
The Contribution of Desert-Dwelling Bats to Pest Control in Hyper-Arid Date Agriculture
by
 , and
, and
Jessica Schäckermann
1,2,3,* ,
,
Evie J. Morris
4,5,
Antton Alberdi
6,
Orly Razgour
4 and
Carmi Korine
7 1
Southern Arava R&D, Eilot Regional Council, D.N. Hevel Eilot 8882000, Israel
2
The Arava Institute for Environmental Studies, D.N. Hevel Eilot, Ketura 8884000, Israel
3
Ben-Gurion University of the Negev, Eilat Campus, Sderot HaTmarim 90, Eilat 8855630, Israel
4
Biosciences, University of Exeter, Streatham Campus, Hatherly Laboratories, Prince of Wales Road, Exeter EX4 4PS, UK
5
Biological Sciences, University of Southampton, Life Sciences Building, Highfield Campus, Southampton SO17 1BJ, UK
6
Center for Evolutionary Hologenomics, Globe Institute, University of Copenhagen, Øster Farimagsgade 5, 1353 Copenhagen, Denmark
7
Mitrani Department of Desert Ecology, Jacob Blaustein Institutes for Desert Research, Ben-Gurion University of the Negev, Midreshet Ben-Gurion 8499000, Israel
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(12), 1034; https://0-doi-org.brum.beds.ac.uk/10.3390/d14121034
Submission received: 15 October 2022
/
Revised: 12 November 2022
/
Accepted: 18 November 2022
/
Published: 26 November 2022
(This article belongs to the Special Issue Key Aspects in the Ecology and Diversity of Desert-Dwelling Bats)
Abstract
:Over 40% of the Earth’s surface has been converted to agricultural use and agroecosystems have become important habitats for wildlife. In arid regions, intensive agriculture creates artificial oasis-like habitats due to their high irrigation inputs. Date production is one of the primary agricultural practices in the deserts of the Middle East and North Africa. Insectivorous bats are known to use agricultural areas, but the role of date plantations as their foraging habits and the importance of insectivorous bats as date bio-pest control agents are still unknown. We assessed the role of date plantations as foraging habitats for local desert bat species by acoustically recording bat activity in conventional and organic date plantations in the southern Arava Valley, Israel. In addition, we captured bats in the plantations and collected feces for DNA metabarcoding analysis to investigate the presence of pest species in their diets. We found that 12 out of the 16 known species of bats in this region frequently used both conventional and organic date plantations as foraging habitats. Species richness was highest in the organic plantation with complex ground vegetation cover. Foraging activity was not affected by plantation type or management. However, bat species richness and activity increased in all plantations during summer date harvesting. Molecular analysis confirmed that bats feed on a variety of important date pests, but the particular pests consumed and the extent of consumption varied among bat species. Our results highlight a win–win situation, whereby date plantations are an important foraging habitat for desert bats, while bats provide bio-pest control services that benefit the date plantations. Therefore, date farmers interested in bio-pest control should manage their plantations to support local desert bat populations.
1. Introduction
Agricultural land expanses are growing worldwide, and as a result agroecosystems have become increasingly relevant habitats for wildlife [1,2]. Arid lands make up 41% of the total land area on Earth [3] and pose numerous challenges for agriculture. However, specific agricultural practices for desert environmental conditions have been developed in Israel’s arid regions since the Stone Age [4,5,6]. Today, these agroecosystems are dominated by vineyards, date plantations [7], fields of melon, pumpkin, onions, and other arable crops. Agricultural patches in desert landscapes may act as artificial oases for wildlife [8]. These artificial oases attract crop pests, natural enemies like wasps and spiders [9,10], and vertebrates such as insectivorous bats [11,12].
Insectivorous bats that use agricultural land for foraging may play an important role in regulating nocturnal insect populations in both natural and agricultural ecosystems [13,14]. Bats are known to forage in agricultural areas, and their contribution to pest suppression has been shown in several crops worldwide, such as corn [15], cotton [16,17], cacao [18], macadamia [19], and rice [20]. The economic value of bat pest control services is substantial: 1 billion USD globally/year in corn [15], 3.7 billion USD/year in all North American crops [13], and 21 Euro/ha in Mediterranean rice paddies [20].
The southern Arava is located in the hyper-arid desert region of southern Israel [21]. Agricultural produce is a major source of income in the region, derived primarily from date farming [22]. Insect pests, particularly night-active moths and beetles, are the main threats to crop production in date plantations [23,24]. Pesticides exist for the treatment of many pests [25,26,27], but not for all, and some farmers report that specific pests show resistance to pesticides (personal communication, Schäckermann). Farmers follow a protocol for pesticide application to achieve optimal results, but these protocols require extensive labor and can be expensive if several rounds of pesticide application are necessary [22]. To fulfill local and export regulations according to Global GAP trademark standards, the agricultural products must meet strict maximal pesticide residual standards [28]. The last pesticide application in date palm plantations must be applied at least 80 days prior to harvest (some farmers maintain 150 days), and in the post spraying period, the fruits are unprotected.
The Arava region is also known for its diversity of desert-dwelling bats, with 16 recorded insectivorous bat species [11,29]. The replacement of natural habitats by agricultural expansion in an ever-developing environment is likely to also affect the resident bat species. We hypothesized that bats use agricultural habitats for their foraging activities and that bat activity and species richness is associated with date plantation characteristics and their management style. We predicted that bats consume known pest species and that the diversity and activity of bats is higher in date plantations with organic management compared to conventional management. In addition, we predicted that both diversity and activity increase during the harvesting periods in all types of date plantations due to increased human activity that displaces pests, causing them to swarm and providing easier access to the bats that feed off of them.
