1. Introduction
The
Isoëtes L. genus constitutes heterosporous lycophytes widely distributed globally, occurring essentially in aquatic environments and in seasonally flooded areas [
1,
2]. This group was dominant in the Carboniferous period, forming some of the most extensive plant fossil deposits of any geological period [
3]. South America is considered a significant center of taxonomic diversity, and Brazil is a prominent region, with at least 26
Isoëtes species [
4,
5].
Isoëtes cangae and
Isoëtes serracarajensis were recently described as endemic species of
canga, the ferruginous mountain outcrops of Serra dos Carajás, Brazilian eastern Amazon [
1,
6]. Such mountain outcrops harbor several phytophysiognomies (grasslands, scrublands, wetlands, and forest formations [
7]), facing severe environmental conditions (high temperature, UV radiation, high evapotranspiration, and poorly developed soils rich in metals [
8]), harboring endemics and/or rare species [
9].
Isoëtes cangae is an aquatic species found submerged only in a permanent lake, while
I. serracarajensis is more widely distributed and found partially or totally submerged in temporary ponds or humid areas on four plateaus of Serra dos Carajás (Serra Sul, Serra Norte, Serra da Bocaina and Serra do Tarzan) [
1]. Because of its restricted distribution and habitat quality deterioration triggered by alterations in the hydroclimatological cycle in the region (prominent forest conversion into pasturelands and mining activities [
1,
10]),
I. cangae was recently classified as a critically endangered species (CR) according to the International Union for Conservation of Nature (IUCN) criteria [
11], while
I. serracarajensis is still not listed.
The development of propagation studies and the determination of their genetic structure and diversity have been the first steps toward the conservation of rare and endemic
Isoëtes species [
12,
13,
14,
15,
16]. The in vitro generation of new
Isoëtes plants can occur by sexual reproduction [
14,
15,
17] and, more rarely, by asexual or vegetative reproduction, usually observed by the division of the corm [
18,
19]. Sporophyte regeneration by sexual reproduction has been more frequently observed and is widely used for in vitro propagation [
13,
14,
15,
17,
20]. Meanwhile, asexual propagation have also been observed and described as fundamental in the reproduction of
Isoëtes andicola [
18] and
Isoëtes savatieri [
4].
