
Serosurvey of Selected Zoonotic Pathogens in Polar Bears (Ursus maritimus Phipps, 1774) in the Russian Arctic

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hale, P.; Aars, J.; Bakhtov, A.; Belikov, S.; DeBruyn, T.; Colligan, M.; Ekker, M.; Erbs-Maibing, S.; Gordon, A.; Hansen, H.; et al. Circumpolar Action Plan: Conservation Strategy for Polar Bears. A product of the representatives of the parties to the 1973 Agreement on the Conservation of Polar Bears; Ilulissat, Greenland, 2015; 80p, Available online: https://www.polarbearscanada.ca/en/management/international-initiatives/circumpolar-action-plan-polar-bears (accessed on 4 May 2022).
- Harvell, C.D.; Mitchell, C.E.; Ward, J.R.; Altizer, S.; Dobson, A.P.; Ostfeld, R.S.; Samuel, M.D. Climate Warming and Disease Risks for Terrestrial and Marine Biota. Science 2002, 296, 2158–2162. [Google Scholar] [CrossRef] [Green Version]
- Hueffer, K.; O’Hara, T.M.; Follmann, E.H. Adaptation of mammalian host-pathogen interactions in a changing arctic environment. Acta Vet. Scand. 2011, 53, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tryland, M.; Derocher, A.E.; Wiig, O.; Godfroid, J. Brucella sp. antibodies in polar bears from Svalbard and the Barents sea. J. Wildl. Dis. 2001, 37, 523–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hara, T.M.; Holcomb, D.; Elzer, P.; Estepp, J.; Perry, Q.; Hagius, S.; Kirk, C. Brucella Species Survey in polar bears (Ursus maritimus) of Northern Alaska. J. Wildl. Dis. 2010, 46, 687–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rah, H.; Chomel, B.B.; Kasten, R.; Hew, C.H.; Farver, T.B.; Follmann, E.H.; Garner, G.; Amstrup, S.C. Serosurvey of selected zoonotic agents in polar bears (Ursus maritimus). Vet. Rec. 2005, 156, 7–13. [Google Scholar] [CrossRef]
- Pilfold, N.W.; Richardson, E.S.; Ellis, J.; Jenkins, E.; Scandrett, W.B.; Hernández-Ortiz, A.; Buhler, K.; McGeachy, D.; Al-Adhami, B.; Konecsni, K.; et al. Long-term increases in pathogen seroprevalence in polar bears (Ursus maritimus) influenced by climate change. Glob. Chang. Biol. 2021, 27, 4481–4497. [Google Scholar] [CrossRef]
- Seymour, J.; Horstmann-Dehn, L.; Rosa, C.; Lopez, J. Occurrence and genotypic analysis of Trichinella species in Alaska marine-associated mammals of the Bering and Chukchi seas. Vet. Parasitol. 2014, 200, 153–164. [Google Scholar] [CrossRef]
- Follmann, E.H.; Garner, G.W.; Evermann, J.F.; McKeirnan, A.J. Serological evidence of morbillivirus infection in polar bears (Ursus maritimus) from Alaska and Russia. Vet. Rec. 1996, 138, 615–618. [Google Scholar] [CrossRef]
- Philippa, J.D.W.; Martina, B.E.E.; Kuiken, T.; Van de Bildt, M.W.G.; Osterhaus, A.D.M.E.; Leighton, F.A.; Daoust, P.Y.; Nielsen, O.; Pagliarulo, M.; Schwantje, H.; et al. Antibodies to selected pathogens in free-ranging terrestrial carnivores and marine mammals in Canada. Vet. Rec. 2004, 155, 135–140. [Google Scholar] [CrossRef]
- Tryland, M.; Neuvonen, E.; Huovilainen, A.; Tapiovaara, H.; Osterhaus, A.; Wiig, Ø.; Derocher, A.E. Serologic survey for selected virus infections in polar bears at Svalbard. J. Wildl. Dis. 2005, 41, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Kirk, C.M.; Amstrup, S.; Swor, R.; Holcomb, D.; O’Hara, T.M. Morbillivirus and Toxoplasma Exposure and Association with Hematological Parameters for Southern Beaufort Sea Polar Bears: Potential Response to Infectious Agents in a Sentinel Species. EcoHealth 2010, 7, 321–331. [Google Scholar] [CrossRef]
