
Vertical Distribution of Beetles (Coleoptera) in Pine Forests in Central European Russia



1
Joint Directorate of the Mordovia State Nature Reserve and National Park «Smolny», Saransk 430005, Republic of Mordovia, Russia
2
Prisursky State Nature Reserve, Cheboksary 428034, Chuvash Republic, Russia
3
Institute of Environmental and Agricultural Biology (X-BIO), Tyumen State University, Tyumen 625003, Russia
*
Author to whom correspondence should be addressed.
Diversity 2022, 14(8), 622; https://0-doi-org.brum.beds.ac.uk/10.3390/d14080622
Submission received: 14 July 2022
/
Revised: 28 July 2022
/
Accepted: 2 August 2022
/
Published: 3 August 2022
(This article belongs to the Special Issue Vertical Distribution of Animals in Forest Ecosystems)
Abstract
:Research on the Arthropoda distribution in temperate forest ecosystems has shown that communities are distributed vertically in unequal numbers. However, this issue still has research gaps in relation to the vertical stratification of Coleoptera in pine forests of the temperate zone. This study was conducted in the Republic of Mordovia situated in central part of European Russia. We used fermental traps to collect Coleoptera (the bait is fermenting beer with sugar and honey). The sampling was conducted from May to September 2021 in five sites of pine forests. One hundred and twenty-five species from 36 families were identified. The highest species richness was found in Nitidulidae and Cerambycidae (19 species each), Elateridae (13), Curculionidae (7) and Scarabaeidae (6). The highest number of species (84 species) was obtained at the height of 1.5 m, while the smallest species richness (44 species) was found at the height of 12.0 m. At the height of 12 m, we also registered the minimum number of specimens. Twenty-four species from 11 families were common to all the heights studied. Cryptarcha strigata and Glischrochilus grandis preferred heights of 3 and 1.5 m. Cryptarcha undata and Protaetia marmorata were more common at a height of 7 m with a frequency of 61.4–79.6% and 68.2–79.6%, respectively. The absolute number of saproxyl and anthophilic beetle species was higher in the ground layer and at the undergrowth level. These studies expand our understanding of the vertical distribution of beetles in pine forests of the temperate zone of European Russia.
1. Introduction
Among temperate ecosystems, pine forests have an outstanding richness of plant and animal communities and biodiversity [1,2,3,4]. In the territory of Europe, coniferous forests dominated by Pinus sylvestris L. are widely spread due to their high environmental and economic importance (e.g., [5,6]). Although pine forests are generally characterized by a lower biodiversity level in comparison with deciduous forests [7], P. sylvestris forms habitats suitable for a large number of various organisms (e.g., [8,9]).
Forest systems are characterized by a high and spatially structured diversity of habitats [10,11,12,13,14,15,16,17]. In recent years, the distribution of insects has been actively studied along vertical gradients in forests of various zones, from tropical forests to temperate forests [18,19,20,21,22,23]. In floodplain forests in Germany, alpha and beta diversity of both rare and common beetle species in undergrowth and canopy differed significantly. It was shown that the species richness is higher in the undergrowth layer than in the forest canopy for common species, but it is lower for rare beetle species [24]. In a Sulawesi rainforest, 56% more common species out of 701 Coleoptera showed no preference for layers. However, there were species expressing obvious preferences for a certain height, 39% out of them were specific to the forest canopy and only 5% for the ground layer [25]. At the height of 10.5 m, the abundance, species richness and biomass of Cetoniinae were higher in trap set. In this case, traps should be located at least 4.5 m above the ground [26]. The average number of Dicerca divaricata (Buprestidae) specimens in traps with attractants was statistically significantly lower in the undergrowth layer than in any other positions of traps [27]. Collections of Monochamus galloprovincialis (Cerambycidae) individuals in traps located in tree crowns were larger than in traps situated at low heights [28].