The understanding of propagation, growth, and development is of paramount importance to the establishment of conservation plans for the rare and endemic species I. cangae and I. serracarajensis. While Caldeira et al. (2019) have reported sexual propagation for I. cangae by self-fecundation, studies on cross-fertilization, apomixis, and vegetative propagation are still missing. Furthermore, there are no results available regarding I. serracarajensis reproduction. Here, we investigated the reproductive strategy of both species and outlined (i) the impact of seasonality on spore germination; (ii) the influence of apomixis, self-fertilization, and cross-fertilization on sporophyte regeneration and growth; (iii) vegetative propagation; and (iv) the pattern of ex situ growth and sporangia and tiller production.
2. Materials and Methods
2.1. Plant Material
Plants of
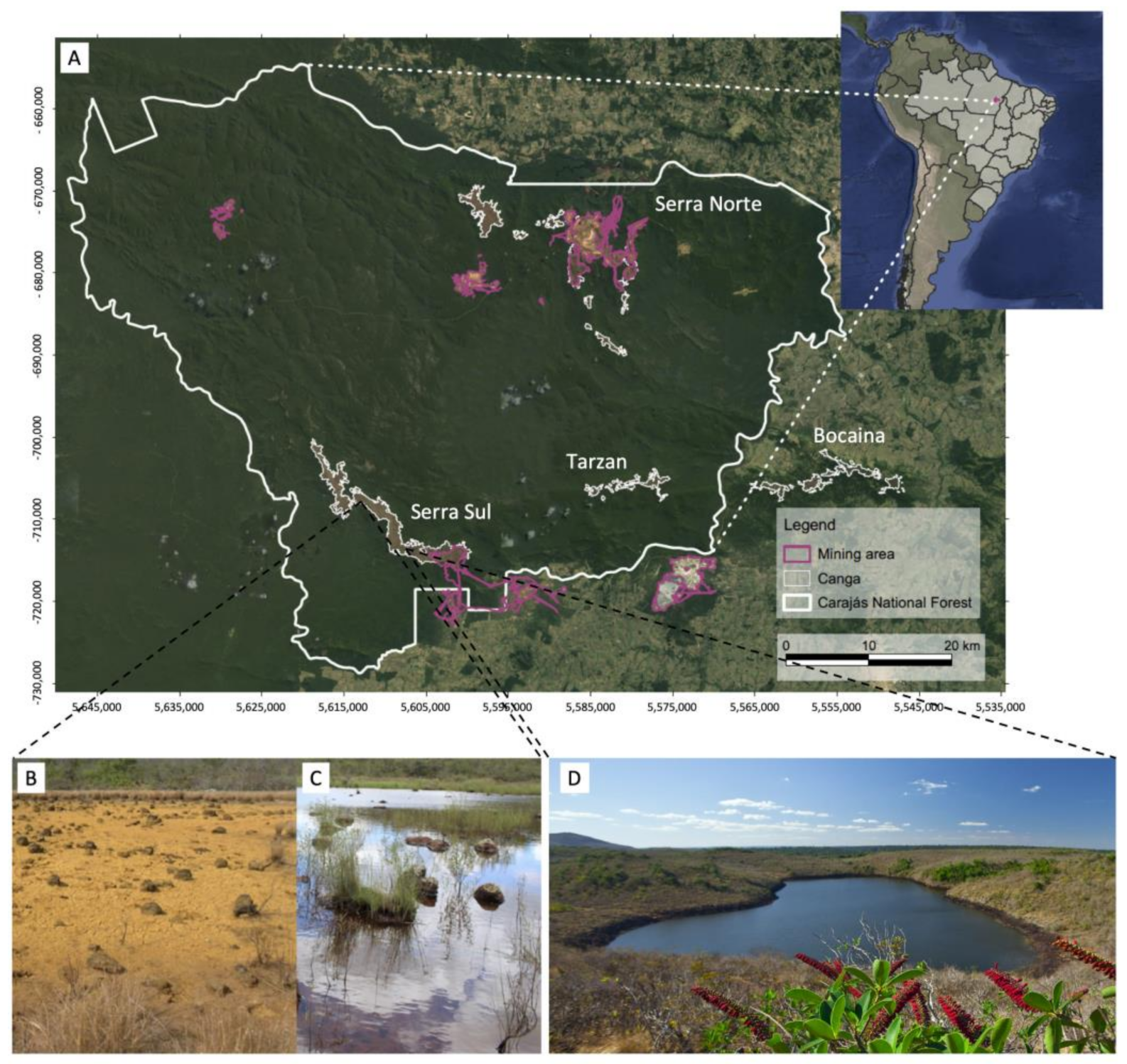
I. cangae were collected from the four portions (north, south, east, and west) of the Amendoim Lake (Serra Sul) in Serra dos Carajás, Brazil (
Figure 1) during the dry (June to October) and rainy seasons (November to May). Permission to collect the specimens was granted by the Chico Mendes Institute of Biodiversity of the Ministry of the Environment (ICMBio/MMA; numbers 641873 and 59724; SISGEN- ADC5012).
Isoëtes serracarajensis was found and collected only during the rainy season in the temporary ponds of four different iron-rich mountain outcrops harboring the
canga ecosystems of Carajás: Serra Norte, Serra Sul, Serra do Tarzan, and Serra da Bocaina (
Figure 1). A total of 220 plants of
I. cangae and 80 plants of
I. serracarajensis were collected. In the laboratory, the plants were transferred to transparent plastic containers (volume of 2 dm
3) containing substrates from their respective lake (or pond) and covered with distilled water. Some of them were maintained in controlled environmental conditions (SCG 120, Weiss Technik, UK–12:12 h photoperiod, 80 μmol m
−2 s
−1 photosynthetic photon flux density and 28:22 °C day:night temperature regime), while others were maintained in greenhouse conditions.
2.2. Spore Germination from Sporangia Collected during the Dry and Rainy Seasons
To outline the influence of seasonality on megaspore germination and in vitro fertilization, 300 mature megaspores of I. cangae (30 plants) were used per collection period (dry or rainy), and 200 megaspores of I. serracarajensis (20 plants), collected only in the dry season. The number of megaspores used was lower in I. serracarajensis due to the smaller number of fertile individuals collected. Sporangia collection and sterilization were carried out as described by Caldeira et al. (2019). Briefly, after gently removing the intact sporangia from the leaf base, they were peeled away with forceps, treated with 70% ethanol for 1 min, rinsed with sterile distilled water, sterilized again for 3 min with a solution of NaClO (1%) containing Tween-20 (0.01%), and rinsed three times with sterile distilled water.