- Une, Y.; Mori, T. Tuberculosis as a zoonosis from a veterinary perspective. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 415–425. [Google Scholar] [CrossRef]
- Alekseev, A.Y.; Reguzova, A.Y.; Rozanova, E.I.; Abramov, A.V.; Tumanov, Y.V.; Kuvshinova, I.N.; Shestopalov, A. Detection of specific antibodies to morbilliviruses, Brucella and Toxoplasma in the Black Sea dolphin Tursiops truncatus ponticus and the beluga whale Delphinapterus leucas from the Sea of Okhotsk in 2002–2007. Russ. J. Mar. Biol. 2009, 35, 494–497. [Google Scholar] [CrossRef]
- Nymo, I.H.; Godfroid, J.; Åsbakk, K.; Larsen, A.K.; das Neves, C.G.; Rødven, R.; Tryland, M. A protein A/G indirect enzyme-linked immunosorbent assay for the detection of anti-Brucella antibodies in Arctic wildlife. J. Vet. Diagn. Investig. 2013, 25, 369–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarnke, R.L.; Harder, T.C.; Vos, H.W.; Ver Hoef, J.M. Osterhaus, A.D. Serologic survey for phocid herpesvirus-1 and -2 in marine mammals from Alaska and Russia. J. Wildl. Dis. 1997, 33, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Frey, H.-R.; Liess, B.; Haas, L.; Lehmann, H.; Marschall, H.-J. Herpesvirus in Harbour Seals (Phoca vitulina): Isolation, Partial Characterization and Distribution4. J. Vet. Med. Ser. B 1989, 36, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Stenvers, O.; Zhang, X.M.; Bries, E.T.; Ludwig, H. Characterization of viruses involved in the seal mass mortality. In Proceedings of the 6th International Conference on Wildlife Diseases, Berlin, Germany, 6–11 August 1990. [Google Scholar]
- Stenvers, O.; Zhang, X.; Ludwig, H. Herpesvirus infection in seals: A summary of present knowledge. Rev. Sci. Et Tech. 1992, 11, 1151–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maness, H.T.; Nollens, H.H.; Jensen, E.D.; Goldstein, T.; LaMere, S.; Childress, A.; Sykes, J.; Leger, J.S.; Lacave, G.; Latson, F.E. Phylogenetic analysis of marine mammal herpesviruses. Vet. Microbiol. 2011, 149, 23–29. [Google Scholar] [CrossRef]
- Derko, A.A.; Alekseev, A.Y.; Sharshov, K.A.; Burkanov, V.N.; Jamalutdinov, J.M.; Abdulgalimova, G.N.; Ibnumaskhudova, P.M. Modern views on the circulation of herpes viruses in the Okhotsk sea Steller sea lion (Eumetopias jubatus, Schreber, 1776). South Russ. Ecol. Dev. 2019, 14, 35–47. [Google Scholar] [CrossRef]
- Borst, G.H.A.; Walvoort, H.C.; Reijnders, P.J.H.; van der Kamp, J.S.; Osterhaus, A.D. An outbreak of a herpesvirus infection in harbor seals (Phoca vitulina). J. Wildl. Dis. 1986, 22, 1–6. [Google Scholar] [CrossRef]
- Dea, S.; Verbeek, A.J.; Tijssen, P. Antigenic and genomic relationships among turkey and bovine enteric coronaviruses. J. Virol. 1990, 64, 3112–3118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agnihothram, S.; Gopal, R.; Yount, B.L., Jr.; Donaldson, E.F.; Menachery, V.D.; Graham, R.L.; Scobey, T.D.; Gralinski, L.E.; Denison, M.R.; Zambon, M.; et al. Evaluation of Serologic and Antigenic Relationships Between Middle Eastern Respiratory Syndrome Coronavirus and Other Coronaviruses to Develop Vaccine Platforms for the Rapid Response to Emerging Coronaviruses. J. Infect. Dis. 2014, 209, 995–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, G.; Jahans, K.L.; Reid, R.J.; Ross, H.M. Isolation of Brucella species from cetaceans, seals and an otter. Vet. Rec. 1996, 138, 583–586. [Google Scholar] [CrossRef]
