1. Introduction
Juniperus sabina L. (Cupressaceae) is a dioecious coniferous evergreen shrub with both sexual and asexual reproduction strategies.
J. sabina (savin juniper) covers a vast area of the northern hemisphere, but its distribution is fragmented across Eurasia, from Spain in the west to Mongolia and Siberia in the east. The World Flora (WFO) database recognises five varieties within this species: var.
sabina, var.
mongoliensis R.P. Adams, var.
davurica (Pall.) Farjon, var.
arenaria (E.H.Wilson) Farjon and var.
balkanesis R.P. Adams & Tashev [
1]. Due to its ability to spontaneously produce adventitious roots on soil covered branches and its resistance to pruning, it plays a significant role in the prevention of land desertification and the improvement of the urban landscape [
2].
Balkan savin juniper is a just recently described variety [
3]. It is an extremely rare variety that grows only in the Balkans. It can only be found in very inaccessible terrains, growing mostly on screes and vertical cliffs. The most recent research shows that it is common in the Balkans and Anatolia region [
4,
5,
6,
7,
8,
9]; however, there is no data for Serbia, where its central part of distribution should be found. While Adams found slight morphological differences (e.g., in the colour and texture of leaves, position of leaf glands and shape of seed cones), the most distinct characteristics are found in their genome, reflecting an ancient hybridisation event with
J. thurifera [
3]. This was also confirmed by Farhat et al. [
4,
10,
11], who discovered that all of the var.
balkanensis individuals had twice the amount of DNA than other varieties of this species, suggesting that the variety is tetraploid, rather than diploid.
In addition to its significance in natural and urban environments, savin juniper produces large amounts of bioactive compounds, mainly terpenoids; therefore, their essential oil composition has largely been studied from this aspect [
8,
12,
13,
14,
15,
16,
17]. However, there are not many reports dealing with the intra- and inter-population variability of their essential oil. Essential oils (EOs) are mixtures of volatile organic compounds, mostly commonly terpenoids. Monoterpens and sesquiterpenes dominate juniper essential oils, in which up to 100 different terpene and non-terpene compounds could be found. In addition to their bioactivity, conifer essential oils, and particularly juniper essential oils, have been studied for their chemophenetic significance as chemophenetic markers that have proven useful in describing and assessing the genetic diversity of populations, as their composition is mostly genetically determined [
18,
19,
20,
21,
22]. Present literature data on savin juniper shows the domination of two biosynthetically linked monoterpenes—sabinene and
trans-sabinyl acetate (
Figure 1); however, not all enzymes involved in the biosynthesis are known [
23,
24,
25,
26]. Nevertheless, it appears that sabinene is the starting compound for several highly abundant savin juniper terpenes.
While the species has been proven as hardy and is widely grown for ornamental purposes, it cannot withstand competition with other shrubs or trees in its natural habitat. As these remaining fragments of natural populations are important gene pools, it is important to preserve their genetic resources and reconstruct or find new habitats for this species. Thus, the starting point in these efforts is determining the genetic diversity and structure of natural populations of
J. sabina var.
balkanensis. However, previous investigations of the population variability using molecular markers have only been conducted in Asia [
2], where two genetic pools were discovered. Recently, different authors have suggested using inter-simple sequence repeat (ISSR) markers as an alternative system with reliability and advantages over other molecular marker systems [
27,
28,
29,
30]. This technique involves the amplification of genomic segments flanked by inversely oriented microsatellite sequences. These regions are amplified using primers based on SSRs anchored with a couple of nucleotides. The resulting PCR products are multiple bands of different lengths that could be coded into a presence/absence matrix. ISSR markers are dominant markers that show Mendelian inheritance.
Protecting this narrowly distributed taxon and its genetic potential is of paramount significance in studies of changing climate on its growth and distribution. The aim of this study was to establish intra- and inter-population variability using leaf essential oil composition and their possible correlation with environmental factors. Individuals with different chemotypes were chosen for molecular analyses. ISSR were used to assess intra- and inter-population variability and correspondence with phytochemical markers, while chloroplast trnG-trnS sequences were used to further confirm that the samples are J. sabina var. balkanensis.