Traps placed in the canopy of deciduous forest at the height of 13 m caught significantly more specimens of Agrilus planipennis (Buprestidae) than those placed at the ground level [29]. The Ips duplicatus (Curculionidae) abundance in pheromone traps was higher at the height of 1.5–2.0 m than in traps at the ground level or at the height of 3.5–4.0 m [30]. Using bait traps located at various heights, Miller et al. [31] found certain preferences for many Coleoptera species. Among them, Cnestus mutilatus (Curculionidae), Neoclytus scutellaris, and Monochamus titillator (Cerambycidae) confined to heights of 18–23 m, while Xylosandrus crassiusculus, Dendroctonus terebrans (Curculionidae) and Neoclytus acuminatus (Cerambycidae) were more frequent at the height of 0.3–0.5 m. In forest systems of the Eastern Canada, the Cerambycidae diversity was higher in the canopy than in the undergrowth layer, whereas their abundance was higher in the undergrowth layer compared with the canopy layer [32]. In the understory layer of beech–fir and oak forests of France, there was a higher abundance and richness of Coleoptera species [33]. In temperate deciduous forests in Japan, the abundance and composition of flying Coleoptera varied depending on layers and seasons: Elateridae were most common in the lower layer, while Attelabidae and Cantharidae were most common in the upper layer of the forest [34].
In this paper, we used beer-baited traps, placed at various heights in the tree canopy, to study the occurrence frequency, abundance and diversity of Coleoptera species along the vertical gradient in pine forests in the Central European Russia.
2. Materials and Methods
2.1. Research Area
The study was conducted in the Republic of Mordovia (Temnikov district, Mordovia State Nature Reserve: 54.42–54.56° N, 43.04–43.36° E) (Figure 1). The studied protected area covers 321.62 km2. According to the natural zoning, the study area is located on the border of the zone of coniferous–deciduous forests and the forest-steppe. In the Mordovia State Nature Reserve, forests cover 89.3% of the entire protected area. This forest type represents the largest refugium for threatened insects in the region. Thus, pine forests provide a high diversity of habitats [35,36].
The study was conducted in the green-moss pine forest. Pinus sylvestris L. forms the canopy layer. The understory layer is quite thin, being represented by Betula pendula Roth, and singular trees of Picea abies (L.) H.Karst. and Populus tremula L. In the undergrowth layer, the following shrubs are represented: Sorbus aucuparia L., Rubus idaeus L. and Frangula alnus Mill. with a total percent cover of ca. 35%. The ground layer vegetation is dominated by Convallaria majalis L. (ca. 45%) with participation of Calamagrostis epigejos (L.) Roth (ca. 5–7%). Other vascular plants of the ground layer are Dryopteris carthusiana (Vill.) H.P.Fuchs, Fragaria vesca L., Melampyrum pratense L., Vaccinium myrtillus L., Anthoxanthum odoratum L., Galium mollugo L., Agrostis gigantea Roth, Lysimachia europaea (L.) U.Manns & Anderb., Luzula pilosa (L.) Willd. and Calamagrostis arundinacea (L.) Roth.
2.2. Sampling Procedures
Coleoptera were collected from May to September 2021, a period when activity of insects is the highest. In each fragment of a pine forest, we collected samples at four heights. In this case, traps were located at a distance of no more than 15 m from each other. Between each fragment of a forest, where four traps were located at different heights, there was a certain distance indicated below. This distance between sample stations was an attempt to ensure an independence between samples. In total, there were five such forest fragments of the pine forest. Each fragment of a forest was a separate habitat. Each part of the forest was located no less than 0.8 km from each other and by representing an independent forest site. This distance is needed to obtain a representative sampling size, since it was previously demonstrated that the distance between traps should be at least 0.3 km [37].
In each sampling site, a set of four traps was installed on various heights (1.5, 3, 7 and 12 m above the ground) on branches of trees. We selected these heights to facilitate the manual installation of traps without the need to climb trees or to use special techniques or equipment to install traps. To avoid a possible edge effect, we installed traps inside the forest areas. The total sampling was equal to 176 trap exposures, where there were 9 repetitions at each height (while there were eight repetitions in one plot).
All collections were conducted by traps made by authors. Each trap consisted of a 5 L plastic container with an opening on one side at a distance of 10 cm from the bottom [37]. For bait, we used the beer mixed with honey and sugar. The collected insects in plastic bags containing 70% alcohol were delivered from forest sites to the laboratory. Then, the collected samples were sorted and stored in alcohol. In total, 15,117 specimens were studied.