- Nielsen, O.; Nielsen, K.; Stewart, R.E. Serologic Evidence of Brucella Spp. Exposure in Atlantic Walruses (Odobenus Rosmarus Rosmarus) and Ringed Seals (Phoca Hispida) of Arctic Canada. Arctic 1996, 49, 383–386. [Google Scholar] [CrossRef] [Green Version]
- Jepson, P.D.; Brew, S.; MacMillan, A.P.; Baker, J.R.; Barnett, J.; Kirkwood, J.K.; Kuiken, T.; Robinson, I.R.; Simpson, V.R. Antibodies to Brucella in marine mammals around the coast of England and Wales. Vet. Rec. 1997, 141, 513–515. [Google Scholar] [CrossRef]
- Tryland, M.; Kleivane, L.; Alfredsson, A.; Kjeld, M.; Arnason, A.; Stuen, S.; Godfroid, J. Evidence of Brucella infection in marine mammals in the North Atlantic Ocean. Vet. Rec. 1999, 144, 588–592. [Google Scholar] [CrossRef]
- Retamal, P.; Abalos, P.; Blank, O.; Torres, D. Detection of anti-Brucella antibodies in pinnipeds from the Antarctic territory. Vet. Rec. 2000, 146, 166–167. [Google Scholar] [CrossRef]
- Van Bressem, M.F.; Van Waerebeek, K.; Raga, J.A.; Godfroid, J.; Brew, S.D.; MacMillan, A.P. Serological evidence of Brucella species infection in odontocetes from the South Pacific and the Mediterranean. Vet. Rec. 2001, 148, 657–661. [Google Scholar] [CrossRef]
- Alekseev, A.Y.; Shpak, O.V.; Adamenko, L.S.; Glazov, D.M.; Galkina, I.V.; Schelkanov, M.Y.; Shestopalov, A.M. Serological Detection of Causative Agents of Infectious and Invasive Diseases in the Beluga Whale Delphinapterus leucas (Pallas, 1776) (Cetacea: Monodontidae) from Sakhalinsky Bay. Russ. J. Mar. Biol. 2017, 43, 485–490. [Google Scholar] [CrossRef]
- Akhmedov, D.R. Brucellosis. In Infectious Diseases: National Recommendations, 2nd ed.; Yushchuk, N.D., Vengerov, Y.Y., Eds.; Series of National Guidelines; GEOTAR-Media: Moscow, Russia, 2019; pp. 309–317. ISBN 978-5-9704-4912-7. [Google Scholar]
- Gordienko, L.N.; Kulikova, E.V.; Anisimova, A.A. The domestic dog (Canis familiaris) is a reservoir of the causative agent of brucellosis of reindeer in natural foci of infection. Vet. Pathol. 2007, 3, 41–46. (In Russian) [Google Scholar]
- Layshev, K.A.; Zabrodin, V.A.; Prokudin, A.V.; Samandas, A.M. The Evaluation of the Epizootic Situation in the Populations of Wild Reindeer of the Arctic Zone of the Russian Federation (Literature Review). Top. Issues Vet. Biol. 2015, 4, 38–46. Available online: http://www.invetbio.spb.ru/avvb/AVVB_2015_04.pdf (accessed on 4 May 2022). (In Russian).
- Zheludkov, M.M.; Tsirelson, L.E. Reservoirs of Brucella infection in nature. Zool. Zhurnal. 2010, 89, 53–60. ISSN 00445134(In Russian) [Google Scholar] [CrossRef]
- Kerschengoltz, B.M.; Chernyavsky, V.F.; Repin, V.E.; Nikiforov, O.I.; Sofronova, O.N. Influence of global climatic changes on realization of infectious potential diseases of the population in the Russian arctic regions (on an example of Yakutia) Review. Ekol. Cheloveka (Hum. Ecol.) 2009, 9, 34–39. Available online: https://www.elibrary.ru/download/elibrary_12962854_38039529.pdf (accessed on 4 May 2022). (In Russian).