4. Discussion
Analysis of EOs from 100 individuals from seven populations showed high variability in the EO composition between individuals from the same population that grow only a few meters apart, which corroborates the chemophenetic significance of EOs. This also suggests that the environment does not strongy influence EO composition, as different chemotypes could be detected on the same substratum, growing under the same conditions. Statistical analysis confirmed strong and statistically significant negative correlation of sabinene and
trans-sabinyl acetate, thus confirming the hypothesis of their biosynthethic connection. Three chemotypes were detected in the studied populations. Our data correspond to the available literature data (
Table 6). Previous reports show that sabinene was the dominant compound in most of the populations. However,
trans-sabynil acetate was dominant in the populations from Canada, Switzerland, and Iran. Only in one sample, from Bulgaria, was myrtenyl acetate the dominant compound [
9]. Our population from Bulgaria was very similar to those EOs obtained by Zheljazkov et al. [
9]. Aside from the dominant compounds, the overall relatively high variability of the EO corresponds to the literature data.
In the present analysis, three chemotypes were more-or-less equally distributed in the sample; however, the distribution of chemotypes differed between populations. In the populations from Mt. Bistra and Mt. Biokovo,
trans-sabinyl acetate chemotype was not detected. While in the case of Mt. Biokovo, this could be due to a sampling artefact (
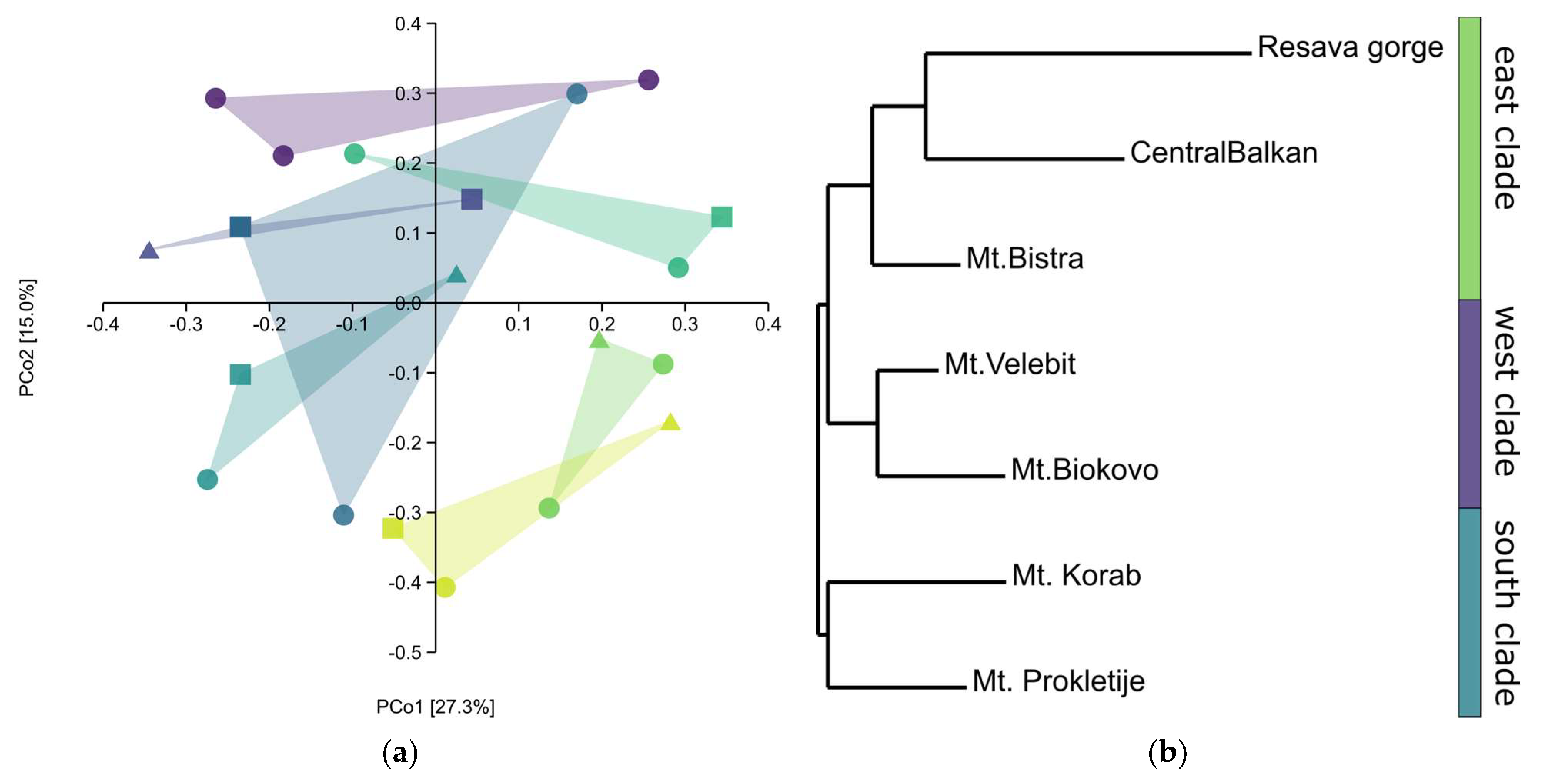
n = 5), this cannot be the case with the Mt. Bistra population, particularly considering the almost equal distribution of chemotypes in the whole sample. Additionally, ISSRs show that Mt. Bistra has more-or-less the same genetic diversity as most of other populations (
Figure 5a). In the model, a single gene is crucial for the transformation of sabinene into
trans-sabinyl acetate; it follows classic Mendelian dominant/recessive inheritance that the amount of these compounds would correspond to the number of alleles present in the genotype. However, isolation of the enzymes responsible for biosynthesis and the identification of the genes are necessary to confirm this. Previous reports on