The megasporangia were then ruptured with forceps, and the spores were counted and incubated in 50 mL Falcon tubes filled with sterile distilled water. Then, microspores were also released from the sporangia and simultaneously added to the tubes. The tubes were kept in a growth chamber (SCG 120, Weiss Technik, Loughborough, UK) under the same environmental conditions described in
Section 2.1. Megaspore germination was evaluated twice a week using a stereomicroscope, and fertilization was carried out with the germinated megaspores. The sporophyte appearance was checked and removed once every two days for 90 days. Young sporophytes were kept in distilled water until root emergence, when they were transferred to pots with substrate covered by a thin layer of sterilized quartzite sand to avoid the spread of debris [
14].
2.3. Sexual Propagation Assays
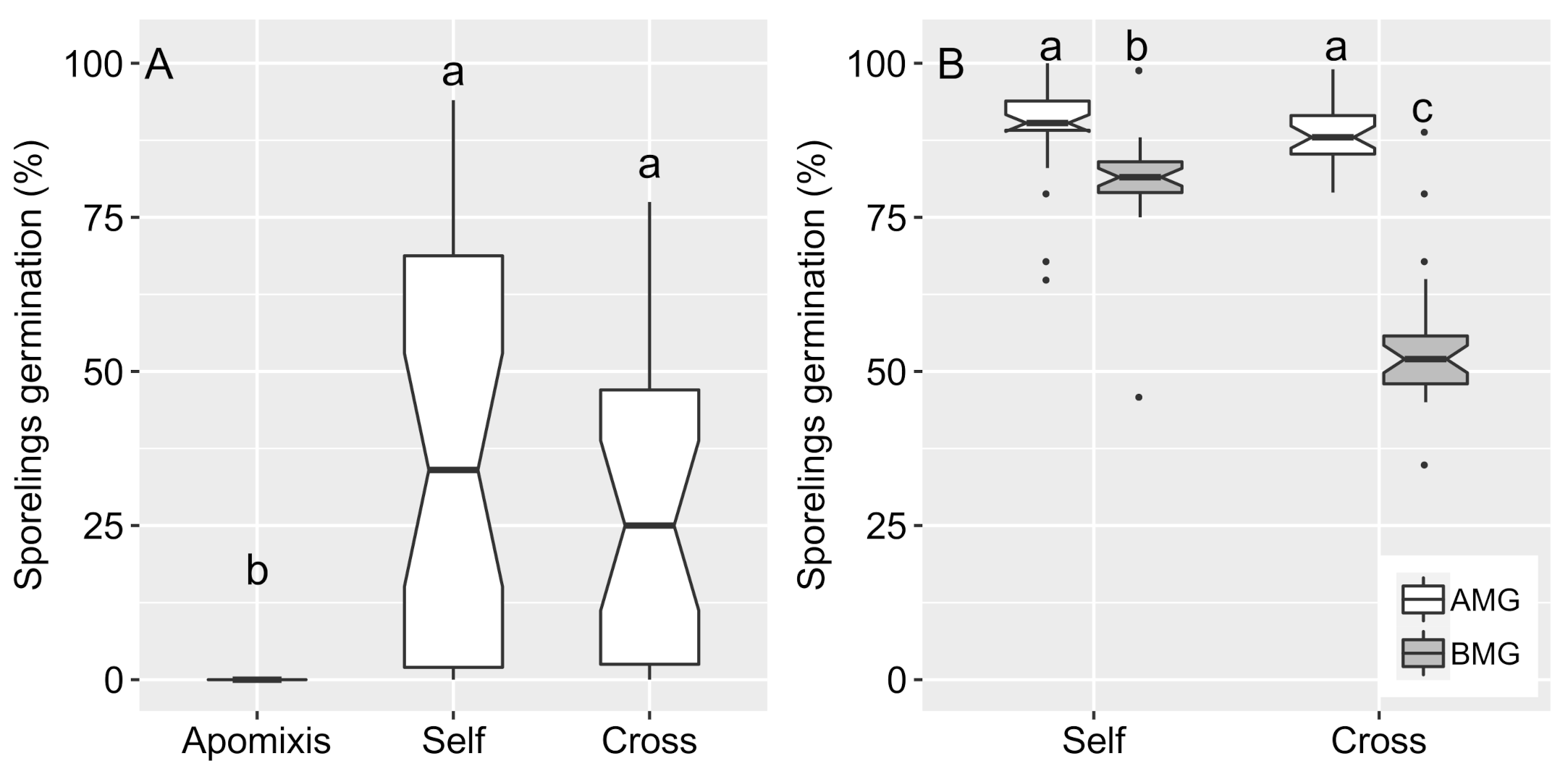
To evaluate spore germination and in vitro sporophyte regeneration, we carried out two sets of experiments with both species. In all experiments, we evaluated: (i) self-fertilization (mixed megaspores and microspores of the same individual), (ii) cross-fertilization (mixed megaspores and microspores of different individuals), and (iii) apomixis (only megaspores isolated). Initially, we used mature spores collected from 13 adult plants of I. cangae (3.520 megaspores) and two plants of
I. serracarajensis (448 megaspores). We carried out four tests for
I. cangae and one for
I. serracarajensis. The tests were conducted on different numbers of plants, following the availability of male and female sporangia (
Table S1). Sporangia collection and sterilization were carried out as described in
Section 2.2, except for the apomixis treatment, which did not receive the addition of micropores. To check for megaspore viability in the apomixis treatment, the megaspores were observed under a stereomicroscope. Subsets of germinated megaspores (marked by a trilete aperture, exposure of the archegonia, and a rhizoid appearance) were isolated in sterile distilled water. Then, after 15 days of isolation, microspores were added to the tubes once no leaf emission was observed. The sporophyte appearance was checked and removed once every two days for 90 days.