- Ovsyanikov, N.G. Polar bears on New Siberian Islands in ice free seasons. In Marine Mammals of the Holarctic, Collection of Scientific Papers; RPO “Marine Mammal Council”: Moscow, Russia, 2015; Volume 2, pp. 45–52. [Google Scholar]
- Yushchuk, N.D.; Ogienko, O.L. Mycoplasmosis. In Infectious Diseases: National Recommendations, 2nd ed.; Yushchuk, N.D., Vengerov, Y.Y., Eds.; Series of National Guidelines; GEOTAR-Media: Moscow, Russia, 2019; pp. 478–484. ISBN 978-5-9704-4912-7. [Google Scholar]
- Alekseev, A.I.; Ustinova, E.N.; Tumanov, I.V.; Zasypkin, M.I.; Burkanov, V.N.; Kuvshinova, I.N.; Shestopalov, A.M. Detection of specific antibodies to morbilliviruses, Toxoplasma, and Brucella species in eared seals in North-West of Pacific Ocean. J. Microbiol. Epidemiol. Immunobiol. 2008, 1, 63–65. [Google Scholar]
- Morris, P.J.; Legendre, A.M.; Bowersock, T.L.; Brooks, D.E.; Krahwinkel, D.J.; Shires, G.M.H.; Walker, M.A. Diagnosis and Treatment of Systemic Blastomycosis in a Polar Bear (Ursus maritimus) with Itraconazole. J. Zoo Wildl. Med. 1989, 20, 336–345. [Google Scholar]
- Finn, J.P. Pyocephalus and gastritis in a polar bear (Thalarctus maritimus). J. Am. Vet. Med. Assoc. 1969, 155, 1086–1089. [Google Scholar]
- Pozio, E.; Murrell, K.D. Systematics and epidemiology of Trichinella. Adv. Parasitol. 2006, 63, 367–439. [Google Scholar] [CrossRef]
- Uspensky, A.; Bukina, L.; Odoevskaya, I.; Movsesyan, S.; Voronin, M. The epidemiology of trichinellosis in the Arctic territories of a Far Eastern District of the Russian Federation. J. Helminthol. 2018, 93, 42–49. [Google Scholar] [CrossRef]