Juniperus deltoides,
J. communis var.
communis and
J. communis var.
saxatilis show different distributions of chemotypes across the Balkans [
18,
19,
21,
22]. In
J. deltoides, the limonene chemotype was more common in coastal populations, while
α-pinene was more common in the continental populations [
21]. A similar pattern was observed in the two varieties of
J. communis where five different chemotypes were detected [
18,
19]. In.
J. communis var.
communis, several chemotypes were detected in the literature [
19,
51,
52,
53,
54], which showed different abundances related to the geographic distribution. While the distribution range of
J. communis var.
communis and
J. deltoides is somewhat continuous in this region,
J. communis var.
saxatilis has a more fragmented distribution, as it grows above the tree line in high mountains. Nevertheless, in this taxon, the geographic distribution of chemotypes was also apparent, where
δ-3-carene chemotype was most common in the areas with greater precipitation, and
α-pinene and sabinene were more frequent in the more arid areas of the Balkans. In this research, the sabinene chemotype was more common in the areas with lower temperatures [
18]. On the other hand, in more closely related species (
J. phoenicea var.
turbinata), EO was strongly dominated by a single compound (
α-pinene), although several other compounds were present in higher concentrations (e.g.,
δ-3-carene, caryophyllene oxide, germacrene D, limonene and myrcene). No apparent pattern was noticed in the geographic distribution [20 and refs. cited therein].
The detected genetic variability was on par with the available literature data. To the best of our knowledge, ISSRs have never previously been used in
J. sabina. However, Boogat et Salehi [
28] used 12 ISSR primers to analyse five populations of
Juniperus polycarpos, obtaining PIC values between 0.38 and 0.5, which was higher than our results. Since ISSRs are dominant genetic markers, their highest PIC value cannot be greater than 0.5; therefore, the obtained PIC values suggest that selected ISSRs were moderately informative in our study.
