

Diversity Patterns of Late Jurassic Chondrichthyans: New Insights from a Historically Collected Hybodontiform Tooth Assemblage from Poland


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fossil Material in This Study
2.2. Methods
3. Results
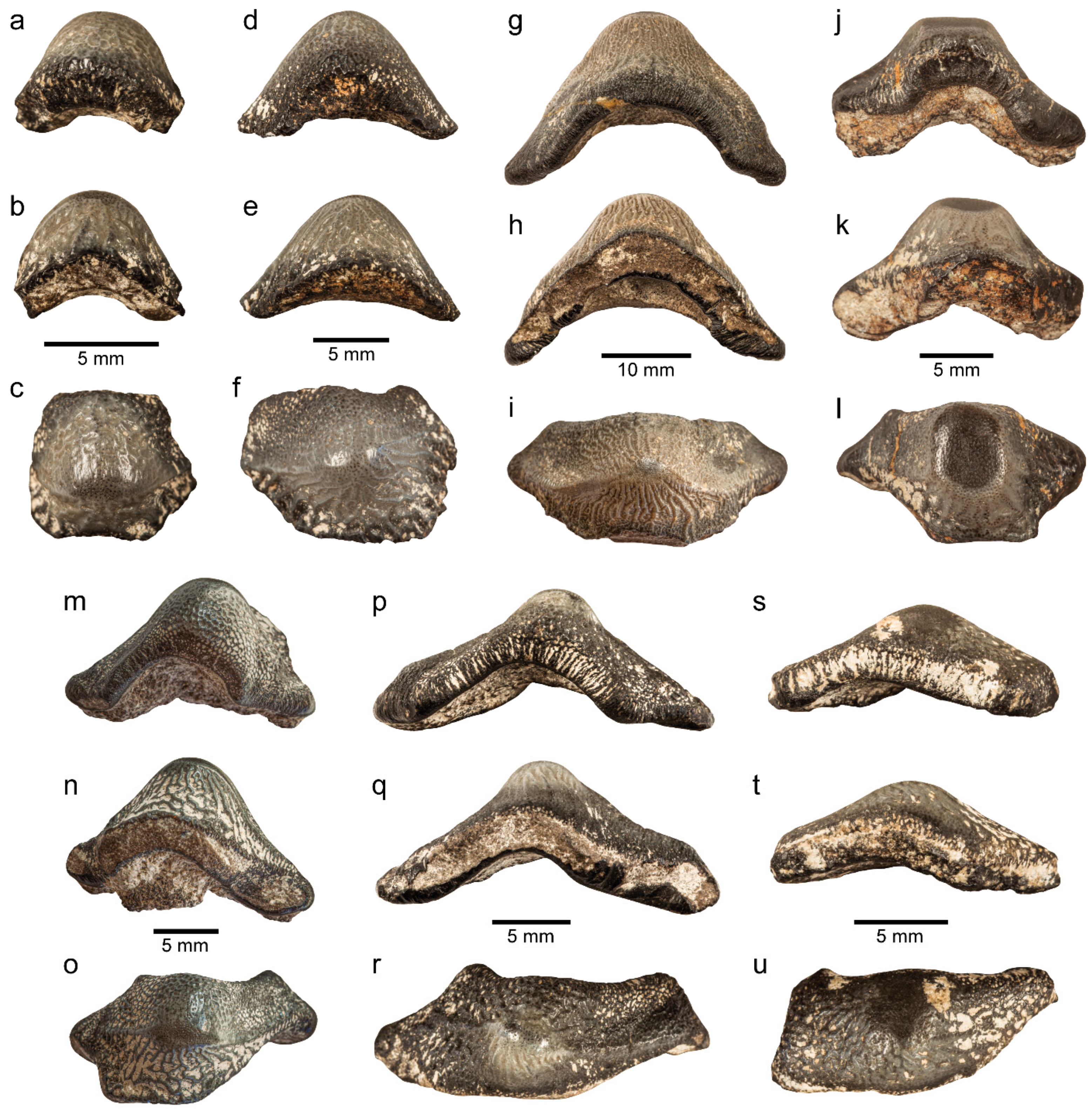
3.1. Systematic Palaeontology
3.1.1. Material
3.1.2. Description
3.1.3. Remarks
3.1.4. Material
3.1.5. Description
3.1.6. Remarks
3.1.7. Material
3.1.8. Description
3.1.9. Remarks
3.1.10. Material
3.1.11. Description
3.1.12. Remarks
4. Discussion
4.1. Palaeoecology
4.2. Late Jurassic Hybodontiform Diversity and Distribution Patterns
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tennant, J.P.; Mannion, P.D.; Upchurch, P.; Sutton, M.D.; Price, G.D. Biotic and environmental dynamics through the Late Jurassic–Early Cretaceous transition: Evidence for protracted faunal and ecological turnover. Biol. Rev. 2017, 92, 776–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upchurch, P.; Mannion, P.D.; Benson, R.B.J.; Butler, R.J.; Carrano, M.T. Geological and anthropogenic controls on the sampling of the terrestrial fossils record: A case study from the Dinosauria. In Comparing the Geological and Fossil Records: Implications for Biodiversity Studies; McGowan, A.J., Smith, A.B., Eds.; Special Publications 358; The Geological Society: London, UK, 2011; pp. 209–240. [Google Scholar] [CrossRef]
- Friedman, M.; Sallan, L.C. Five hundred million years of extinction and recovery: A phanerozoic survey of large-scale diversity patterns in fishes. Palaeontology 2012, 55, 707–742. [Google Scholar] [CrossRef]
- Benson, R.B.J.; Druckenmiller, P.S. Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition. Biol. Rev. 2014, 89, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Guinot, G.; Cavin, L. ‘Fish’ (Actinopterygii and Elasmobranchii) diversification patterns through deep time. Biol. Rev. 2016, 91, 950–981. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, T.L.; Benton, M.J. Ecomorphological diversifications of Mesozoic marine reptiles: The roles of ecological opportunity and extinction. Paleobiology 2016, 42, 547–573. [Google Scholar] [CrossRef] [Green Version]
- Tennant, J.P.; Mannion, P.D.; Upchurch, P. Sea level regulated tetrapod diversity dynamics through the Jurassic/Cretaceous interval. Nat. Commun. 2016, 7, 12737. [Google Scholar] [CrossRef] [Green Version]
- Compagno, L.J.V. Interrelationships of living elasmobranchs. Zool. J. Linn. Soc. 1973, 53, 15–61. [Google Scholar]
- Cappetta, H.; Buffetaut, E.; Cuny, G.; Suteethorn, V. A new elasmobranch assemblage from the Lower Cretaceous of Thailand. Palaeontology 2006, 49, 547–555. [Google Scholar] [CrossRef]
- Cuny, G.; Ouaja, M.; Srarfi, D.; Schmitz, L.; Buffetaut, E.; Benton, M.J. Fossil sharks from the Early Cretaceous of Tunisia. Rev. Paléobiol. 2004, 9, 127–142. [Google Scholar]
- Cuny, G.; Suteethorn, V.; Khamha, S.; Buffetaut, E.; Philippe, M. A new hybodont shark assemblage from the Lower Cretaceous of Thailand. Hist. Biol. 2006, 18, 21–31. [Google Scholar] [CrossRef]