2. Materials and Methods
2.1. Study Areas and Study Crop
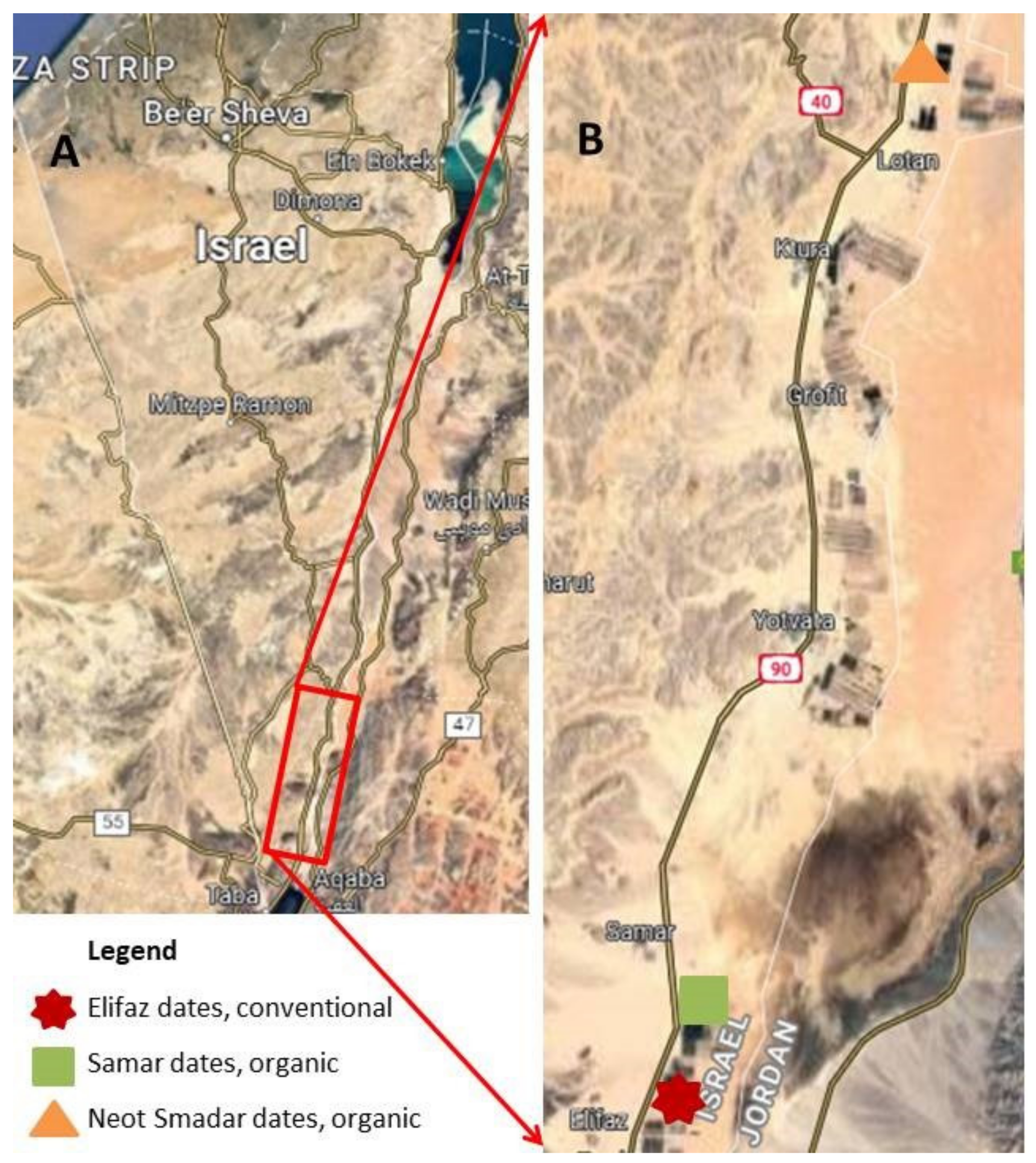
The study was carried out in and next to date plantations in the southern Arava, Israel. The acoustic monitoring was conducted in one conventional orchard (Elifaz), and the only available organic date plantations (Samar and Neot Smadar) in the southern Arava valley (Figure 1), which is a hyper-arid desert (following classifications in Noy-Meir, 1973) [30] with a mean annual temperature of 25.4 ℃ (January: 15.8 ℃–July: 33.8 ℃) and average annual rainfall of 27.6 mm (Israel Meteorological Service, https://ims.gov.il/en/ClimateAtlas accessed on 5 May 2022). The mist netting for fecal sample collection was carried out within date plantations of the region and in adjacent sites within a 1 km radius of the next date plantation (Supplementary Table S1). The date palm (Phoenix dactylifera L.) is the most important fruit crop in the arid climate regions of North Africa and the Middle East [31]. The date fruit industry is a highly important major source of income along the arid Jordan River and Arava valleys [22]. The flowering season of dates lasts from February to April, and fruit harvest takes place from August to October. Fruit bunches in most plantations are covered with net bags two months before harvest to protect them from fruit bats, birds, and fruit moths [22]. However, some pests damage other parts of the tree (e.g., branches, roots, and trunks), and some attack the fruits at an earlier stage before they are covered [32]. The lesser date moth, Batrachedra amydraula (Meyerick, 1916), is a highly specific, major pest of dates. It attacks the fruits about six months before harvest [23]. During the larval stage, the moth penetrates the young green fruit, feeds on the pulp, and can cause up to 75% yield loss if not treated [24,26]. Five annual generations of moth were recorded in plantations of the Arava Valley. The first three were the major generations, emerging periodically from March to July, creating massive peaks in the moth population, followed by peaks in larvae infestation. The next two generations were considerably smaller, occurring in August and September with no consequential larval infestation [26]. Larvae of the third to fifth generations underwent diapause until the next spring [26,33]. Currently, Batrachedra outbreaks are treated with insecticides [33,34]. In addition, the use of pheromone traps is under development, and some natural enemies of the moth have been identified—the main ones being predatory ladybug beetles and several parasitoid wasps [23,24,26]. Several parasitoids were shown to infest larvae or eggs of Batrachedra, but their efficacy as control agents is unclear [24].
The date plantations of Elifaz and Samar are nearby, separated by other crops and an open area of approximately 400 m. The date plantation of Neot Smadar is located 15 km north of the two other plantations (Figure 1). All three date plantations differ from each other in management and ground cover. Farmers of the region (both conventional and organic) treat plantations with an integrated pest management (IPM) approach, in which the caterpillar abundance of the lesser date moth is monitored between the end of February and the end of June on a weekly basis, in cooperation with the Israeli Ministry of Agriculture and Rural Development and the local extension service. Based on the abundance of caterpillars, chemical pesticides are applied two to three times per year. Conventional farms also remove the ground-covering weeds using both physical methods and chemical herbicides. Organic growers do not use herbicides. In Neot Smadar, ground-covering weeds and shrubs are removed by donkeys and camels, and in Samar, weeds and shrubs are cut with machinery once or twice a season, leaving a patch of green vegetation around the trunk of the tree [22]. In addition, fruits are only covered with bags in Elifaz and Neot Smadar.
2.2. Acoustic Monitoring
Bat foraging activity within the date plantations of Neot Smadar, Elifaz, and Samar was recorded using SD2 AnaBat bioacoustic detectors (Titley Electronics, Ballina, Australia). Each detector was placed on the ground at a 45° angle at the sampling point, facing the date palms. In each plantation, we selected five sampling plots at a distance of > 400 m from one another to avoid recording calls of the same individuals at different locations. Elifaz and Samar plantations are close to each other (Figure 1), and therefore, the nearest recording plot between the two plantations was more than >1000 m. We recorded bat activity for five full nights both before and during the harvesting. Recordings were made simultaneously at all the sites. In all three locations, sampling plots consisted of adult trees of approximately 15–20 years of age. Sampling was conducted for five contiguous nights in July 2018, before the harvesting of the dates, and again for five nights during the harvesting period in September 2018. Recordings were not taken within five days on either side of the full moon. Due to technical problems, one detector did not record the calls of the bats in Elifaz before the harvesting of the dates, and our sample size for this plantation was based on only four plots. In addition, due to technical problems, we did not have recordings from 7 and 9 nights before and during the harvest, respectively. Thus, our total sampling nights before and after the harvesting were 63 and 66, respectively. We monitored foraging activity by identifying and counting bat passes [35]. Prior to the recording sessions, all detectors were calibrated to a sensitivity level of 7, using a constant and monotonic ultrasonic signal. Calls were analyzed manually using the software AnalookW version 3.8 v. Identification of bat calls and species was determined based on known species-specific acoustic characters [36,37].
2.3. Statistical Analyses
We used generalized linear mixed models (GLMMs) with bat species richness and bat activity (passes per night) as response variables to compare the three date plantations (Elifaz, Samar, and Neot Smadar) during each recorded stage (before and during the harvesting of the dates). For each night of recordings, we obtained hourly weather conditions (temperature, relative humidity, and wind velocity) and used their night average as co-variables. Weather data were obtained from the Yotvata meteorological station located 5 km north of Elifaz and Samar (https://ims.gov.il/en accessed on 5 May 2022). The recording date of the night, sampling plot, and detector number were entered as random factors. All co-variables were checked for autocorrelation. When a significant treatment effect was found, we used Tukey’s post hoc tests to establish differences among treatments. We used t-tests to compare species richness and bat activity before and during the harvesting. Significance was set at p < 0.05.