2.3. Identification and Taxonomic Status of Samples
Coleoptera families were classified according to Bouchard and co-authors [38], with additions [39]. We took into account changes from the Catalog of Palaearctic Coleoptera [40,41,42,43,44,45,46], publications of Robertson and co-authors [47] for Cucujoidea, and Alonso-Zarazaga and co-authors [48] for Curculionoidea. The nomenclature of beetles was standardized according to the publications cited above, with addition of the Catalog of Palaearctic Coleoptera [49,50]. Years of description for some species are used according to Bousquet [51].
2.4. Data Analysis
We analyzed the number of trapped beetles per exposure time. Exposure time represented the period (counted in days) between installing a trap a taking the samples to be analyzed; we collected a total of 44 exposure times for each height value. The occurrence frequency is considered the ratio of the number of samples in which a taxon is present to the total number of samples (counted in %). Saproxylic species were identified based on approaches adopted in some publications [52,53,54,55]. We classified anthophilic beetles based on our personal long-term observations.
The Jaccard index was used to assess the similarity of species composition between applied heights. Those insects that could not be identified to the species levels were discarded from the analyses. The diversity index of Shannon and the dominance index of Simpson were calculated based on the collected data [56,57].
All statistical analyses were performed using PAST 4.07 [58]. The ordination was made using PCA (principal component analysis). Using this technique, we defined major gradients in the spatial arrangement of the studied species selected for analysis. We used the species