- Cuny, G.; Suteethorn, V.; Khamha, S.; Buffetaut, E. Hybodont sharks from the Lower Cretaceous Khok Kruat Formation of Thailand, and hybodont diversity during the Early Cretaceous. In Fishes and the Break-up of Pangaea; Cavin, L., Longbottom, A., Richter, M., Eds.; Special Publications 295; The Geological Society: London, UK, 2008; pp. 93–107. [Google Scholar] [CrossRef]
- Cuny, G.; Cavin, L.; Suteethorn, V. A new hybodont with a cutting dentition from the Lower Cretaceous of Thailand. Cret. Res. 2009, 30, 515–520. [Google Scholar] [CrossRef]
- Cuny, G.; Laojumpon, C.; Cheychiw, O.; Lauprasert, K. Fossil vertebrate remains from Kut Island (Gulf of Thai land, Early Cretaceous). Cret. Res. 2010, 31, 415–423. [Google Scholar] [CrossRef]
- Cuny, G. Freshwater hybodont sharks in Early Cretaceous ecosystems: A review. In Bernissart Dinosaurs and Early Cretaceous Terrestrial Ecosystems; Godefroit, P., Ed.; Indiana University Press: Bloomington, IN, USA, 2012; pp. 518–529. [Google Scholar]
- Cappetta, H. Chondrichthyes: Mesozoic and Cenozoic Elasmobranchii. In Handbook of Paleoichthyology, 3E; Verlag Dr. Friedrich Pfeil: München, Germany, 2012; p. 512. [Google Scholar]
- Fricke, K. Die fossilen Fische aus den oberen Juraschichten von Hannover. Palaeontographica 1876, 22, 347–398. [Google Scholar]
- Woodward, A.S. Catalogue of the Fossil Fishes in the British Museum (Natural History); Part I; British Museum: London, UK, 1889; p. 474. [Google Scholar]
- Gallinek, E. Der obere Jura bei Inowrazlaw in Posen. Verh. Kais. Russ.-Miner. Ges. St. Petersbg. 1895, 33, 353–427. [Google Scholar]
- Candoni, L. Deux faunes inédites de sélaciens dans le Jurassique terminal Français: Premiers résultats stratigraphiques. Bull. Tr. Soc. Géol. Norm. Amis Muséum Havre 1995, 82, 29–49. [Google Scholar]
- Kriwet, J. Beitrag zur Kenntnis der Fisch-Fauna des Ober-Jura (unteres Kimmeridge) der Kohlengrube Guimarota bei Leiria, Mittel-Portugal: 1. Asteracanthus biformatus n. sp. (Chondrichthyes: Hybodontoidea). Berl. Geowiss. Abh. 1995, E16, 683–691. [Google Scholar]
- Kriwet, J. Late Jurassic elasmobranch and actinopterygian fishes from Portugal and Spain. Cuad. Geol. Ibér. 1998, 24, 241–260. [Google Scholar]
- Kriwet, J. Late Jurassic selachians (Chondrichthyes: Hybodontiformes, Neoselachii) from Central Portugal. N. Jb. Geol. Paläontol. Mh. 2004, 4, 233–256. [Google Scholar] [CrossRef]
- Duffin, C.J.; Thies, D. Hybodont shark teeth from the Kimmeridgian (Late Jurassic) of northwest Germany. Geol. Palaeontol. 1997, 31, 235–256. [Google Scholar]
- Underwood, C.J. Sharks, rays and a chimaeroid from the Kimmeridgian (Late Jurassic) of Ringstead, southern England. Palaeontology 2002, 45, 297–325. [Google Scholar] [CrossRef] [Green Version]
- Underwood, C.J. 2. Sharks and rays. In Fossils of the Kimmeridge Clay Formation. Vol. 2: Vertebrate Palaeontology; Martill, D.M., Etches, S., Eds.; Field Guide to Fossils 16; Palaeontological Association: London, UK, 2020; pp. 14–32. [Google Scholar]