2.4. DNA Metabarcoding Protocols
Fecal samples for DNA analysis were obtained directly from bats caught within date plantations (“agricultural” sites, N = 7) and at sites within 1 km of a date plantation (“adjacent” sites, N = 4) (Supplementary Table S1). Nets were set up at locations within the plantations where possible. Adjacent sites are located within 1 km of date plantations. In no cases did netting overlap with acoustic sampling. Bats were captured between April and September 2018 using ground-level monofilament mist nets. Captured bats were identified using morphological measurements. Individuals of the six species, Eptesicus bottae, Hypsugo ariel, Pipistrellus kuhlii, Pipistrellus rueppellii, Plecotus christii, and Otonycteris hemprichii were retained in clean cloth bags for up to one hour. Any fecal matter produced during this time was collected and transferred immediately to absolute ethanol. All samples were frozen (−20 °C) within 24 h of collection and remained so until required for further analysis.
We extracted DNA from one to five bat droppings (dry weight 15–20 mg) per bat using the PowerSoil®® DNA Isolation Kit (MoBio, CA, USA). The manufacturer’s protocol was used with the modifications detailed in [38]. Each extraction round included nine bat fecal samples and one negative control. Following the one-locus-several-primer-sets methodology (as set out by [39]), a fragment of DNA was amplified from the mitochondrial gene, cytochrome oxidase C subunit 1 (COI). We used two primer sets: ZBJ (amplicon length: 157 bp), which has been a stalwart of metabarcoding in the past decade [40] and the recently developed ANML set (amplicon length: 180 bp) [41]. In order to optimize the PCR amplifications, qPCR with SYBR green chemistry was carried out on a subset of samples and negative controls [42]; specifically, we tested the amplification dynamics in the presence of different volumes of BSA (1.0, & 0μL). QPCRs were carried out on an Agilent Technologies Stratagene Mx3005P qPCR Thermocycler (Agilent Technologies, Santa Clara, CA, USA).
Metabarcoding was carried out on all sample extractions and all negative extraction controls. We exclusively used primers carrying matching tag combinations when performing the metabarcoding PCRs (F1-R1, F2-R2, etc.), which allowed us to build PCR replicates into each Illumina library while avoiding tag jumps that cause false assignments of sequences to samples [43]. PCR products were pooled into 12 libraries per primer and sequenced using an Illumina MiSeq. Resultant reads were trimmed, merged, and de-replicated using cutadapt, usearch, and vsearch [44,45,46] before being filtered for chimeras and clustered into OTUs (operational taxonomic units) using usearch (unoise3 default settings). These sequences were uploaded to the mBrave platform [47] (www.mbrave.net, accessed on 10 May 2022) and, with minimal further processing, compared to the BOLD database to assign taxonomy; mBrave only returns species-level taxonomic assignments, which greatly simplified the search through the results for known date-pest species.
3. Results
3.1. Acoustic Monitoring
Our acoustic dataset included a total of 87,682 bat calls from 11 bat species before and 12 species during the harvesting of the dates (Table 1). We found a total species richness of 11 bats in Samar, 11 bats in Neot Smadar, and 9 bats in Elifaz before harvest. During the harvest, we found a total species richness of 12 bats in Samar, 11 bats in Neot Smadar, and 11 bats in Elifaz (Table 1). In both periods, Hypsugo ariel and Eptesicus bottae were the most common species, while Plecotus christii and Rhinolophus clivosus were the rarest species in all three date plantations. Rhinopoma microphyllum was only recorded during the harvesting period in all three plantations. Barbastella leucomelas was recorded in all three plantations before and during the harvesting of the dates (Table 1).
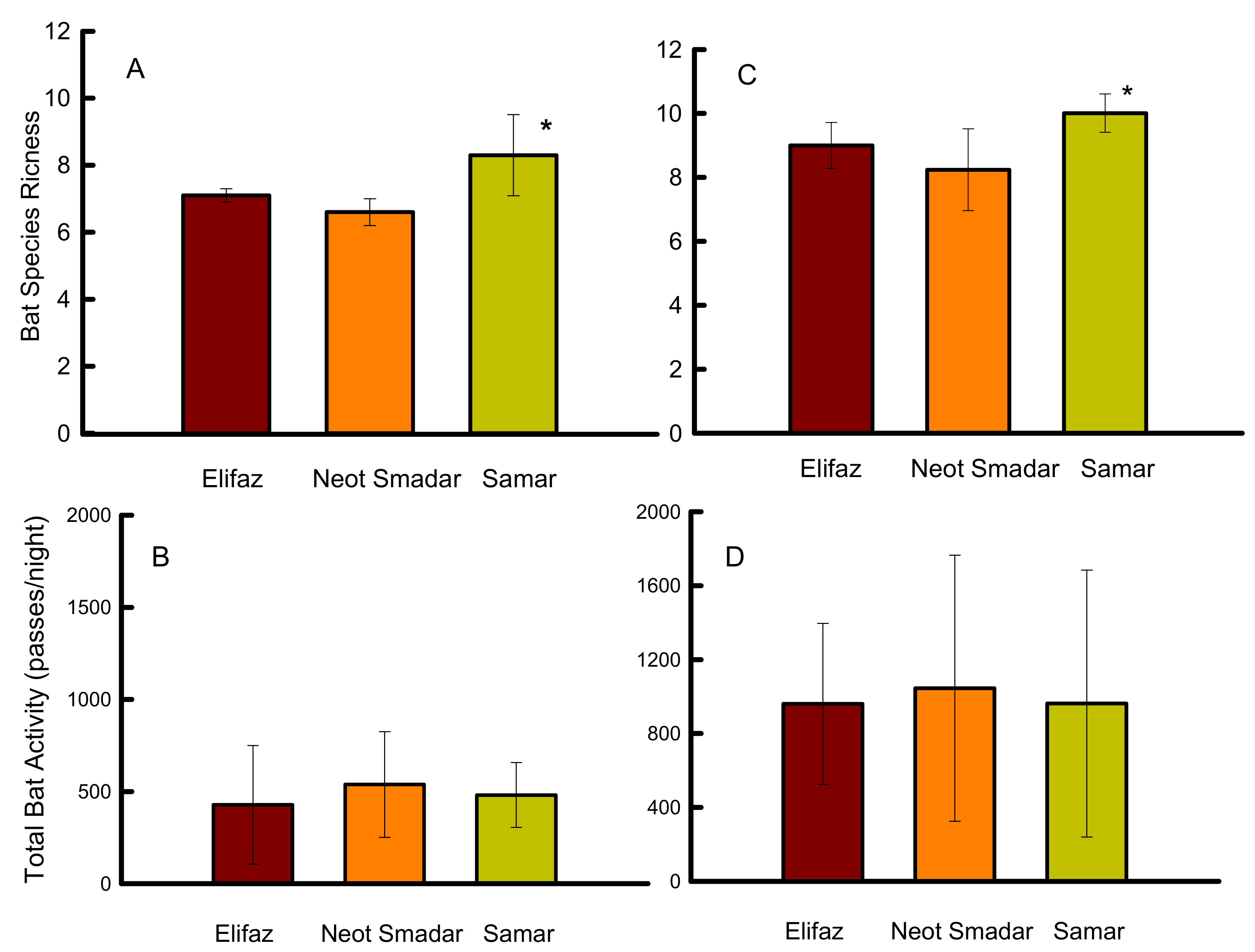
Species richness was not associated with any of the abiotic measures or the day of the recordings. Average species richness of bats before harvest was significantly higher at the Samar date plantation compared to the Elifaz and Neot Smadar plantations, and did not differ between Elifaz and Neot Smadar. Average species richness was affected by the date plantation itself and by the recording plot within the plantation (F2, 63 = 14.0, p < 0.001; r2 = 0.46, Figure 2A). Bat activity before the harvest was only affected by the plot of the recordings (F2, 63 = 3.7, p < 0.007; r2 = 0.46). Bat activity did not differ (p > 0.13) between the three plantations before harvest (Figure 2B).