abundance as response variables in PCA. For interpretation of the ordination axes, groups of the used height were plotted onto the PCA ordination diagram as supplementary environmental data. In the analysis, we included species represented by at least 100 specimens collected over the sampling period. The coefficient of determination (R2, or R-squared) was used according to Moore et al. [59].
3. Results
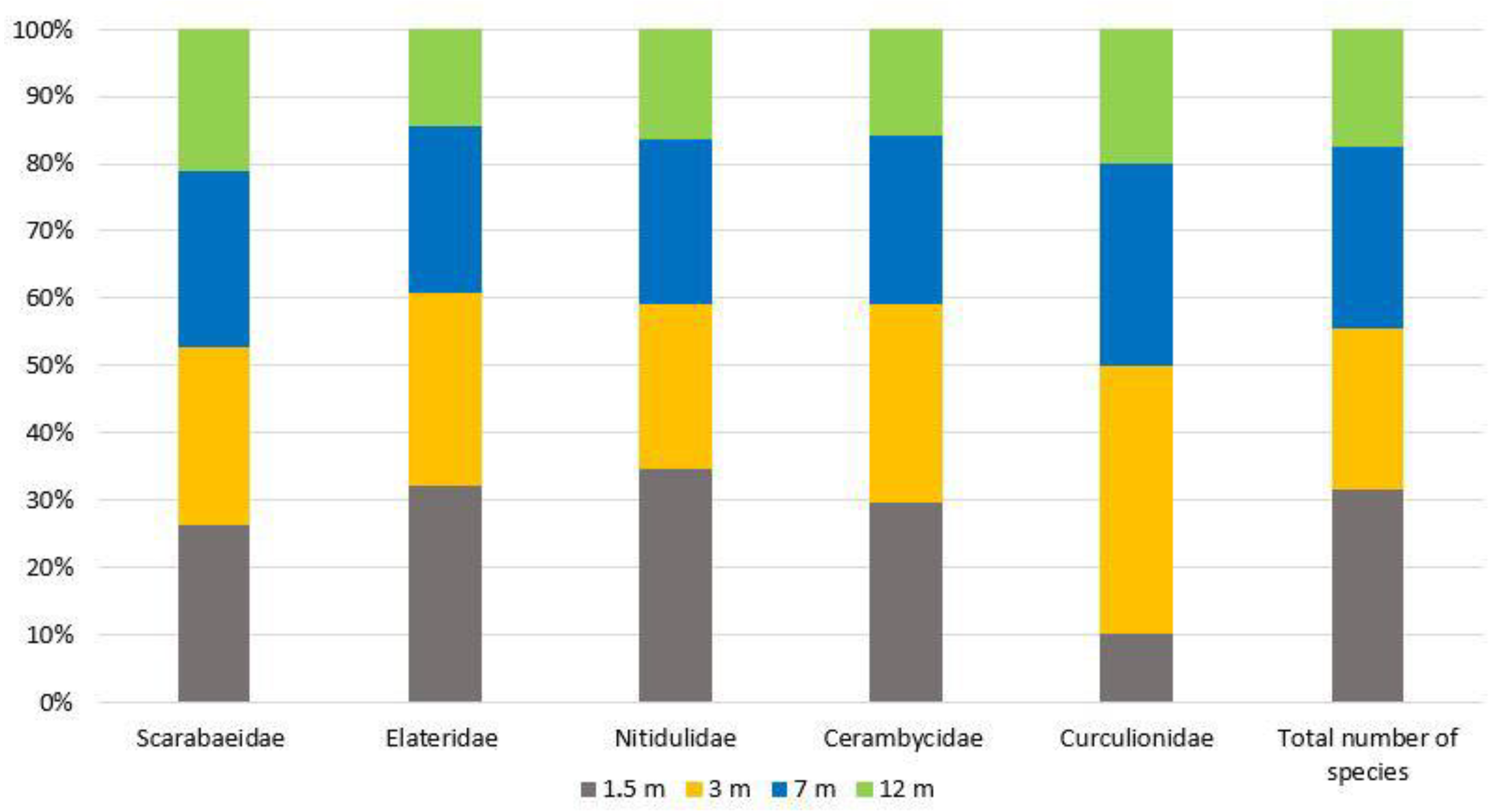
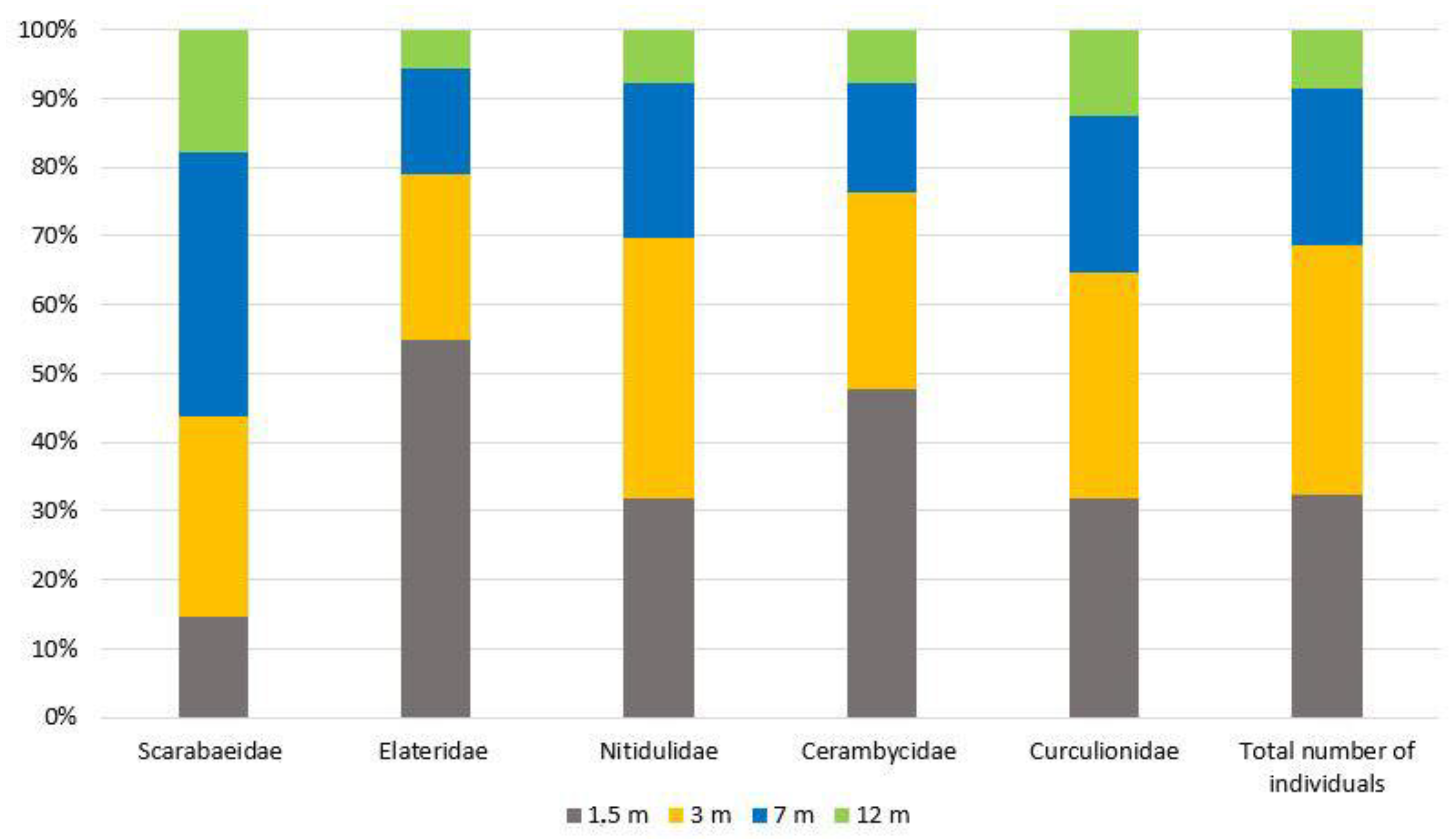
As a result of the conducted research, 125 beetle species from 36 families (Appendix A) were identified. Some specimens from the families of Staphylinidae, Nitidulidae, Throscidae, Scraptiidae, Laemophloeidae and Scirtidae were not identified to the species level. The highest species richness was recorded in Nitidulidae and Cerambycidae (19 species each), Elateridae (13 species), Curculionidae (7 species) and Scarabaeidae (6 species) (Figure 2 and Figure 3). In total, these five families, along with the family Staphylinidae, include 95.8% of all identified specimens.
Cryptarcha strigata (Fabricius, 1787) (5809 specimens in total), Glischrochilus grandis (Tournier, 1872) (3028 specimens), Protaetia marmorata (Fabricus, 1792) (765 specimens), Rhagium mordax (De Geer, 1775) (615 specimens) and Glischrochilus hortensis (Geoffroy, 1785) (519 specimens) had the highest number of specimens in traps.
Twenty-four species from 11 families were common to all studied heights (Gnathoncus buyssoni Auzat, 1917; Quedius dilatatus (Fabricius, 1787); Cetonia aurata (Linnaeus, 1758); Gnorimus variabilis (Linnaeus, 1758); P. marmorata; Dalopius marginatus (Linnaeus, 1758); Attagenus schaefferi (Herbst, 1792); Ctesias serra (Fabricius, 1792); Trogoderma glabrum (Herbst, 1783); Thanasimus femoralis (Zetterstedt, 1828); C. strigata; Cryptarcha undata (G.