- Kriwet, J.; Klug, S. Late Jurassic selachians (Chondrichthyes, Elasmobranchii) from southern Germany: Re-evaluation on taxonomy and diversity. Zitteliana A 2004, 44, 67–95. [Google Scholar]
- Kriwet, J.; Klug, S. Knorpelfische (Chondrichthyes). In Solnhofen. Ein Fenster in die Jurazeit; Arratia, G., Schultze, H.-P., Tischlinger, H., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 2015; pp. 334–359. [Google Scholar]
- Leuzinger, L.; Cuny, G.; Popov, E.; Billon-Bruyat, J.-P. A new chondrichthyan fauna from the Late Jurassic of the Swiss Jura (Kimmeridgian) dominated by hybodonts, chimaeroids and guitarfishes. Pap. Palaeontol. 2017, 3, 471–511. [Google Scholar] [CrossRef] [Green Version]
- Stumpf, S.; López-Romero, F.A.; Kindlimann, R.; Lacombat, F.; Pohl, B.; Kriwet, J. A unique hybodontiform skeleton provides novel insights into Mesozoic chondrichthyan life. Pap. Palaeontol. 2021, 7, 1479–1505. [Google Scholar] [CrossRef]
- Stumpf, S.; Etches, S.; Underwood, C.J.; Kriwet, J. Durnonovariaodus maiseyi gen. et sp. nov., a new hybodontiform shark-like chondrichthyan from the Upper Jurassic Kimmeridge Clay Formation of England. PeerJ 2021, 9, e11362. [Google Scholar] [CrossRef] [PubMed]
- Cuny, G.; Suteethorn, V.; Khamha, S. A review of the hybodont sharks from the Mesozoic of Thailand. In Proceedings of the International Conference on Geology, Geotechnology and Mineral Resources of Indochina, GEOINDO 2005, Khon Kaen, Thailand, 28–30 November 2005; Wanakao, L., Ed.; Department of Geotechnology: Khon Kaen, Thailand, 2005; pp. 588–593. [Google Scholar]
- Cuny, G.; Suteethorn, V.; Khamha, S.; Lauprasert, K.; Srisuk, P.; Buffetaut, E. The Mesozoic fossil record of sharks in Thailand. In Proceedings of the International Conference on Geology of Thailand: Towards Sustainable Development and Sufficiency Economy, GEOTHAI’2007, Bangkok, Thailand, 21–22 November 2007; Tantiwanit, W., Ed.; Department of Mineral Resources: Bangkok, Thailand, 2007; pp. 349–354. [Google Scholar]
- Cuny, G.; Liard, R.; Deesri, U.; Liard, T.; Khamha, S.; Suteethorn, V. Shark faunas from the Late Jurassic—Early Cretaceous of northeastern Thailand. Paläontol. Z. 2014, 88, 309–328. [Google Scholar] [CrossRef] [Green Version]
- Klug, S.; Tütken, T.; Wings, O.; Pfretzschner, H.-U.; Martin, T. A Late Jurassic freshwater shark assemblage (Chondrichthyes, Hybodontiformes) from the southern Junggar Basin, Xinjiang, Northwest China. Palaeobiodivers. Palaeoenviron. 2010, 90, 241–257. [Google Scholar] [CrossRef]
- Goodwin, M.B.; Clemens, W.A.; Hutchison, J.H.; Wood, C.B.; Zavada, M.S.; Kemp, A.; Duffin, C.J.; Schaff, C.R. First Mesozoic terrestrial vertebrates with associated palynostratigraphic dates from the northwestern Ethiopian Plateau. J. Vert. Paleontol. 1999, 19, 728–741. [Google Scholar] [CrossRef]
- Arratia, G.; Kriwet, J.; Heinrich, W.-D. Selachians and actinopterygians from the Upper Jurassic of Tendaguru, Tanzania. Mitt. Mus. Für Naturkd. Berl. Geowiss. Reihe 2002, 5, 207–230. [Google Scholar]
- Cione, A.L.; Acosta Hospitaleche, C.; Mennucci, J.A.; Cocca, S. The first Triassic-Jurassic shark of South America. N. Jb. Geol. Paläontol. Mh. 2002, 1, 9–18. [Google Scholar] [CrossRef]