J. sabina genetic diversity was previously studied using SSRs and RAPDs [
2,
55]. For SSRs, authors obtained PIC values between 0.179 and 0.751. ISSRs were used to study the genetic diversity and differentiation of populations of
J. polycarpos,
J. brevifolia,
J. phoenicea, J. chinenesis, and
J. excelsa [
28,
30,
56,
57,
58]. In all of these studies, ISSRs showed a high number of polymorphic bands. Most of the studies of different junipers, however, relied on the SSRs [
2,
56,
59,
60,
61,
62,
63,
64] or AFLPs [
18,
65,
66,
67,
68,
69], which have shown to have high correlation with the ISSRs [
56]. Furthermore, studies performed by several authors showed high correlation of SSRs and ISSRs with other genetic markers [
58,
60,
70].
Juniperus sabina from Asia showed that 88% of variability lies within populations, while only 12% exists between populations [
2]. These results are somewhat lower than ours, particularly when taking the area of study into consideration. Bettencourt et al. [
56] showed that ISSRs tend to have a lower population variability than SSRs. Similar results of genetic diversity were obtained for other junipers, where intrapopulation variability accounted of between 50.9% in
J. communis [
69] and 98% in
J. oxycedrus [
71], depending on both the geographic area analysed and the species. However, in most cases, the population variability was around 90% [
18,
56,
59,
61,
62,
63,
66,
68]. Conifers, and juniper among them, tend to show high genetic diversity within populations, and low separation of populations and regions. This can be explained by the biology of conifers. Junipers are long living species, the seeds of which are dispersed by birds, and the pollen is carried by wind. As one can see from the widely available data, genetic distances between populations in junipers are usually low, even when the distribution is fragmented and populations live at great distances.
Both ISSRs and EOs showed high intrapopulation variability with moderate separation of populations. Additionally, both sets of markers did not show a correlation with the geographic region, according to the Mantel test. However, the grouping of the populations differed widely, and no correlation was found between genetic and phenotypic markers. Molecular markers showed that populations from the same mountain ranges were grouped closely together. The only exception was the population from Mt. Bistra, which grouped with the east clade, rather than the south. This could be due to the different origins of these two populations, with the population from Mt. Bistra coming from the east, rather than from Albania and Montenegro, but this could also be due to the low sample size in the present study and the high genetic diversity within this taxon. Additionally, the present genetic pattern could also be explained by the distribution of this taxon during Pleistocene. Savin juniper grows in very arid areas, where there is no competition from other trees and woody species. As pollen cannot be used in the reconstruction of savin juniper’s historic distribution, a precise distribution range cannot be reconstructed. Nevertheless, based on the biology of the species, one can assume that the areas suitable for this taxon were more abundant during the Pleistocene in the Balkans. The genetic data suggest the possibility of two refugia, although further studies are necessary to test this. The EO composition showed different groups than the molecular data, which were not geographically correlated. For example, populations from Mt. Biokovo and Mt. Bistra were grouped close to each other, as well as the populations from Mt. Velebit and Resavska gorge. The chemical composition of the leaf essential oils and the distribution of the chemotypes were not correlated with the environmental data and geological substratum, suggesting that other ecological factors might be at play, e.g., selection due to herbivory. The high genetic diversity detected in the studied populations suggests that the sexual reproduction is more prevalent than the vegetative, with the exception of the population from Resavska gorge, which had the lowest genetic diversity of the studied populations, indicating that this population might have gone through a genetic bottleneck.

 ,
, 


 —Central Balkan,
—Central Balkan,  —Resava gorge,
—Resava gorge,  —Mt. Bistra,
—Mt. Bistra,  —Mt. Korab,
—Mt. Korab,  —Mt. Prokletije,
—Mt. Prokletije,  —Mt. Biokovo,
—Mt. Biokovo,  —Mt. Velebit. Principle components given in the left-hand side corner: S—Sabinene, SA—trans-Sabinyl acetate, So—trans-Sabinol, T—terpinen-4-ol, A—Abieta-7,13-dien-3-one.
—Mt. Velebit. Principle components given in the left-hand side corner: S—Sabinene, SA—trans-Sabinyl acetate, So—trans-Sabinol, T—terpinen-4-ol, A—Abieta-7,13-dien-3-one.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}