Average species richness during the harvest was higher at the Samar date plantation (F5, 64 = 5.2, p < 0.001; r2 = 0.37, Figure 2C) compared to the two other plantations, and was affected by the sampling plot (p < 0.04). Bat activity during the harvest was also affected by the plot (F2, 64 = 15.7, p < 0.001; r2 = 0.55) within the plantations, but there were no differences in bat activity between the three date plantations (Figure 2D).
The combined species richness and total bat activity for all three plantations was higher during the harvest compared to the pre-harvesting period (t = 4.32, p < 0.0001 and t = 2.77, p < 0.012, respectively). When analyzed individually for each plantation, species richness was higher during the harvest compared to the pre-harvest period, while bat activity did not differ between the two periods (Table 2).
3.2. Consumption of Insect Pests by Bats in Date Plantations
A total of 79 samples were included in the metabarcoding analysis (Ebottae: N = 25, H. ariel: N = 11, Otonycteris hemprichii: N = 16, Pipistrellus kuhlii: N = 8, Pipistrellus rueppellii: N = 11, and P. christii N = 8). Sixteen known date pest species had reference sequences on the BOLD database. Of the 16 potential pest species, nine (56%) were identified in the droppings of the studied bats. The droppings of all bat species, but not all individuals, contained some evidence of pest consumption (Table 3). In almost all cases, any given pest species was only detected in a few individuals per bat species. In the case of E. bottae, O. hemprichii, and P. rueppellii, over half of the individuals sampled showed evidence of consuming at least one pest species. P. christii showed some evidence of pest consumption, with Lepidopteran pests identified in the droppings of 37.5% of the individuals. The most commonly identified pest from this species was Cadra figulilella (the raisin moth). Otonycteris hemprichii consumed the widest diversity of pest species, with seven pest species detected across 16 individual bats. Further, 69% of the studied O. hemprichii individuals consumed pests, the highest proportion of positive samples for any of the studied bat species (Table 3). While most pest species were only identified from one or two bat individuals, all eleven were found to have consumed Oryctes agamemnon (the Rhinocerus beetle). Eptesicus bottae consumed an equally high diversity of pest species (seven), and 44% of samples contained evidence of pest consumption. Two pest species were found in more than one sample: the Dubas bug, Ommatissus lybicus, and the lesser date moth. The former was also consumed by the other three cultured space aerial hawking bats (Table 1). The lesser date moth was the only other pest identified in the diet of P. kuhlii. Pipistrellus rueppellii showed evidence of consumption of two Coleopteran species. Pipistrellus rueppellii also had the second highest proportion of samples containing crop pests after O. hemprichii. The most prevalent pest found in this species’ diet was O. lybicus. Hypsugo ariel had a very low prevalence of crop pests in its diet. Only 27% of the individuals produced evidence of pest species consumption, and each of those had a single instance of one crop pest per sample.
4. Discussion
4.1. Bat Species Richness and Activity in Organic and Conventional Plantations
Date plantations are frequently used as foraging habitats by most of the bat species present in the area. In total, 12 species were recorded in the three plantations out of 16 species known to be in the Arava valley [29,37], including rare species and threatened species such as B. leucomelas, one of the rarest species in the region, and R. clivosus [52], thus indicating the importance of date plantations as a foraging habitat for desert-dwelling bats.
Bats have equally high activity rates in conventional and organic date plantations. These results are in contrast to our predictions, and also in contrast to results of other studies from temperate regions that found a higher activity of bats on organic farms [53,54]. However, it should be noted that, due to a limited number of organic plantations, Elifaz (conventional) and Samar (organic) plantations were in close proximity, and the bats could easily forage between them. There are stark differences between irrigated agricultural land and the natural arid surroundings that are likely to affect both the abiotic and biotic components of these ecosystems [8]. Artificial green patches in hyper-arid areas, such as date plantations, may act as an artificial oasis, attracting both pests and their natural enemies regardless of their management practices. In contrast to bat activity, species richness is higher at the organic date plantation of Samar compared to the other organic plantation in Neot Smadar and to the conventional plantation of Elifaz. One of the major differences between Samar and the two other plantations is the vegetation cover around the trunk of the palm trees, which does not exist in the other two plantations due to spraying with herbicides or grazing by donkeys and camels [22]. These green patches around the trunks may contribute to the spatial heterogeneity of the date plantation by attracting a higher diversity of insects and increasing bat species richness. Other studies also show that spatial heterogeneity provided through a number of vegetation layers or crop margins with natural vegetation are important factors for determining species richness and activity of bats in different agroecosystems [12,55,56].
Bat activity rates increased during the date harvesting period in all plantations. This might be explained by the appearance of young bats [57] and the life cycle of different moths and other night-active insects, and hence higher prey availability for the bats, although the abundance of the lesser date moth does not change at this time of year [26]. The time of year might also influence bat activity in date plantations, since these artificial oases become one of few green areas in the region in late summer months. Towards the middle of summer, all temporary flood ponds that are filled by flash floods in winter and early spring dry out, and water availability in the desert becomes scarce [58]. The harvesting activities themselves in the plantations may also have an impact on bat activity. Harvest takes place during nighttime due to the high temperatures in the region in August and September. While harvesting, heavy machinery operates in the crown area of the date palms, and entire palms are shaken (Samar) or fruit branches are handled (Neot Smadar and Elifaz). These activities were observed to disturb insects that fly in higher numbers during harvesting activities (personal communication, Schäckermann). During the harvest season, bats are observed foraging next to the machinery and the plantation workers towards the top of the palms (personal communication, Schäckermann). Finally, although we only recorded near adult trees, our model showed that some of the variance in bat activity was partially explained by the recording site. This may indicate that the activity of the bats was also influenced by other spatial variables, such as the nearby youngest plantation, that were not measured in the present study and are known to affect the activity [12]. Detailed habitat assessment study may reveal how spatial features and the plantation structure affect bat activity within the date plantations.
4.2. Bats as Contributors to Natural-Integrated Pest Management
The activity rates and species richness of the bats in the date plantations highlight the potential of insectivorous bats to provide pest control services. All sampled bat species preyed on at least some pest species known to damage the dates. Epetsicus bottae, one of the most common bats identified acoustically in the date plantations, was also one of the species found to consume the widest diversity of crop pests, suggesting that this species may be of particular economic importance to date farmers. Pipistrellus rueppellii may be a control agent against Dubas bug because it commonly consumes this pest. Plecotus christii is a known Lepidopteran specialist [48], but it was detected less often in the date plantations than other species. Because it is a whispering bat species that is very difficult to record with acoustic monitoring methods [37], it might be underrepresented in our dataset and its actual abundance in the plantations might be higher. Hence, its contribution to bio-pest control cannot be determined without further monitoring. Finally, H. ariel, the smallest species in the area [59], consumed the lowest diversity of crop pests. This species primarily consumes small-sized Diptera and Lepidoptera [48], and therefore the crop pests may be too large for it.
The lesser date moth is one of the most severe date pests [24,60], and is the primary pest for dates of the Southern Arava. Some farms report resistance of this pest to the common chemical control methods and are looking for alternatives (personal communication, Schäckermann). We found that three species of bats consumed this pest in date plantations, showing that at least these species are potential bio-control agents against the lesser date moth. Five bat species were found to be potential pest control agents against the Dubas bug, an economically important pest in date plantations [61].