- Soto, M.; Perea, D.; Toriño, P. New remains of Priohybodus arambourgi (Hybodontiformes: Hybodontidae) from Late Jurassice? Earliest Cretaceous deposits in Uruguay. Cret. Res. 2012, 35, 118–123. [Google Scholar] [CrossRef]
- Alvarado-Ortega, J.; Barrientos-Lara, J.I.; Espinosa-Arrubarrena, L.; Melgarejo-Damián, M.D.P. Late Jurassic marine vertebrates from Tlaxiaco, Oaxaca State, southern Mexico. Paleontol. Electron. 2014, 17, 1–25. [Google Scholar] [CrossRef]
- Dames, W. Verhandlungen der Gesellschaft. 1. Protokoll der November-Sitzung. Z. Dtsch. Geol. Gesell. 1888, 40, 777–778. [Google Scholar]
- Deecke, W. Geologie von Pommern; Verlag Gebrüder Borntraeger: Berlin, Germany, 1907; p. 302. [Google Scholar]
- Kin, A.; Gruszczynński, M.; Martill, D.; Marshall, J.D.; Błażejowski, B. Palaeoenvironment and taphonomy of a Late Jurassic (Late Tithonian) Lagerstätte from central Poland. Lethaia 2013, 46, 71–81. [Google Scholar] [CrossRef]
- Young, M.T.; Hua, S.; Steel, L.; Foffa, D.; Brusatte, S.L.; Thüring, S.; Mateus, O.; Ruiz-Omeñaca, J.I.; Havlik, P.; Lepage, Y.; et al. Revision of the Late Jurassic teleosaurid genus Machimosaurus (Crocodylomorpha, Thalattosuchia). R. Soc. Open Sci. 2014, 1, 140222. [Google Scholar] [CrossRef] [Green Version]
- Błażejowski, B.; Lambers, P.; Gieszcz, P.; Tyborowski, D.; Binkowski, M. Late Jurassic jaw bones of Halecomorph fish (Actinopterygii: Halecomorphi) studied with X-ray microcomputed tomography. Paleontol. Electron. 2015, 18, 1–10. [Google Scholar] [CrossRef]
- Tyborowski, D. A new ophthalmosaurid ichthyosaur from the Late Jurassic of Owadów-Brzezinki Quarry, Poland. Acta Palaeontol. Pol. 2016, 61, 791–803. [Google Scholar] [CrossRef] [Green Version]
- Tyborowski, D. Large predatory actinopterygian fishes from the Late Jurassic of Poland studied with X-ray microtomography. N. Jb. Geol. Paläontol. Abh. 2017, 283, 161–172. [Google Scholar] [CrossRef]
- Tyborowski, D.; Błażejowski, B. New marine reptile fossils from the Late Jurassic of Poland with implications for vertebrate faunas palaeobiogeography. Proc. Geol. Assoc. 2019, 130, 741–751. [Google Scholar] [CrossRef]
- Hoffmann, N. Knorpel- und Knochenfische aus dem Oberjura von Nordwestpolen. Diploma Thesis, University of Greifswald, Greifswald, Germany, 2006; p. 109. [Google Scholar]
- Rees, J. Neoselachian sharks from the Callovian–Oxfordian (Jurassic) of Ogrodzieniec, Zawiercie Region, Southern Poland. Palaeontology 2010, 53, 887–902. [Google Scholar] [CrossRef]
- Brüggemann, L.W. Ausführliche Beschreibung des Gegenwärtigen Zustandes des Königl. Preußischen Herzogthums Vor- und Hinterpommern–Theil 2, Band 1: Beschreibung der zu dem Gerichtsbezirk der Königl. Landescollegien in Stettin gehörigen Hinterpommerschen; Kreise, H.G., Ed.; Essenbart: Stettin, Poland, 1784; p. 460. [Google Scholar]
- Dadlez, R.; Marek, S.; Pokorski, J. Geological Map of Poland without Cainozoic Deposits (1:1.000.000); Polish Geological Institute: Warszawa, Poland, 2010.