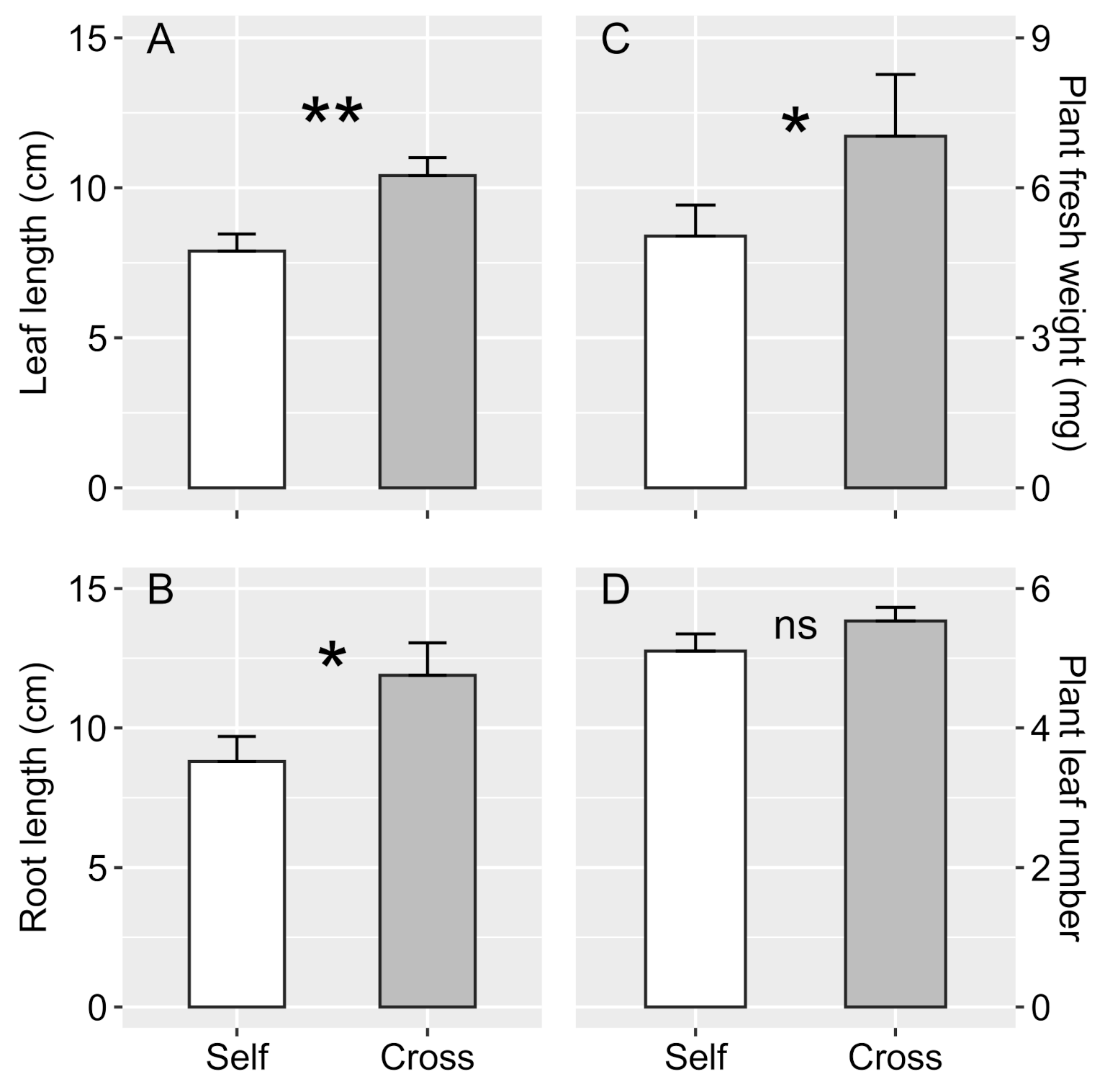
In the second set of experiments, we examined the influence of microspore addition on megaspore germination. Treatments consisted of (i) megaspore germination in the presence of microspores, i.e., micropores added simultaneously to the mixture with megaspores, and (ii) megaspore germination in the absence of microspores, which were added after archegonia exposure and rhizoid emission. In this case, the germinated megaspores were periodically removed from the mixture and transferred to other tubes containing microspores. These experiments were carried out considering the effect of self- and cross-fertilization, totaling four treatments with 300 megaspores each for I. cangae e 100 megaspores for I. serracarajensis (10 megaspores collected per plant of each species). Because of the absence of megaspore germination for I. serracarajensis, we did not carry out the treatment involving microspore addition after germination. Apomixis tests were also performed with the same number of megaspores for each species. Sporangia collection and sterilization, spore incubation, and germination were carried out under the same environmental conditions as described previously. Likewise, the sporophyte appearance was checked and removed once every two days over 90 days, and the young sporophytes were transferred to pots with substrate and distilled water. After growing for 90 days, we counted the number of leaves and measured the plant height, root length, and fresh weight of the entire plant.
Nuclear DNA Extraction and PCR Amplification of ISSR Markers
The plants generated in the apomixis treatment were analyzed to compare them with the mother plants’ total genomic DNA using ISSR molecular markers. For that, the total genomic DNA was isolated from 200 mg of leaves. The leaf tissues were macerated in liquid nitrogen, and total DNA extraction was performed following the modified CTAB protocol [