Even though our data reflect that several bats consume the rhinoceros beetle, we believe this not to be the case. The rhinoceros beetle is an economically important date pest against which farmers currently have very little in their arsenal to combat [32,62]. It was identified in the droppings of all bat species in this study, despite its large size precluding it from being consumed by small bats such as H. ariel. Although the DNA was genuinely identified from the samples, we do not think it to be indicative of direct predation by the bats. Instead, this may be a reflection of how common this pest species is in the plantations. The beetles were ubiquitous at agricultural sites and were frequently found tangled in mist nets used to capture bats. Their removal (anecdotally) coated the nets in a reasonable amount of ‘beetle dust’ and was therefore likely coating the bats themselves when they entered the holding bags. Recent research has indicated that DNA is freely available in the environment at detectable levels [63,64]. These findings highlight the potential for mistakes to be made if results are not logically scrutinized following bioinformatics analysis. We conclude that the only likely natural predator of this pest included in our study is O. hemprichii, which is known to consume other large invertebrates (such as scorpions) and hunts by catching prey walking on the ground [49]. Although there is no reliable way to infer actual levels of pest consumption from metabarcoding data [65], it is still reasonable to conclude that much higher read numbers are indicative of true presence, which was true of this pest–species pair.
5. Conclusions
Management of Date Plantations
Our results highlight the potential for a win–win situation in date plantations of the Southern Arava. Farmers can gain from the natural pest control potential that the bats can provide, and the bats can use date plantations as alternative habitats, especially when human development threatens natural habitats or when temporary water sources dry out. The date plantations are located in an area that used to be local floodplains. Seasonal flash floods in the area used to fill up small valley arms once or twice a year in this area, making the desert bloom after the floods subsided. This gave a boost to biodiversity, including a variety of night-active insects, which are the food source for desert-dwelling insectivorous bats. However, the hydrology of the area has changed over recent decades due to development [66,67], and these flood plains have partially disappeared. Hence, date plantations and other agricultural crops provide alternative habitats for desert bats in lieu of lost natural foraging grounds. However, these agricultural habitats do not necessarily include all components that are needed to support bat populations. Food sources are abundantly available in date plantations, since most of the pests are night-active insects; however, they potentially lack in other aspects, such as freely available drinking water sources and roosting sites. Additional landscape structures, including hedgerows and flowering strips, roosting provisions, and creation of water sources could enhance biodiversity and make organic and conventional date plantations more suitable bat habitats.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/d14121034/s1, Table S1: Mist net trapping locations for bats. Agricultural site reflects mist nets set up at locations within the plantations. Adjacent site reflect sites where it was not possible to set up nets within the closest date plantation, hence sites within 1 km radius were chosen. The coordinates in decimal degrees reflect the location of the netting sites.
Author Contributions
Conceptualization, J.S. and C.K.; methodology, C.K., J.S., E.J.M., O.R. and A.A.; validation, J.S.; formal analysis, E.J.M. and C.K.; investigation, C.K., J.S. and E.J.M.; resources J.S., C.K., O.R. and A.A.; data curation, A.A.; writing—original draft preparation, J.S. and C.K.; writing—review and editing, O.R. and E.J.M.; visualization, J.S. and C.K.; supervision, O.R., A.A. and C.K.; project administration, J.S.; funding acquisition, J.S., C.K., O.R. and E.J.M. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by ICA in Israel grant 867, and by KKL-JNF to JS and CK, and by Israel Ministry of Agriculture and Rural Development, # 857071314 (CK). EJM was supported through a GW4+ NERC DTP PhD studentship and EMBO laboratory visit funding 8075. Fieldwork was also supported through a Genetic Society Travel grant and Bat Conservation International grant, both awarded to EJM. Molecular lab work and sequencing were supported through a University of Southampton start-up grant for OR.
Institutional Review Board Statement
This study was approved by the University of Southampton Ethics Committee, and was performed under permit 41282 of CK approved by the Israel Nature and National Parks Protection Authority.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is part of bigger unpublished data sets which will be available once published.
Acknowledgments
We thank the farmers of Samar, Neot Smadar, and Elifaz. Without their help and contribution data collection would not have been possible. Thank you to the staff of the Southern Arava R&D that helped with fieldwork preparation. Thank you to Diana Spinrad for English proof reading. This is publication 1118 of the Mitrani Department of Desert Ecology.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Meyer, W.B.; Turner, B.L. Human Population Growth and Global Land-Use/Cover Change. Annu. Rev. Ecol. Syst. 1992, 23, 39–61. [Google Scholar] [CrossRef]
- Gibbs, H.K.; Ruesch, A.S.; Achard, F.; Clayton, M.K.; Holmgren, P.; Ramankutty, N.; Foley, J.A. Tropical Forests Were the Primary Sources of New Agricultural Land in the 1980s and 1990s. Proc. Natl. Acad. Sci. USA 2010, 107, 16732–16737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United Nations Environment Programme; Middleton, N.; Thomas, D.S.G. World Atlas of Desertification, 2nd ed.; Edward Arnold: London, UK, 1997; ISBN 0340691662. [Google Scholar]
- Blake, G.; Evenari, M.; Shanan, L.; Tadmor, N. The Negev: The Challenge of a Desert; Harvard University Press: Cambridge, CA, USA, 1982. [Google Scholar]
- Fedoroff, N.V.; Battisti, D.S.; Beachy, R.N.; Cooper, P.J.M.; Fischhoff, D.A.; Hodges, C.N.; Knauf, V.C.; Lobell, D.; Mazur, B.J.; Molden, D.; et al. Radically Rethinking Agriculture for the 21st Century. Science 2010, 327, 833–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulata, G.; Simon, Y. Aquaculture in Desert and Arid Lands: Development Constraints and Opportunities; FAO: Rome, Italy, 2010; Volume 6, ISBN 9781479970988. [Google Scholar]