- Dohm, B. Über den oberen Jura von Zarnglaff i. P. und seine Ammonitenfauna. Abh. Geol.-Palaeontol. Inst. Univ. Greifsw. 1925, 4, 1–40. [Google Scholar]
- Richter, D. Geologischer Führer durch die Zarnglaff-Schwanteshagener Malmbrüche. Mitt. Geol.-Palaeontol. Inst. Univers. Greifsw. 1931, 7, 1–24. [Google Scholar]
- Wilczyñski, A. Stratygrafia górnej jury w Czarnogłowach i Świętoszewie. Acta Geol. Pol. 1962, 12, 3–112. [Google Scholar]
- Dmoch, I. Ślimaki i małże górnojurajskie Czarnogłowów i Świętoszewa oraz warunki paleookologiczne w jurze górnej na Pomorzu Zachodnim. Stud. Soc. Sci. Tor. 1970, 7, 1–113. [Google Scholar]
- Hoffmann, N. Upper Jurassic Limestone Quarry of Czarnogłowy (Zarnglaff). In Geo-Pomerania Szczecin 2007: Geology Cross-Bordering the Western and Eastern European Platform. Excursion Guide; Niedermeyer, R.-O., Dobracki, R., Schütze, K., Eds.; Państwowy Instytut Geologiczny: Warszawa, Poland, 2007; pp. 44–47. [Google Scholar]
- Maisey, J.G. Hamiltonichthys mapesi, g. & sp. nov. (Chondrichthyes; Elasmobranchii), from the Upper Pennsylvanian of Kansas. Am. Mus. Novit. 1989, 2931, 1–42. [Google Scholar]
- Rees, J.; Underwood, C.J. Hybodont sharks of the English Bathonian and Callovian (Middle Jurassic). Palaeontology 2008, 51, 117–147. [Google Scholar] [CrossRef] [Green Version]
- Huxley, T.H. On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proc. Zool. Soc. Lond. 1880, 649–662. [Google Scholar]
- Bonaparte, C.L. Synopsis vertebratorum systematis. Nuovi Ann. Sci. Nat. Bologna 1838, 2, 105–133. [Google Scholar]
- Maisey, J.G. The interrelationships of phalacanthous selachians. N. Jb. Geol. Paläontol. Mh. 1975, 553–567. [Google Scholar]
- Owen, R. Lectures on the Comparative Anatomy and Physiology of the Vertebrate Animals, Delivered at the Royal College of Surgeons of England in 1844 and 1846; Part 1. Fishes; Longman: London, UK, 1846. [Google Scholar]
- Maisey, J.G. Cranial anatomy of the Lower Jurassic shark Hybodus reticulatus (Chondrichthyes: Elasmobranchii), with comments on hybodontid systematics. Am. Mus. Novit. 1987, 2878, 1–39. [Google Scholar]
- Rees, J. Early Jurassic selachians from the Hasle Formation on Bornholm, Denmark. Acta Palaeontol. Pol. 1998, 43, 439–452. [Google Scholar]
- Maisch, M.W.; Matzke, A.T. A new hybodontid shark (Chondrichthyes, Hybodontiformes) from the Lower Jurassic Posidonienschiefer Formation of Dotternhausen, SW Germany. N. Jb. Geol. Paläontol. Abh. 2016, 280, 241–257. [Google Scholar] [CrossRef]
- Agassiz, L.J.R. Recherches sur les Poissons Fossils; Petitpierre: Neuchâtel, Switzerland, 1833; Volume 5. [Google Scholar]
- Underwood, C.J.; Cumbaa, S.L. Chondrichthyans from a Cenomanian (Late Cretaceous) bonebed, Saskatchewan, Canada. Palaeontology 2010, 53, 903–944. [Google Scholar] [CrossRef]
- Rees, J.; Underwood, C.J. Hybodont sharks from the Middle Jurassic of the Inner Hebrides, Scotland. Trans. R. Soc. Edinb. Earth Sci. 2006, 96, 351–363. [Google Scholar] [CrossRef]
- Bermúdez-Rochas, D.D. New hybodont shark assemblage from the Early Cretaceous of the Basque-Cantabrian Basin. Geobios 2009, 42, 675–686. [Google Scholar] [CrossRef]
- Sharma, A.; Singh, S. A small assemblage of marine hybodont sharks from the Bathonian of the Jaisalmer Basin, India. N. Jb. Geol. Paläontol. Abh. 2021, 301, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, J.; Wright, K.; Lucas, S.G.; Spielmann, J.A.; Pence, R. Selachians from the Upper Cretaceous (Santonian) Hosta Tongue of the Point Lookout Sandstone, central New Mexico. Bull. N. M. Mus. Nat. Hist. Sci. 2011, 52, 1–54. [Google Scholar]
- Woodward, A.S. The fossil fishes of the English Wealden and Purbeck formations. Monogr. Palaeontogr. Soc. Lond. 1916, 69, 1–48. [Google Scholar] [CrossRef]
- Underwood, C.J.; Rees, J. Selachian faunas from the earliest Cretaceous Purbeck Group of Dorset, southern England. Pap. Palaeontol. 2002, 68, 83–101. [Google Scholar]