21,
22]. The DNA quality was evaluated with an agarose gel (1%) and its concentration was estimated using a Nanodrop
®. The DNA samples (20 µg/µL) were used to amplify the DNA by PCR using genetic ISSR markers (
Table S2). The proportion of the same or different bands within each family (the mother plant and its respective progeny) was observed, and the percentage of polymorphic bands between parent plants and progeny was calculated as described by Santos et al. (2020).
2.4. Isoëtes cangae Ex Situ Growth and Sporangia Production
Isoëtes cangae sporophyte growth and development were monitored in experiments carried out in a controlled environment (growth chamber) and under greenhouse conditions. Approximately two thousand sporophytes were continuously generated from the procedures described by Caldeira et al. (2019) and cultivated in growth shelters with the same environmental conditions described in
Section 2.1. These young sporophytes (1 cm long) were cultivated in organic substrates (Jiffy-7
®), lake sediments, and a mixture (1:1) of both substrates. In all cases, the substrates were covered with a layer of sterile quartzite sand, and distilled water was used to fill the containers. The sporophytes were monthly counted to estimate the survival rate and to check for new individuals. Some of these sporophytes (approximately 300 plants) were kept in these environmental conditions for a period longer than 24 months. Some of them were cultivated in groups of three plants in glass containers of 9 dm
3 (with 0.9 dm
3 of substrate), while others were isolated in plastic pots of 2 dm
3 (a single plant per pot, with 0.2 dm
3 of substrate). Fifty plants aged 12 months were randomly selected to determine the number of leaves and sporangia. The leaves were manually detached from the plant base (corm) to check for the presence of sporangia.