- Tal, A. To Make a Desert Bloom: The Israeli Agricultural Adventure and the Quest for Sustainability. Agric. Hist. 2007, 81, 228–257. [Google Scholar] [CrossRef]
- Mihi, A.; Tarai, N.; Chenchouni, H. Can Palm Date Plantations and Oasification Be Used as a Proxy to Fight Sustainably against Desertification and Sand Encroachment in Hot Drylands? Ecol. Indic. 2019, 105, 365–375. [Google Scholar] [CrossRef]
- Pluess, T.; Opatovsky, I.; Gavish-Regev, E.; Lubin, Y.; Schmidt, M.H. Spiders in Wheat Fields and Semi-Desert in the Negev (Israel). J. Arachnol. 2008, 36, 368–373. [Google Scholar] [CrossRef] [Green Version]
- Segoli, M.; Kishinevsky, M.; Rozenberg, T.; Hoffmann, I. Parasitoid Abundance and Community Composition in Desert Vineyards and Their Adjacent Natural Habitats. Insects 2020, 11, 580. [Google Scholar] [CrossRef]
- Hackett, T.D.; Korine, C.; Holderied, M.W. The Importance of Acacia Trees for Insectivorous Bats and Arthropods in the Arava Desert. PLoS ONE 2013, 8, e52999. [Google Scholar] [CrossRef] [Green Version]
- Kahnonitch, I.; Lubin, Y.; Korine, C. Insectivorous Bats in Semi-Arid Agroecosystems—Effects on Foraging Activity and Implications for Insect Pest Control. Agric. Ecosyst. Environ. 2018, 261, 80–92. [Google Scholar] [CrossRef]
- Boyles, J.G.; Cryan, P.M.; McCracken, G.F.; Kunz, T.H. Economic Importance of Bats in Agriculture. Science 2011, 332, 41–42. [Google Scholar] [CrossRef]
- Williams-Guillén, K.; Olimpi, E.; Maas, B.; Taylor, P.J.; Arlettaz, R. Bats in the Anthropogenic Matrix: Challenges and Opportunities for the Conservation of Chiroptera and Their Ecosystem Services in Agricultural Landscapes. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Springer International Publishing: New York, NY, USA, 2015; pp. 151–186. ISBN 9783319252209. [Google Scholar]
- Maine, J.J.; Boyles, J.G. Bats Initiate Vital Agroecological Interactions in Corn. Proc. Natl. Acad. Sci. USA 2015, 112, 12438–12443. [Google Scholar] [CrossRef] [Green Version]
- Federico, P.; Hallam, T.G.; McCracken, G.F.; Purucker, S.T.; Grant, W.E.; Correa-Sandoval, A.N.; Westbrook, J.K.; Medellín, R.A.; Cleveland, C.J.; Sansone, C.G.; et al. Brazilian Free-Tailed Bats as Insect Pest Regulators in Transgenic and Conventional Cotton Crops. Ecol. Appl. 2008, 18, 826–837. [Google Scholar] [CrossRef]
- Cohen, Y.; Bar-David, S.; Nielsen, M.; Bohmann, K.; Korine, C. An Appetite for Pests: Synanthropic Insectivorous Bats Exploit Cotton Pest Irruptions and Consume Various Deleterious Arthropods. Mol. Ecol. 2020, 29, 1185–1198. [Google Scholar] [CrossRef]
- Maas, B.; Clough, Y.; Tscharntke, T. Bats and Birds Increase Crop Yield in Tropical Agroforestry Landscapes. Ecol. Lett. 2013, 16, 1480–1487. [Google Scholar] [CrossRef]
- Taylor, P.J.; Bohmann, K.; Steyn, J.N.; Schoeman, M.C.; Matamba, E.; Zepeda-Mendoza, M.L.; Nangammbi, T.; Gilbert, M.T.P. Bats Eat Pest Green Vegetable Stink Bugs (Nezara viridula): Diet Analyses of Seven Insectivorous Species of Bats Roosting and Foraging in Macadamia Orchards. In South African Macadamia Growers’ Association Yearbook; Academia: San Francisco, CA, USA, 2013; Volume 21. [Google Scholar]
- Puig-Montserrat, X.; Torre, I.; López-Baucells, A.; Guerrieri, E.; Monti, M.M.; Ràfols-García, R.; Ferrer, X.; Gisbert, D.; Flaquer, C. Pest Control Service Provided by Bats in Mediterranean Rice Paddies: Linking Agroecosystems Structure to Ecological Functions. Mamm. Biol. 2015, 80, 237–245. [Google Scholar] [CrossRef]
- Bruins, H.J.; Sherzer, Z.; Ginat, H.; Batarseh, S. Degradation of Springs in the Arava Valley: Anthropogenic and Climatic Factors. Land Degrad. Dev. 2012, 23, 365–383. [Google Scholar] [CrossRef]
- Abu-Qaoud, H. Date Palm Status and Perspective in Palestine. In Date Palm Genetic Resources and Utilization: Volume 2: Asia and Europe; Springer: Dordrecht, The Netherlands, 2015; pp. 423–439. ISBN 9789401797078. [Google Scholar]
- El-Shafie, H. Review: List of Arthropod Pests and Their Natural Enemies Identified Worldwide on Date Palm, Phoenix dactylifera L. Agric. Biol. J. N. Am. 2012, 3, 516–524. [Google Scholar] [CrossRef]
- Blumberg, D. Date Palm Arthropod Pests and Their Management in Israel. Phytoparasitica 2008, 36, 411–448. [Google Scholar] [CrossRef]
- Al Samarraie, A.I.; Al Hafdh, E.; Abdul Majed, K.; Basumy, M.A. The Chemical Control of the Lesser Date Moth, Batrachedra amydraula Meyr., and Residue Levels of Organophosphate Insecticides in Dates. Pestic. Sci. 1989, 25, 227–230. [Google Scholar] [CrossRef]
- Levi-Zada, A.; Sadowsky, A.; Dobrinin, S.; Ticuchinski, T.; David, M.; Fefer, D.; Dunkelblum, E.; Byers, J.A. Monitoring and Mass-Trapping Methodologies Using Pheromones: The Lesser Date Moth Batrachedra amydraula. Bull. Entomol. Res. 2017, 108, 58–68. [Google Scholar] [CrossRef]
- Mahmoudi, M.; Sahragard, A.; Pezhman, H.; Ghadamyari, M. Efficacy of Biorational Insecticides against Dubas Bug, Ommatissus lybicus (Hem.: Tropiduchidae) in a Date Palm Orchard and Evaluation of Kaolin and Mineral Oil in the Laboratory. J. Entomol. Soc. Iran 2014, 33, 1–10. [Google Scholar]
- GLOBALG.A.P. Available online: https://www.globalgap.org/uk_en/ (accessed on 13 October 2022).
- Yom-Tov, Y.; Kadmon, R. Analysis of the Distribution of Insectivorous Bats in Israel. Divers. Distrib. 1998, 4, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Noy-Meir, I. Desert Ecosystems: Environment and Producers. Annu. Rev. Ecol. Syst. 1973, 4, 25–51. [Google Scholar] [CrossRef]
- Chao, C.C.T.; Krueger, R.R. The Date Palm (Phoenix dactylifera L.): Overview of Biology, Uses, and Cultivation. HortScience 2007, 42, 1077–1082. [Google Scholar] [CrossRef] [Green Version]
- Wakil, W.; Faleiro, J.R.; Miller, T.A. Sustainable Pest Management in Date Palm: Current Status and Emerging Challenges; Springer International Publishing: New York, NY, USA, 2015. [Google Scholar]
- Perring, T.M.; El-Shafie, H.A.F.; Wakil, W. Carob Moth, Lesser Date Moth, and Raisin Moth. In Sustainable Pest Management in Date Palm: Current Status and Emerging Challenges; Springer: Cham, Switzerland, 2015; pp. 109–167. [Google Scholar]
- Blumberg, D. Preliminary Notes on the Phenology and Biology of Batrachedra amydraula Meyrick (Lepidoptera: Cosmopterygidae), a New Pest of Date Palms in Israel. Phytoparasitica 1975, 3, 55–57. [Google Scholar] [CrossRef]
- Fenton, M.B. A Technique for Monitoring Bat Activity with Results Obtained from Different Environments in Southern Ontario. Can. J. Zool. 1970, 48, 847–851. [Google Scholar] [CrossRef]
- Benda, P.; Dietz, C.; Andreas, M.; Hotový, J.; Lučan, R.K.; Malthy, A.; Meakin, K.; Truscott, J.; Vallo, P. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 6. Bats of Sinai (Egypt) with Some Taxonomic, Ecological and Echolocation Data on That Fauna. Acta Soc. Zool. Bohemicae 2008, 72, 103. [Google Scholar]