- Duffin, C.J.; Sweetman, S.C. 17. Sharks. In English Wealden Fossils; Batten, D.J., Ed.; Field Guide to Fossils 11; Palaeontological Association: London, UK, 2011; pp. 205–224. [Google Scholar]
- Turmine-Juhel, P.; Wilks, R.; Brockhurst, D.; Austen, P.A.; Duffin, C.J.; Benton, M.J. Microvertebrates from the Wadhurst Clay Formation (Lower Cretaceous) of Ashdown Brickworks, East Sussex, UK. Proc. Geol. Assoc. 2019, 130, 752–769. [Google Scholar] [CrossRef]
- Pinheiro, F.L.; de Figueiredo, A.E.Q.; Dentzien-Dias, P.C.; Fortier, D.C.; Schultz, C.L.; Viana, M.S.S. Planohybodus marki sp. nov., a new fresh-water hybodontid shark from the Early Cretaceous of northeastern Brazil. Cret. Res. 2013, 41, 210–216. [Google Scholar] [CrossRef]
- Rees, J. Jurassic and Early Cretaceous selachians—Focus on southern Scandinavia. Lund Publ. Geol. 2001, 153, 1–19. [Google Scholar]
- Vullo, R. Direct evidence of hybodont shark predation on Late Jurassic ammonites. Naturwissenschaften 2011, 98, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Vullo, R.; Abit, D.; Ballèvre, M.; Billon-Bruyat, J.-P.; Bourgeais, R.; Buffetaut, E.; Daviero-Gomez, V.; Garcia, G.; Gomez, B.; Mazin, J.-M.; et al. Palaeontology of the Purbeck-type (Tithonian, Late Jurassic) bonebeds of Chassiron (Oléron Island, western France). Comptes Rendus Palevol 2014, 13, 421–441. [Google Scholar] [CrossRef]
- Case, G.R. A new selachian fauna from the Coleraine Formation (Upper Cretaceous ⁄ Cenomanian) of Minnesota. Palaeontogr. Abt. A 2001, 261, 103–112. [Google Scholar]
- Thurmond, J.T. Cartilaginous fishes of the Trinity Group and related rocks (Lower Cretaceous) of North Central Texas. Southeast. Geol. 1971, 13, 207–227. [Google Scholar]
- Case, G.R. A new selachian fauna from the Judith River formation (Campanian) of Montana. Palaeontogr. Abt. A 1978, 160, 176–205. [Google Scholar]
- Case, G.R.; Cappetta, H. Additions to the elasmobranch fauna from the late Cretaceous of New Jersey (lower Navesink Formation, early Maastrichtian). Palaeovertebrata 2004, 33, 1–16. [Google Scholar]
- Cicimurri, D.J.; Ciampalgio, C.N.; Runyon, K.E. Late Cretaceous elasmobranchs from the Eutaw Formation at Luxapalila Creek, Lowndes County, Mississippi. PalArch’s J. Vert. Palaeontol. 2014, 11, 1–36. [Google Scholar]
- Jaekel, O. Die Selachier aus dem oberen Muschelkalk Lothringens. Abh. Geol. Spec. Els.-Lothr. 1889, 3, 275–340. [Google Scholar]
- Rees, J. Interrelationships of Mesozoic hybodont sharks as indicated by dental morphology-preliminary results. Acta Geol. Pol. 2008, 58, 217–221. [Google Scholar]
- Egerton, P.M.G. On some new genera and species of fossil fishes. Ann. Mag. Nat. Hist. 1854, 13, 433–436. [Google Scholar] [CrossRef]
- Sauvage, H.E. Notes sur les poissons fossils (suite). XVI–XXIII. Bull. Soc. Géol. Fr. 1880, 8, 451–462. [Google Scholar]
- Stromer, E. Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharije-Stufe (unterstes Cenoman). 9. Die Plagiostomen, mit einem Anhang über käno- und mesozoische Rückenflossenstacheln von Elasmobranchiern. Abh. Bayer. Akad. Wiss. Math.-Naturwiss. Kl. 1927, 31, 1–64. [Google Scholar]
- Woodward, A.S. On some remains of the Extinct Selachian Asteracanthus from the Oxford Clay of Peterborough, preserved in the collection of Alfred, N. Leeds, Esq., of Eyebury. J. Nat. Hist. 1888, 2, 336–342. [Google Scholar] [CrossRef] [Green Version]
- Szabó, M.; Főzy, I. Asteracanthus (Hybodontiformes: Acrodontidae) remains from the Jurassic of Hungary, with the description of a new species and with remarks on the taxonomy and paleobiology of the genus. N. Jb. Geol. Paläontol. Abh. 2020, 297, 295–309. [Google Scholar] [CrossRef]
- Duffin, C.J. The dentition of Hybodus hauffianus Fraas, 1895 (Toarcian, Early Jurassic). Stuttg. Beitr. Naturkd. B 1997, 256, 1–20. [Google Scholar]
- Maisey, J.G. Cranial antomy of Hybodus basanus Egerton from the Lower Cretaceous of England. Am. Mus. Novit. 1983, 2758, 1–26. [Google Scholar]