- Bolshakova, V.A.; Grigoriev, I.I. Helminthiasis of young domestic reindeer in the mountain taiga zone of Yakutia. Hippology Vet. Med. [Ippologiay i veterinariya] 2019, 1, 87–90. [Google Scholar] [CrossRef]
- Spiridonov, V.; Solovyev, B.; Chuprina, E.; Pantyulin, A.; Sazonov, A.; Nedospasov, A.; Stepanova, S.; Belikov, S.; Chernova, N.; Gavrilo, M.; et al. Importance of oceanographical background for a conservation priority areas network planned using MARXAN decision support tool in the Russian Arctic seas. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 52–64. [Google Scholar] [CrossRef] [Green Version]
- Polar Bear Diet. Available online: https://arcticwwf.org/species/polar-bear/diet/ (accessed on 4 May 2022).
- Andersen, M.; Lie, E.; Derocher, A.; Belikov, S.; Bernhoft, A.; Boltunov, A.; Garner, G.; Skaare, J.; Wiig, O. Geographic variation of PCB congeners in polar bears (Ursus maritimus) from Svalbard east to the Chukchi Sea. Polar Biol. 2001, 24, 231–238. [Google Scholar] [CrossRef]
- Lie, E.; Bernhoft, A.; Riget, F.; Belikov, S.; Boltunov, A.; Derocher, A.; Garner, G.; Wiig, Ø.; Skaare, J. Geographical distribution of organochlorine pesticides (OCPs) in polar bears (Ursus maritimus) in the Norwegian and Russian Arctic. Sci. Total Environ. 2003, 306, 159–170. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
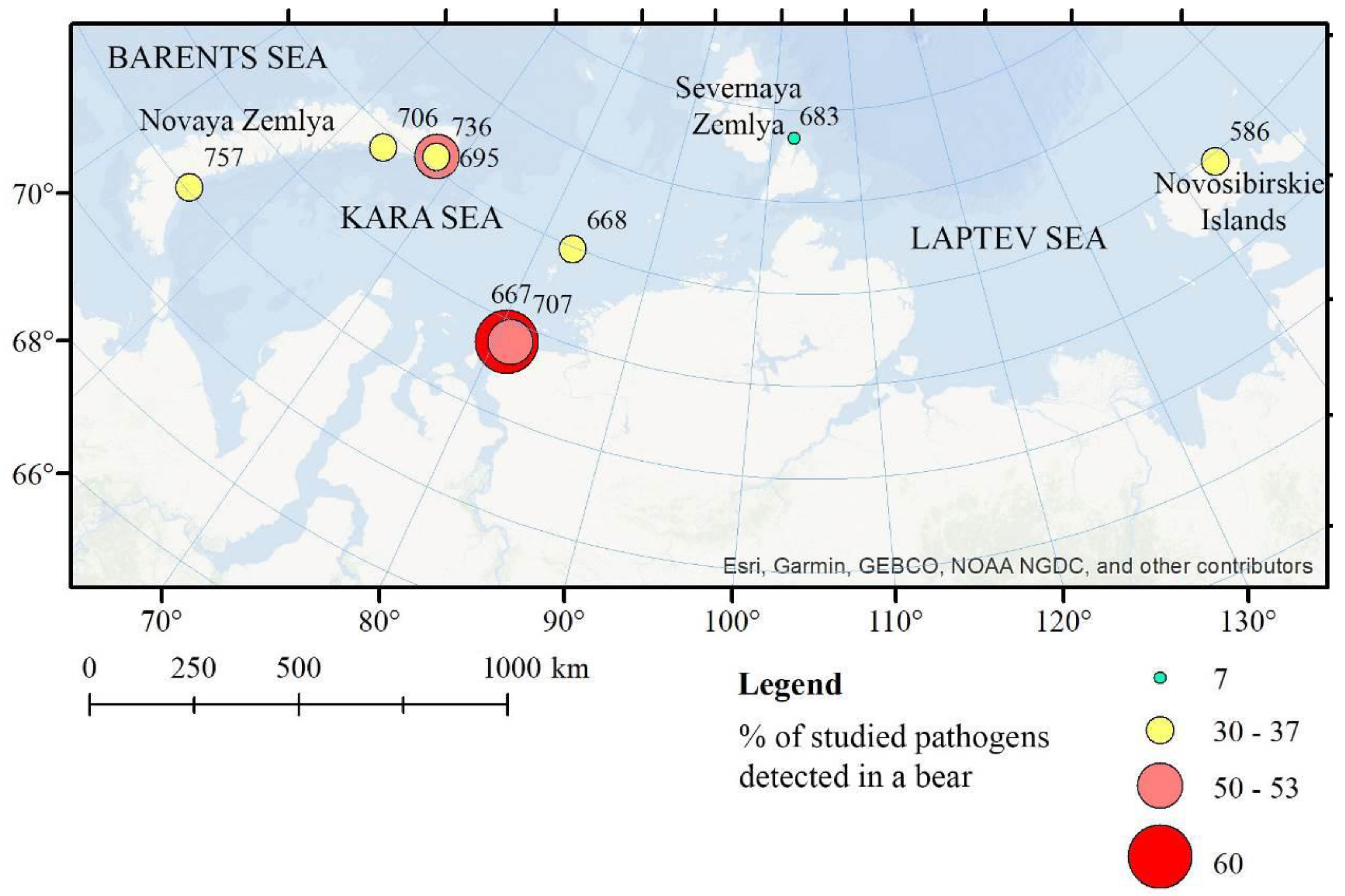
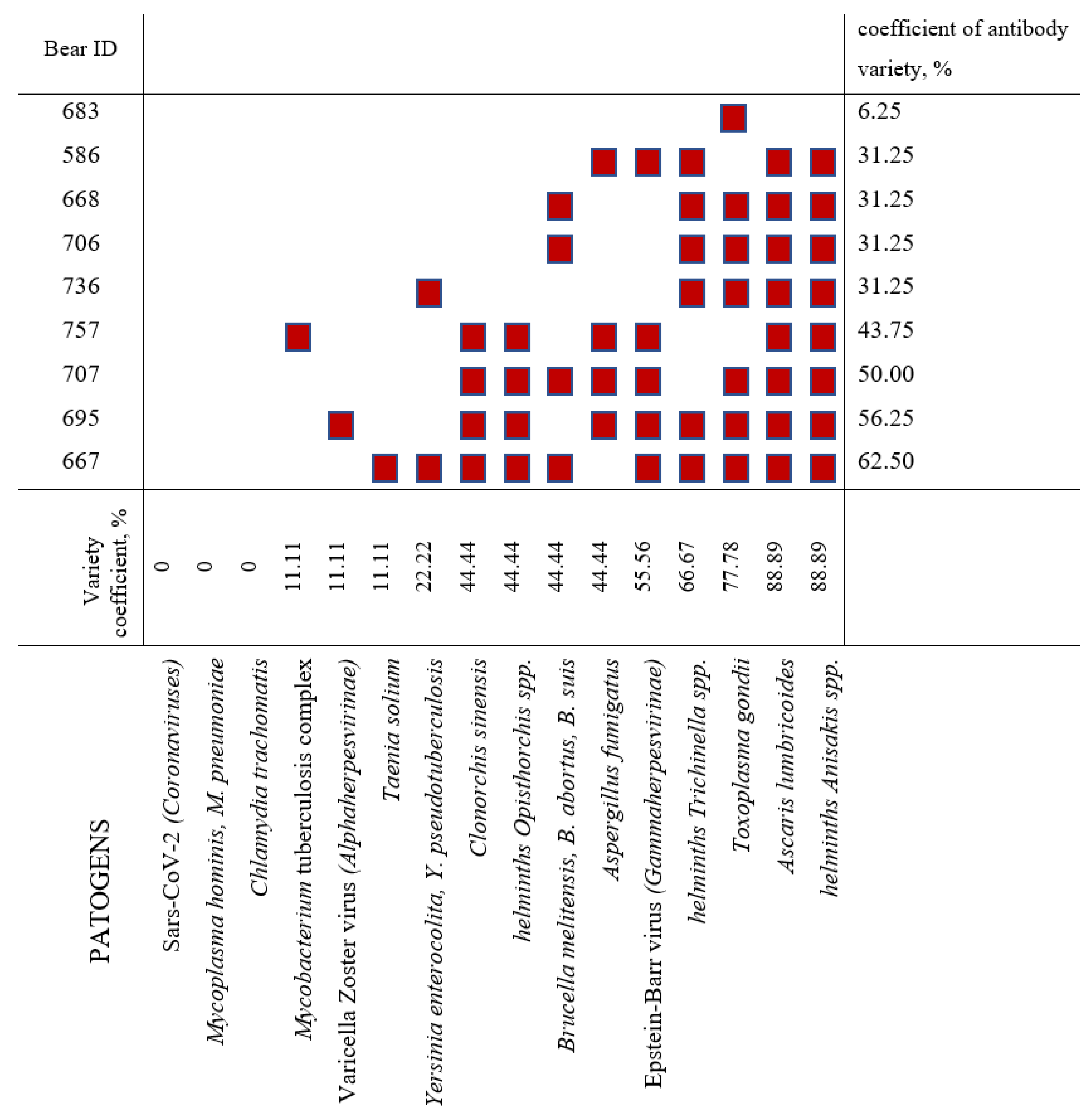
| Animal ID | Sex | Age Category | Sampling Date | Place of Sampling |
| 586 | Female | Mature | 30 August 2016 | Kotelny Island (Novosibirskie Islands) |