In a second experiment, a group of 130 young (nonfertile) and 90 adult (fertile) plants collected in the field were transferred to glass containers (70 L) with 15 plants (young) or individually in plastic beakers of 4 L (adult). All sporangia from the adult plants were removed before planting. The plants were cultivated in a greenhouse for 12 months, using organic substrates (Jiffy-7®) and following the procedures suggested by Zandonadi et al. (2021). Sporangia production and sporophytes apparition was monitored during this period. The plant families were evaluated, and the progenies generated were analyzed and compared to the mother plants’ total genomic DNA using ISSR markers.
2.5. Vegetative or Asexual Propagation Assays
Asexual propagation was evaluated in
I. cangae during the cultivation of a large number of plants (
Section 2.4) and in two series of experiments for
I. serracarajensis. Initially, 25 young plants collected in the field were transferred to individual pots with a mixture of organic substrate (Jiffy-7
®) and lake sediments covered by sterile quartzite sand. They were cultivated in growth shelters away from the
I. cangae plants. After 12 months, we examined the appearance of new plants and counted the tillers attached to the corm of the mother plant. We counted the number of leaves and sporangia at their bases. Then, the tillers recovered were transferred to pots with substrate and cultivated under the same environmental conditions. Their survival was evaluated three months later.
The second experiment was carried out with 20 individuals of I. serracarajensis collected in the field and cultivated under greenhouse conditions in individual pots containing organic substrate (Jiffy-7®). After 12 months, we counted the tillers attached to the plant corm, collected the mother and daughter plants, and examined the families (mothers and daughters) by using ISSR markers.
2.6. Data Analysis
Data analyses were conducted in R version 3.5.2 [
23]. We performed a Shapiro–Wilk normality test to define the subsequent parametric (Student’s
t-test) or non-parametric (Kruskal–Wallis rank test or Mann–Whitney U test) post-hoc tests. For spore germination in the three propagation methods (apomixis, self, and cross-fertilization) and to examine the influence of microspore addition on megaspore germination, we applied a post-hoc Dunn’s test (
p < 0.05). When applicable, the R function
lm() was used to generate regression models. The figures were built using the R package ggplot2.
4. Discussion
Our results suggest that the two endemic and rare
Isoëtes species from Serra dos Carajás show distinct reproductive strategies:
I. cangae reproduces by spore germination (sexual propagation) and
I. serracarajensis reproduces by emitting tillers from the plant corm (vegetative propagation). Sexual propagation has been reported as a common strategy in the
Isoëtes genus [
13,
15,
17,
20] and was previously described as occurring in
I. cangae [
14]. We found that
I. cangae is able to reproduce via self- and cross-fertilization, excluding the occurrence of vegetative propagation and apomixis. The few individuals obtained from megaspore isolation in this species can be attributed to possible fertilization before sporangia collection, as indicated by ISSR analysis. Otherwise, apomixis should be considered a rare event in
I. cangae. In fact, apomixis is still rarely reported for
Isoëtes L. but is fundamental in the propagation of
Isoëtes andicola [
18].
In this study, we obtained a wide range of variations in sporophyte regeneration from in vitro fertilization assays. Higher rates of sporophyte regeneration have been observed for other species, such as
Isoëtes malinverniana [
24],
Isoëtes lacustris [
17], and
Isoëtes sabatiana [
25], and have been attributed to the use of mature reproductive structures, while immature spores could lead to less effective sporophyte regeneration [
15,
25] or even to nongermination, as previously reported for
I. cangae [
14]. Consequently, the lower sporophyte regeneration obtained in some of our experiments could be affected by the use of spores that were not completely developed. Despite their brownish color at sampling, the sporangia were collected mostly from cultivated plants less than 24 months old. Nonetheless, we cannot exclude the hypothesis that for
I. serracarajensis, since this species inhabits seasonal ponds, it has only a few months (the rainy season) in which to grow and reproduce. In that case, the sporangia were sampled from 12-month-old plants.
