simplification of the benthic ecosystems. Ocean Coast Manag. 2013, 84, 77–85. [Google Scholar] [CrossRef]
- Boudouresque, C.F.; Blanfuné, A.; Personnic, S.; Ruitton, S.; Thibaut, T.; Verlaque, M. Where seaweed forests meet animal forests: The examples of macroalgae in coral reefs and the Mediterranean coralligenous ecosystem. In Marine Animal Forests; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Gatti, G.; Bianchi, C.N.; Morri, C.; Montefalcone, M.; Sartoretto, S. Coralligenous reefs state along anthropized coasts: Application and validation of the COARSE index, based on a rapid visual assessment (RVA) approach. Ecol. Ind. 2015, 52, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Ferrigno, F.; Russo, G.F.; Sandulli, R. Coralligenous Bioconstructions Quality Index (CBQI): A synthetic indicator to assess the status of different types of coralligenous habitats. Ecol. Ind. 2017, 82, 271–279. [Google Scholar] [CrossRef]
- Enrichetti, F.; Bo, M.; Morri, C.; Montefalcone, M.; Toma, M.; Bavestrello, G.; Tunesi, L.; Canese, S.; Giusti, M.; Salvati, E.; et al. Assessing the environmental status of temperate mesophotic reefs: A new, integrated methodological approach. Ecol. Ind. 2019, 102, 218–229. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa/Group | Pseudo-F4,8 | P (Perm) | Pair-Wise Test |
|---|---|---|---|
| Multivariate analysis | 4.44 | 0.003 | 2016 = 2017 = 2018 ≠ 2019 ≠ 2020 |
| encrusting coralline | 2.88 | 0.090 | |
| Peyssonnelia spp. | 0.33 | 0.810 | |
| algal turf | 49.02 | 0.001 | 2016 = 2017 = 2018 < 2019 = 2020 |
| encrusting sponges | 0.10 | 0.981 | |
| massive sponges | 2.90 | 0.073 | |
| erect bryozoans | 14.36 | 0.004 | 2016 = 2017 = 2018 = 2019 > 2020 |
| Paramuricea clavata | 17.07 | 0.003 | 2016 = 2017 > 2018 = 2019 = 2020 |
| Eunicella cavolini | 4.98 | 0.028 | 2016 = 2017 = 2018 = 2019 > 2020 |
| Source | df | MS | Pseudo-F | P (Perm) |
|---|---|---|---|---|
| Time = T | 1 | 10,175 | 5.59 | 0.007 |
| Site = S | 2 | 53,273 | 28.66 | 0.003 |
| Plot(S) = P(S) | 6 | 1858 | 4.13 | 0.001 |
| TxS | 2 | 13,359 | 7.34 | 0.002 |
| TxP(S) | 6 | 1818 | 4.04 | 0.001 |
| Residual | 162 | 449 | ||
| Pair-wise test (TxS) | D: | T1 ≠ T2 | ||
| C1: | T1 = T2 | |||
| C2: | T1 = T2 |
| Source | df | MS | Pseudo-F | P (Perm) |
|---|---|---|---|---|
| Time = T | 1 | 12.8 | 9.4 | 0.028 |
| Site = S | 2 | 175.5 | 13.21 | 0.011 |
| Plot(S) = P(S) | 6 | 13.2 | 2.9 | 0.013 |
| TxS | 2 | 96.2 | 70.72 | 0.001 |
| TxP(S) | 6 | 1.3 | 0.29 | 0.941 |
| Residual | 162 | 4.5 | ||
| Pair-wise test (TxS) | D: | T1 > T2 | ||
| C1: | T1 = T2 | |||
| C2: | T1 < T2 |
| Source | df | MS | Pseudo-F | P (Perm) |
|---|---|---|---|---|
| Time = T | 1 | 95.6 | 5.37 | 0.028 |
| Site = S | 2 | 150.7 | 8.47 | 0.009 |
| TxS | 2 | 164.2 | 9.24 | 0.004 |
| Residual | 12 | 17.7 | ||
| Pair-wise test (TxS) | D: | T1 > T2 | ||
| C1: | T1 = T2 | |||
| C2: | T1 = T2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piazzi, L.; Atzori, F.; Cadoni, N.; Cinti, M.F.; Frau, F.; Pansini, A.; Pinna, F.; Stipcich, P.; Ceccherelli, G. Animal Forest Mortality: Following the Consequences of a Gorgonian Coral Loss on a Mediterranean Coralligenous Assemblage. Diversity 2021, 13, 133. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030133
Piazzi L, Atzori F, Cadoni N, Cinti MF, Frau F, Pansini A, Pinna F, Stipcich P, Ceccherelli G. Animal Forest Mortality: Following the Consequences of a Gorgonian Coral Loss on a Mediterranean Coralligenous Assemblage. Diversity. 2021; 13(3):133. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030133
Chicago/Turabian StylePiazzi, Luigi, Fabrizio Atzori, Nicoletta Cadoni, Maria Francesca Cinti, Francesca Frau, Arianna Pansini, Federico Pinna, Patrizia Stipcich, and Giulia Ceccherelli. 2021. "Animal Forest Mortality: Following the Consequences of a Gorgonian Coral Loss on a Mediterranean Coralligenous Assemblage" Diversity 13, no. 3: 133. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030133