- Hackett, T.D.; Holderied, M.W.; Korine, C. Echolocation Call Description of 15 Species of Middle-Eastern Desert Dwelling Insectivorous Bats. Bioacoustics 2017, 26, 217–235. [Google Scholar] [CrossRef] [Green Version]
- Alberdi, A.; Aizpurua, O.; Gilbert, M.T.P.; Bohmann, K. Scrutinizing Key Steps for Reliable Metabarcoding of Environmental Samples. Methods Ecol. Evol. 2018, 9, 134–147. [Google Scholar] [CrossRef] [Green Version]
- Corse, E.; Tougard, C.; Archambaud-Suard, G.; Agnèse, J.F.; Messu Mandeng, F.D.; Bilong Bilong, C.F.; Duneau, D.; Zinger, L.; Chappaz, R.; Xu, C.C.Y.; et al. One-Locus-Several-Primers: A Strategy to Improve the Taxonomic and Haplotypic Coverage in Diet Metabarcoding Studies. Ecol. Evol. 2019, 9, 4603–4620. [Google Scholar] [CrossRef]
- Zeale, M.R.K.; Butlin, R.K.; Barker, G.L.A.; Lees, D.C.; Jones, G. Taxon-Specific PCR for DNA Barcoding Arthropod Prey in Bat Faeces. Mol. Ecol. Resour. 2011, 11, 236–244. [Google Scholar] [CrossRef]
- Jusino, M.A.; Banik, M.T.; Palmer, J.M.; Wray, A.K.; Xiao, L.; Pelton, E.; Barber, J.R.; Kawahara, A.Y.; Gratton, C.; Peery, M.Z.; et al. An Improved Method for Utilizing High-Throughput Amplicon Sequencing to Determine the Diets of Insectivorous Animals. Mol. Ecol. Resour. 2019, 19, 176–190. [Google Scholar] [CrossRef]
- Murray, D.C.; Coghlan, M.L.; Bunce, M. From Benchtop to Desktop: Important Considerations When Designing Amplicon Sequencing Workflows. PLoS ONE 2015, 10, e0124671. [Google Scholar] [CrossRef] [Green Version]
- Schnell, I.B.; Bohmann, K.; Gilbert, M.T.P. Tag Jumps Illuminated—Reducing Sequence-to-Sample Misidentifications in Metabarcoding Studies. Mol. Ecol. Resour. 2015, 15, 1289–1303. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet.journal 2011, 17, 10. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [Green Version]
- Ratnasingham, S. MBRAVE: The Multiplex Barcode Research And Visualization Environment. Biodivers. Inf. Sci. Stand. 2019, 3, e37986. [Google Scholar] [CrossRef]
- Feldman, R.; John, O.W.; Yom-Tov, Y. Dietary Composition and Habitat Use in a Desert Insectivorous Bat Community in Israel. Acta Chiropterol. 2000, 2, 15–22. [Google Scholar]
- Holderied, M.; Korine, C.; Moritz, T. Hemprich’s Long-Eared Bat (Otonycteris hemprichii) as a Predator of Scorpions: Whispering Echolocation, Passive Gleaning and Prey Selection. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2011, 197, 425–433. [Google Scholar] [CrossRef]
- Yomtov, Y. Character displacement among the insectivorous bats of the Dead-Sea area. J. Zool. 1993, 230, 347–356. [Google Scholar]
- Korine, C.; Pinshow, B. Guild structure, foraging space use, and distribution in a community of insectivorous bats in the Negev Desert. J. Zool. 2004, 262, 187–196. [Google Scholar] [CrossRef]
- Dolev, A.; Perevolotsky, A. Vertebrates in Israel: The red book; Israel Nature and Parks Authority: Jerusalem, Israel, 2004. [Google Scholar]
- Puig-Montserrat, X.; Mas, M.; Flaquer, C.; Tuneu-Corral, C.; López-Baucells, A. Benefits of Organic Olive Farming for the Conservation of Gleaning Bats. Agric. Ecosyst. Environ. 2021, 313, 107361. [Google Scholar] [CrossRef]
- Wickramasinghe, L.P.; Harris, S.; Jones, G.; Vaughan, N. Bat Activity and Species Richness on Organic and Conventional Farms: Impact of Agricultural Intensification. J. Appl. Ecol. 2003, 40, 984–993. [Google Scholar] [CrossRef]
- Estrada, C.G.; Damon, A.; Hernández, C.S.; Pinto, L.S.; Núñez, G.I. Bat Diversity in Montane Rainforest and Shaded Coffee under Different Management Regimes in Southeastern Chiapas, Mexico. Biol. Conserv. 2006, 132, 351–361. [Google Scholar] [CrossRef]
- Kelly, R.M.; Kitzes, J.; Wilson, H.; Merenlender, A. Habitat Diversity Promotes Bat Activity in a Vineyard Landscape. Agric. Ecosyst. Environ. 2016, 223, 175–181. [Google Scholar] [CrossRef]
- Mendelssohn, H.; Yom-Tov, Y. Jerusalem: Israel Academy of Sciences and Humanities. In Fauna Palaestina Mammalia of Israel; Keter Press: Jerusalem, Israel, 1999. [Google Scholar]
- Korine, C.; Adams, R.; Russo, D.; Fisher-Phelps, M.; Jacobs, D. Bats and Water: Anthropogenic Alterations Threaten Global Bat Populations. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Springer International Publishing: New York, NY, USA, 2015; pp. 215–241. ISBN 9783319252209. [Google Scholar]
- Yom-Tov, Y.; Makin, D.; Shalmon, B. The biology of Pipistrellus bodenheimeri (Microchiroptera) in the Dead Sea area of Israel. Z. Säugetierkunde 1992, 57, 65–69. [Google Scholar]
- Kakar, M.K.; Nizamani, S.M.; Rustamani, M.A.; Khuhro, R.D. Periodical Lesser Date Moth Infestation on Intact and Dropped Fruits. Sarhad J. Agric. 2010, 26, 393–396. [Google Scholar]
- Bagheri, A.; Fathipour, Y.; Askari-Seyahooei, M.; Zeinalabedini, M. Ommatissus lybicus (Hemiptera: Tropiduchidae), an Economically Important Pest of Date Palm (Arecaceae) with Highly Divergent Populations. Can. Entomol. 2018, 150, 378–392. [Google Scholar] [CrossRef]
- Soltani, R. The Rhinoceros Beetle Oryctes agamemnon arabicus in Tunisia: Current Challenge and Future Management Perspectives. Tunis. J. Plant Prot. 2010, 5, 179–193. [Google Scholar]
- Clare, E.L.; Economou, C.K.; Bennett, F.J.; Dyer, C.E.; Adams, K.; McRobie, B.; Drinkwater, R.; Littlefair, J.E. Measuring Biodiversity from DNA in the Air. Curr. Biol. 2022, 32, 693–700.e5. [Google Scholar] [CrossRef]
- Lynggaard, C.; Bertelsen, M.F.; Jensen, C.V.; Johnson, M.S.; Frøslev, T.G.; Olsen, M.T.; Bohmann, K. Airborne Environmental DNA for Terrestrial Vertebrate Community Monitoring. Curr. Biol. 2022, 32, 701–707.e5. [Google Scholar] [CrossRef]
- Alberdi, A.; Aizpurua, O.; Bohmann, K.; Gopalakrishnan, S.; Lynggaard, C.; Nielsen, M.; Gilbert, M.T.P. Promises and Pitfalls of Using High-Throughput Sequencing for Diet Analysis. Mol. Ecol. Resour. 2019, 19, 327–348. [Google Scholar] [CrossRef]
- Armoza-Zvuloni, R.; Shlomi, Y.; Shem-Tov, R.; Stavi, I.; Abadi, I. Drought and Anthropogenic Effects on Acacia Populations: A Case Study from the Hyper-Arid Southern Israel. Soil Syst. 2021, 5, 23. [Google Scholar] [CrossRef]
- Armoza-Zvuloni, R.; Shlomi, Y.; Abadi, I.; Shem-Tov, R.; Laronne, J.B. Fluvial Sediment Yields in Hyper-Arid Areas, Exemplified by Nahal Nehushtan, Israel. Land 2022, 11, 1050. [Google Scholar] [CrossRef]
Figure 1.
(A) Overview map of the South of Israel indicating the location of the research area displayed with a red rectangle. (B) Location of the three date plantations in the Southern Arava marked with different colorful icons.
Figure 1.
(A) Overview map of the South of Israel indicating the location of the research area displayed with a red rectangle. (B) Location of the three date plantations in the Southern Arava marked with different colorful icons.