-A. Olivier, 1790); G. grandis; G. hortensis; Glischrochilus quadripunctatus (Linnaeus, 1758); Soronia grisea (Linnaeus, 1758); Halyzia sedecimguttata (Linnaeus, 1758); Litargus connexus (Geoffroy, 1785); Leptura quadrifasciata Linnaeus, 1758; Leptura thoracica Creutzer, 1799; Lepturalia nigripes (De Geer, 1775); Pachyta quadrimaculata (Linnaeus, 1758); and Rh. mordax; Anisandrus dispar (Fabricius, 1792)).
The highest species richness (84 species) was found at the height of 1.5 m, and the smallest number of species (44) was registered at the height of 12 m (Table 1). According to the average number of specimens, the highest number of species was found at heights of 1.5 m and 3 m (in average, 120 specimens). The minimum number of specimens was recorded at the height of 12 m. The proportion of saproxylic beetles was lower at the height of 1.5 m, while at other heights it is higher (Table 1). However, the absolute number of saproxylic species was higher at the heights of 1.5 m and 3 m than at other heights. The proportion of anthophilic species was also lower at the lower height. At the height of 12 m, the number of anthophilic species was the largest (Table 1). The absolute number of saproxylic species at heights of 1.5 m and 3 m exceeded the number of saproxylic species at other heights.
The Shannon and Simpson indices show the following results. At the height of 1.5 m, the Shannon index reached its maximal value, while the Simpson index showed its minimal value. Conversely, the minimal values of the Shannon index and maximal values of the Simpson index were found at the height of 12 m. The intermediate values between these values were obtained for other heights (Appendix A).