The sexual propagation of
I. cangae is an important way to expand its genetic diversity and reduce the risks associated with inbreeding. In recent studies carried out with ISSR markers [
16] and a large number of SNPs [
26], it was revealed that
I. cangae consists of a single panmictic population with a moderate levels of genetic diversity and no inbreeding signal. Such diversity is likely to result from the male gamete [
27] and young sporophytes movements in the Amendoim lake [
28], increasing the possibility of cross-fertilization. As observed in this study, such sporophytes showed a faster leaf and root initial growth. Furthermore, the success of conservation plans is based on preserving the genetic diversity of related species. However, the proximity of male and female sporangia and the gradual release of spores through the decomposition of sporangium tissue increases the chances of self-fertilization in
Isoëtes [
29,
30], especially in areas of standing water, such as ponds and swamps, as described by [
31] for the species
I. hypsophila.
The vegetative or asexual reproduction observed in
I. serracarajensis is probably a less common method reported for the
Isoëtes genus, although it has been described as the main reproductive strategy for species such as
Isoëtes savatieri and
Isoëtes chubutiana [
4]. As observed for
I. serracarajensis, these two species are also able to produce megaspores and microspores, although their reproduction is still linked to tiller bud development from the plant corm. As reported for other related species [
32,
33,
34], this reproductive strategy can be advantageous for
I. serracarajensis colonizing adverse environments such as the temporary ponds of Serra dos Carajás, where the short period with available water (five to six months) would not be enough to achieve sexual propagation, i.e., not enough time for young sporophytes to grow and disperse their reproductive structures regenerating new plants able to face severe drought in the region [
8,
28]. In fact, unlike
I. cangae, which remains underwater throughout the year and faces less pronounced environmental changes [
28],
I. serracarajensis has been observed only during the rainy season (November to March). Field observations suggest that water shortage is accompanied by leaf drying and fall, while the plant corm remains protected and alive in the soil until the beginning of the next rainy season, when growth resumes (
Figure 1). However, we cannot rule out the possibility that plants of
I. serracarajensis two years old or older may indeed produce substantial reproductive structures during the rainy season. Then, desiccation and refilling of the temporary ponds may trigger spore germination so that sexual reproduction becomes synchronized with favorable environmental conditions. Interestingly, plants of
I. serracarajensis remained alive and kept green leaves throughout the 12 months in the presence of available water, suggesting that such species can show continuous growth once in favorable environments.
Although