| 667 | Female | 2+ years old | 13 September 2016 | near Dikson (urban-type settlement, Kara Sea) |
| 668 | Female | Mature | 14 September 2016 | Troynoy Island (Izvestiy TSIK Islands, Kara Sea) |
| 683 | Male | Mature | 22 September 2016 | Bolshevik Island (Severnaya Zemlya) |
| 695 | Male | Mature | 26 September 2016 | archipelago Novaya Zemlya |
| 706 | Male | Mature | 27 September 2016 | archipelago Novaya Zemlya |
| 707 | Female | 1+ year old | 28 September 2016 | near Dikson (urban-type settlement, Kara Sea) |
| 736 | Female | Mature | 05 November 2016 | archipelago Novaya Zemlya |
| 757 | Female | Mature | 06 November 2016 | archipelago Novaya Zemlya |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alekseev, A.Y.; Boltunov, A.N.; Derko, A.A.; Sharshov, K.A.; Adamenko, L.S.; Shestopalov, A.M. Serosurvey of Selected Zoonotic Pathogens in Polar Bears (Ursus maritimus Phipps, 1774) in the Russian Arctic. Diversity 2022, 14, 365. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050365
Alekseev AY, Boltunov AN, Derko AA, Sharshov KA, Adamenko LS, Shestopalov AM. Serosurvey of Selected Zoonotic Pathogens in Polar Bears (Ursus maritimus Phipps, 1774) in the Russian Arctic. Diversity. 2022; 14(5):365. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050365
Chicago/Turabian StyleAlekseev, Alexander Yu., Andrei N. Boltunov, Anastasiya A. Derko, Kirill A. Sharshov, Lyubov S. Adamenko, and Alexander M. Shestopalov. 2022. "Serosurvey of Selected Zoonotic Pathogens in Polar Bears (Ursus maritimus Phipps, 1774) in the Russian Arctic" Diversity 14, no. 5: 365. https://0-doi-org.brum.beds.ac.uk/10.3390/d14050365