- Dunker, W. Ueber einen neuen Asteracanthus aus dem Korallenkalk des Lindner Berges bei Hannover. Palaeontographica 1851, 1, 188–189. [Google Scholar]
- Dunker, W. Asteracanthus ornatissimus, Agassiz, aus dem Korallenkalk von Hoheneggelsen bei Hildesheim. Palaeontographica 1851, 1, 316–318. [Google Scholar]
- Platnauer, H.M. Note on Hybodus obtusus. Ann. Rep. Yorks. Phil. Soc. 1887, 35–36. [Google Scholar]
- Priem, F. Etude des poissons fossiles du Bassin Parisien (supplément). Ann. Paléontol. 1911, 6, 1–144. [Google Scholar]
- Casier, E. Contributions a l’etude des Poissons fossils de la Belgique. XII. Selaciens et Holocephales sinemuriens de la province de Luxembourg. Bull. Inst. R. Sci. Nat. Bel. 1959, 35, 1–27. [Google Scholar]
- Rieppel, O. The hybodontiform sharks from the Middle Triassic of Mte. San Giorgio, Switzerland. N. Jb. Geol. Paläontol. Abh. 1981, 16, 324–353. [Google Scholar]
- Goto, M. Palaeozoic and early Mesozoic fish faunas of the Japanese Islands. Isl. Arc. 1994, 3, 247–254. [Google Scholar] [CrossRef]
- Cuny, G.; Srisuk, P.; Khamha, S.; Suteethorn, V.; Tong, H. A new elasmobranch fauna from the Middle Jurassic of southern Thailand. In Late Palaeozoic and Mesozoic Ecosystems in SE Asia; Buffetaut, E., Cuny, G., Le Loeuff, J., Suteethorn, V., Eds.; Special Publications 315; The Geological Society: London, UK, 2009; pp. 97–113. [Google Scholar] [CrossRef]
- Guinot, G.; Cappetta, H.; Adnet, S. A rare elasmobranch assemblage from the Valanginian (Lower Cretaceous) of southern France. Cret. Res. 2014, 48, 54–84. [Google Scholar] [CrossRef]
- Vincent, P.; Martin, J.E.; Fischer, V.; Suan, G.; Khalloufi, B.; Suchéras-Marx, B.; Lena, A.; Janneau, K.; Rousselle, B.; Rulleau, L. Marine vertebrate remains from the Toarcian–Aalenian succession of southern Beaujolais, Rhône, France. Geol. Mag. 2013, 150, 822–834. [Google Scholar] [CrossRef] [Green Version]
- Citton, P.; Fabbi, S.; Cipriani, A.; Jansen, M.; Romano, M. Hybodont dentition from the Upper Jurassic of Monte Nerone Pelagic Carbonate Platform (Umbria-Marche Apennine, Italy) and its ecological implications. Geol. J. 2019, 54, 278–290. [Google Scholar] [CrossRef] [Green Version]
- Peyer, B. Die schweizerischen Funde von Asteracanthus (Strophodus). Schweiz. Palaeontol. Abh. 1946, 64, 1–101. [Google Scholar]
- Rigal, S.; Cuny, G. On the rarity of anterior teeth of Asteracanthus magnus (Euselachii: Hybodontiformes). N. Jb. Geol. Paläontol. Abh. 2016, 279, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Bajpai, S.; Pandey, P.; Ghosh, T.; Bhattacharya, D. Hybodont sharks from the Jurassic of Jaisalmer, western India. Hist. Biol. 2021. [Google Scholar] [CrossRef]
- Owen, R. Description of a great part of a jaw with the teeth of Strophodus medius, Ow., from the Oolite of Caen in Normandy. Geol. Mag. 1869, 6, 193–196. [Google Scholar] [CrossRef] [Green Version]
- Martill, D.M. Fossils of the Oxford Clay; Martill, D.M., Hudson, J.D., Eds.; Field Guide to Fossils 4; Palaeontological Association: London, UK, 1991; pp. 197–225. [Google Scholar]
- Pfeil, F.H. Ein neues Asteracanthus-Gebiss aus den Kieselplattenkalken (Oberjura, Tithonium, Malm Zeta 3, Mörnsheim-Formation) des Besuchersteinbruchs in Mühlheim. Fr. Bayer; Staatssammlung für Paläontologie und Historische Geologie München: München, Germany, 2011; Volume 39, pp. 36–60. [Google Scholar]
- Münster, G.V. Beschreibung einiger neuer Fische aus den lithographischen Schiefern von Bayern. Beitr. Petrefactenkd. 1842, 5, 55–64. [Google Scholar]
- Wagner, J.A. Charakteristik neuer Arten von Knorpelfischen aus den lithographischen Schiefern der Umgegend von Solnhofen. Gelehrte Anz. Königl. Bayer. Akad. Wiss. 1857, 44, 288–293. [Google Scholar]
- Kriwet, J.; Klug, S. Diversity and biogeography patterns of Late Jurassic neoselachians (Chondrichthyes: Elasmobranchii). In Fishes and the Break-Up of Pangaea; Cavin, L., Longbottom, A., Richter, M., Eds.; Special Publications 295; The Geological Society: London, UK, 2008; pp. 55–70. [Google Scholar] [CrossRef]