Figure 2.
Bat species richness (A) and bat activity rates (B) before date harvesting, and bat species richness (C) and bat activity rates (D) during date harvesting. Recordings were made with bat detectors in the three date plantations of Elifaz (conventional), Neot Smadar (organic no ground cover), Samar (organic with intensive ground cover) in June 2018 before harvest time (63 nights) and in September 2018 during harvest time (66 nights) of the dates. Results are presented as average ± S.D. Asterisk indicates significant differences.
Figure 2.
Bat species richness (A) and bat activity rates (B) before date harvesting, and bat species richness (C) and bat activity rates (D) during date harvesting. Recordings were made with bat detectors in the three date plantations of Elifaz (conventional), Neot Smadar (organic no ground cover), Samar (organic with intensive ground cover) in June 2018 before harvest time (63 nights) and in September 2018 during harvest time (66 nights) of the dates. Results are presented as average ± S.D. Asterisk indicates significant differences.


{kind=link}
{kind=link}
Table 1.
Total bat passes per species and their foraging mode in three date plantations located at the Southern Arava valley, Israel. Neot Smadar and Samar are organic plantations, while Elifaz is a conventional date plantation. See Methods for more details on the differences in management between the three plantations. Foraging mode of the bats species is based on [11,38,48,49,50,51].
Table 1.
Total bat passes per species and their foraging mode in three date plantations located at the Southern Arava valley, Israel. Neot Smadar and Samar are organic plantations, while Elifaz is a conventional date plantation. See Methods for more details on the differences in management between the three plantations. Foraging mode of the bats species is based on [11,38,48,49,50,51].
| Location | Elifaz | Neot Smadar | Samar | Foraging Mode | |||
|---|---|---|---|---|---|---|---|
| Bat Species | Before Harvest | During Harvest | Before Harvest | During Harvest | Before Harvest | During Harvest | |
| Eptesicus bottae | 5334 | 3094 | 6347 | 5023 | 3931 | 5414 | Cultured space— Ariel hawking |
| Hypsugo ariel | 2121 | 11,649 | 3799 | 7120 | 3503 | 6600 | Cultured space— Ariel hawking |
| Taphozous nudiventris | 438 | 1966 | 493 | 1608 | 1431 | 909 | Open space— Ariel hawking |
| Rhinopoma cystops | 190 | 612 | 613 | 145 | 198 | 541 | Open space— Ariel hawking |
| Rhinopoma microphyllum | 0 | 176 | 0 | 10 | 0 | 206 | Open space— Ariel hawking |
| Pipistrellus kuhlii | 133 | 1387 | 78 | 395 | 699 | 3972 | Cultured space— Ariel hawking |
| Otonycteris hemprichii | 135 | 91 | 4 | 48 | 116 | 433 | Gleaners |
| Pipistrellus rueppellii | 86 | 614 | 182 | 55 | 286 | 2763 | Cultured space— Ariel hawking |
| Barbastella leucomelas | 61 | 353 | 77 | 174 | 137 | 300 | Cultured space— Ariel hawking |
| Plecotus christii | 55 | 19 | 13 | 0 | 55 | 159 | Gleaners |
| Rhinolophus clivosus | 0 | 0 | 64 | 12 | 14 | 154 | Highly cluttered space |
| Tadarida teniotis | 0 | 64 | 3 | 247 | 20 | 517 | Open space— Ariel hawking |
| Unknown | 27 | 41 | 21 | 57 | 39 | 51 | |
| Sub Total | 8580 | 20,066 | 11,694 | 14,894 | 10,429 | 22,019 | |
| Total | 87,862 | ||||||
Table 2.
Average species richness and total bat activity ± SD before and during the harvest of the dates in three date plantations in the Arava valley, Israel. Neot Smadar and Samar are organic plantations, while Elifaz is a conventional date plantation.
Table 2.
Average species richness and total bat activity ± SD before and during the harvest of the dates in three date plantations in the Arava valley, Israel. Neot Smadar and Samar are organic plantations, while Elifaz is a conventional date plantation.
| Date Plantations | Before Harvest | During Harvest | t-Test | p-Value |
|---|---|---|---|---|
| Elifaz | ||||
| Species richness | 7.2 ± 0.2 | 9.1 ± 0.7 | 5.8 | 0.001 |
| Total bat activity (passes/night) | 427.2 ± 322.7 | 960.9 ± 436.1 | 2.1 | 0.07 |
| Neot Smadar | ||||
| Species richness | 6.6 ± 0.4 | 8.21 ± 1.3 | 2.7 | 0.04 |
| Total bat activity (passes/night) | 538.4 ± 286.6 | 946.6 ± 760.1 | 1.1 | 0.3 |
| Samar | ||||
| Species richness | 8.32 ± 1.2 | 10.1 ± 0.6 | 2.8 | 0.03 |
| Total bat activity (passes/night) | 480.8 ± 176.2 | 962.6 ± 722.5 | 1.2 | 0.3 |
Table 3.
Pests found in fecal samples of desert bats in the Arava valley, Israel through DNA metabarcoding. N is for the number of samples containing the pest species in each studied bat species.
Table 3.
Pests found in fecal samples of desert bats in the Arava valley, Israel through DNA metabarcoding. N is for the number of samples containing the pest species in each studied bat species.
| Pest Species | Pest Order | Eptesicus bottae (N = 25) | Hypsugo ariel (N = 11) | Otonycteris hemprichii (N = 16) | Pipistrellus kuhlii (N = 8) | Pipistrellus rueppellii (N = 11) | Plecotus christii (N = 8) |
|---|---|---|---|---|---|---|---|
| Apomyelois ceratoniae/Ectomyelois ceratoniae | Lepidoptera | 1 | 0 | 0 | 0 | 0 | 1 |
| Arenipses sabella | Lepidoptera | 1 | 0 | 0 | 0 | 0 | 1 |
| Batrachedra amydraula | Lepidoptera | 2 | 0 | 2 | 1 | 0 | 0 |
| Cadra figulilella | Lepidoptera | 1 | 0 | 1 | 0 | 1 | 3 |
| Carpophilus hemipterus | Coleoptera | 1 | 1 | 2 | 0 | 1 | 0 |
| Carpophilus mutilatus | Coleoptera | 0 | 0 | 1 | 0 | 1 | 0 |
| Epuraea luteolus | Coleoptera | 0 | 0 | 1 | 0 | 0 | 0 |
| Ommatissus lybicus | Hemiptera | 3 | 1 | 1 | 1 | 3 | 0 |
| Oryctes agamemnon | Coleoptera | 7 | 1 | 11 | 2 | 3 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Schäckermann, J.; Morris, E.J.; Alberdi, A.; Razgour, O.; Korine, C. The Contribution of Desert-Dwelling Bats to Pest Control in Hyper-Arid Date Agriculture. Diversity 2022, 14, 1034. https://0-doi-org.brum.beds.ac.uk/10.3390/d14121034
AMA Style
Schäckermann J, Morris EJ, Alberdi A, Razgour O, Korine C. The Contribution of Desert-Dwelling Bats to Pest Control in Hyper-Arid Date Agriculture. Diversity. 2022; 14(12):1034. https://0-doi-org.brum.beds.ac.uk/10.3390/d14121034
Chicago/Turabian StyleSchäckermann, Jessica, Evie J. Morris, Antton Alberdi, Orly Razgour, and Carmi Korine. 2022. "The Contribution of Desert-Dwelling Bats to Pest Control in Hyper-Arid Date Agriculture" Diversity 14, no. 12: 1034. https://0-doi-org.brum.beds.ac.uk/10.3390/d14121034
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.