I. cangae does not face strong environmental seasonality as
I. serracarajensis does, we did observe a significant difference in sporophyte regeneration from spores collected in field-growing adult plants during the rainy and dry seasons. Marked variability in spore germination has been described for other species, such as
I. lacustris, when changes in temperature trigger germination and growth [
17]. Overall, it has been reported that fine-tuning synchrony with environmental conditions could be advantageous for plant performance under both stressed and nonstressed conditions [
35]. Therefore, this synchrony with environmental conditions is still an important trait conserved by both
I. cangae and
I. serracarajensis.
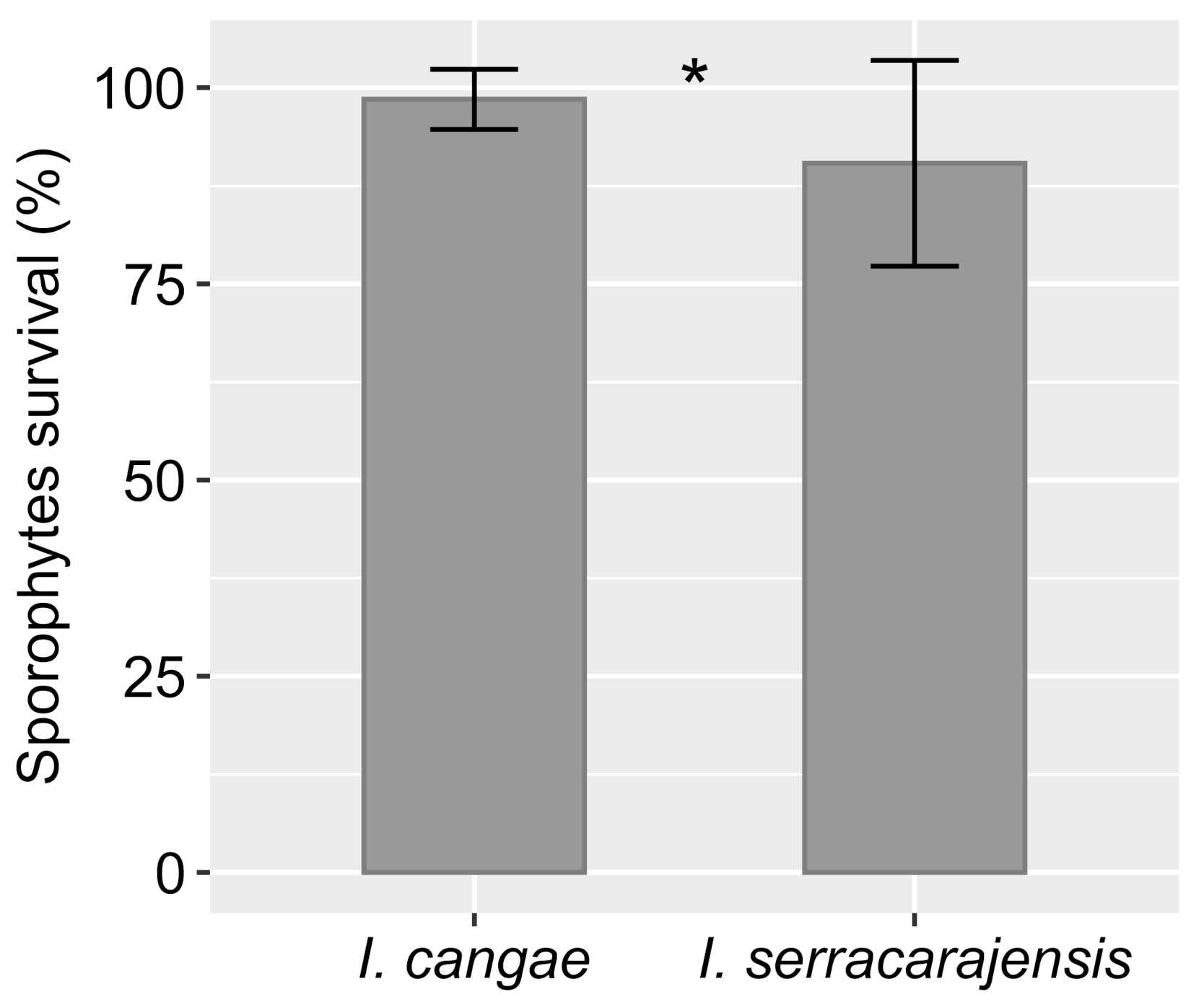
Sporophytes of
I. cangae can produce a large number of viable sporangia when they reach the adult phase growing in favorable conditions, showing a general tendency of larger mother plants producing larger amounts of offspring. Under nonstressing environmental conditions, it is well established that plants developing larger leaf areas to improve their light interception capacity and carbon fixation as well, developing higher yields and seed production [
36,
37]. However, this pattern was not clearly observed in
I. serracarajensis, as no correlation between plant size and tillering rate or the number of sporangia was found. Nevertheless, a dichotomy was observed between tillering rate and sporangia production. Their distinct reproductive strategy notwithstanding, both species (
I. cangae and
I. serracarajensis) showed successful sporophyte survival. Such successful propagation and growth, previously reported for the earlier developmental phases of
I. cangae [
14], were confirmed here for this species and extended for
I. serracarajensis, reinforcing that both species are resilient and able to grow and thrive ex situ and possibly in other environments than those currently occupied.
In this study, we outlined the distinct reproductive strategy of the two endemic Isoëtes species of Serra dos Carajás and attributed that difference to the contrasting habitat occupied by each of them. We improved the available knowledge of the reproductive biology of both species, showing that sporophytes of I. cangae can be propagated in larger numbers from mature plant spores, requiring the release of the male gamete to fecundate the megaspore, while I. serracarajensis displays vegetative propagation by developing new sporophytes from the plant corm. We also highlighted details of plant growth and development and the relationship between plant growth and reproductive traits. Finally, we demonstrated that both species could produce sporophytes with a high survival rate. Together, these results are of paramount importance to establish conservation plans for both species, considering the advantages of sexual propagation to maintain the genetic diversity of I. cangae and the diligent management required to do the same with asexually propagated I. serracarajensis.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