- Leuzinger, L.; Kocsis, L.; Billon-Bruyat, J.-P.; Spezzaferri, S.; Vennemann, T. Stable isotope study of a new chondrichthyan fauna (Kimmeridgian, Porrentruy, Swiss Jura): An unusual freshwater-influenced isotopic composition for the hybodont shark Asteracanthus. Biogeosciences 2015, 12, 6945–6954. [Google Scholar] [CrossRef] [Green Version]
- Maisey, J.G. The anatomy and interrelationships of Mesozoic hybodont sharks. Am. Mus. Novit. 1982, 2724, 1–48. [Google Scholar]
- Soler-Gijón, R.; Poyato-Ariza, F.J.; Maisey, J.G.; Lane, J.A. Chondrichthyes. In Las Hoyas: A Cretaceous Wetland; Poyato-Ariza, F.J., Buscalioni, A.D., Fregenal-Martinez, M., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 2016; pp. 103–113. [Google Scholar]
- Duffin, C.J. 16. Fishes—Sharks and rays. In Fossils from the Lower Lias of the Dorset Coast; Lord, A.R., Davis, P.G., Eds.; Field Guide to Fossils 13; Palaeontological Association: London, UK, 2010; pp. 317–340. [Google Scholar]
- Stumpf, S.; Kriwet, J. A new Pliensbachian elasmobranch (Vertebrata, Chondrichthyes) assemblage from Europe, and its contribution to the understanding of late Early Jurassic elasmobranch diversity and distributional patterns. PalZ 2019, 93, 637–658. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.G.; Smith, D.G.; Funnell, B.M. Atlas of Mesozoic and Cenozoic Coastlines; Cambridge University Press: Cambridge, London, 2004; p. 112. [Google Scholar]
- Lott, G.K.; Wong, T.E.; Dusar, M.; Andsbjerg, J.; Mönnig, E.; Feldman-Olszewska, A.; Verreussel, R.M.C.H. Jurassic. In Petroleum Geological Atlas of the Southern Permian Basin Area; Doornenbal, J.C., Stevenson, A.G., Eds.; EAGE Publications B.V.: Houten, The Netherlands, 2010; pp. 175–193. [Google Scholar]
- Madzia, D.; Szczygielski, T.; Wolniewicz, A.S. The giant pliosaurid that wasn’t—revising the marine reptiles from the Kimmeridgian, Upper Jurassic, of Krzyżanowice, Poland. Acta Palaeontol. Pol. 2021, 66, 99–129. [Google Scholar] [CrossRef]
- Rees, J.; Underwood, C.J. The status of the shark genus Lissodus, and the position of nominal Lissodus species within the Hybodontoidea. J. Vert. Paleontol. 2002, 22, 471–479. [Google Scholar] [CrossRef]
- Underwood, C.J. Diversification of the Neoselachii (Chondrichthyes) during the Jurassic and Cretaceous. Paleobiology 2006, 32, 215–235. [Google Scholar] [CrossRef] [Green Version]
- Kriwet, J.; Kiessling, W.; Klug, S. Diversification trajectories and evolutionary life-history traits in early sharks and batoids. Proc. R. Soc. B 2009, 276, 945–951. [Google Scholar] [CrossRef] [Green Version]
- Rees, J.; Cuny, G.; Pouech, J.; Mazin, J.-M. Non-marine selachians from the basal Cretaceous of Charente, SW France. Cret. Res. 2013, 44, 122–131. [Google Scholar] [CrossRef]
- Klug, C.; Schweigert, G.; Hoffmann, R.; Weis, R.; De Baets, K. Fossilized leftover falls as sources of palaeoecological data: A ‘pabulite’ comprising a crustacean, a belemnite and a vertebrate from the Early Jurassic Posidonia Shale. Swiss J. Palaeontol. 2021, 140, 10. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stumpf, S.; Meng, S.; Kriwet, J. Diversity Patterns of Late Jurassic Chondrichthyans: New Insights from a Historically Collected Hybodontiform Tooth Assemblage from Poland. Diversity 2022, 14, 85. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020085
Stumpf S, Meng S, Kriwet J. Diversity Patterns of Late Jurassic Chondrichthyans: New Insights from a Historically Collected Hybodontiform Tooth Assemblage from Poland. Diversity. 2022; 14(2):85. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020085
Chicago/Turabian StyleStumpf, Sebastian, Stefan Meng, and Jürgen Kriwet. 2022. "Diversity Patterns of Late Jurassic Chondrichthyans: New Insights from a Historically Collected Hybodontiform Tooth Assemblage from Poland" Diversity 14, no. 2: 85. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020085