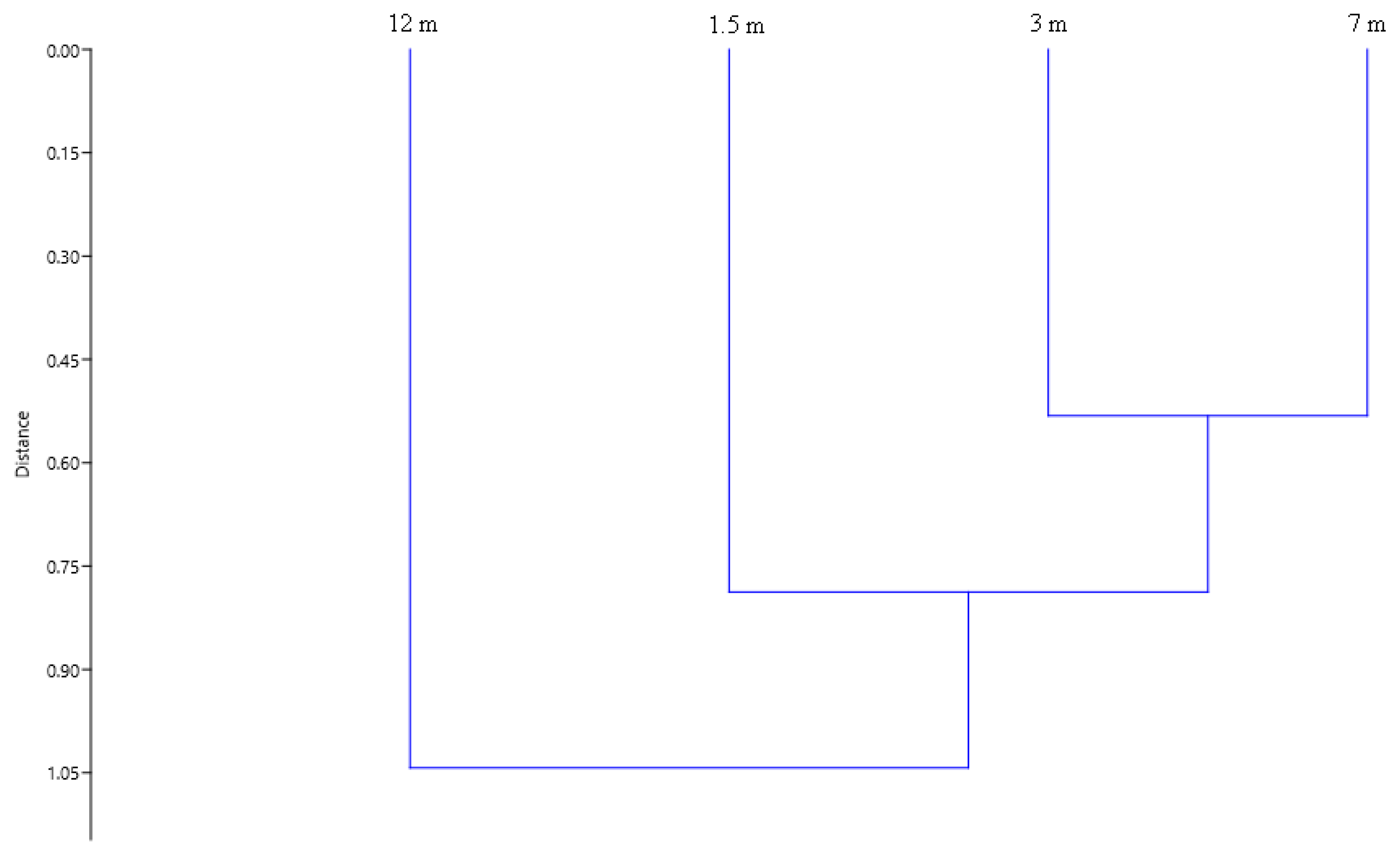
The values of the calculated Jaccard similarity index demonstrated certain differences among the studied heights (Figure 4). The highest difference values were obtained between heights of 1.5 m and 12 m. At the same time, differences were minimal between the heights of 3 m and 7 m.
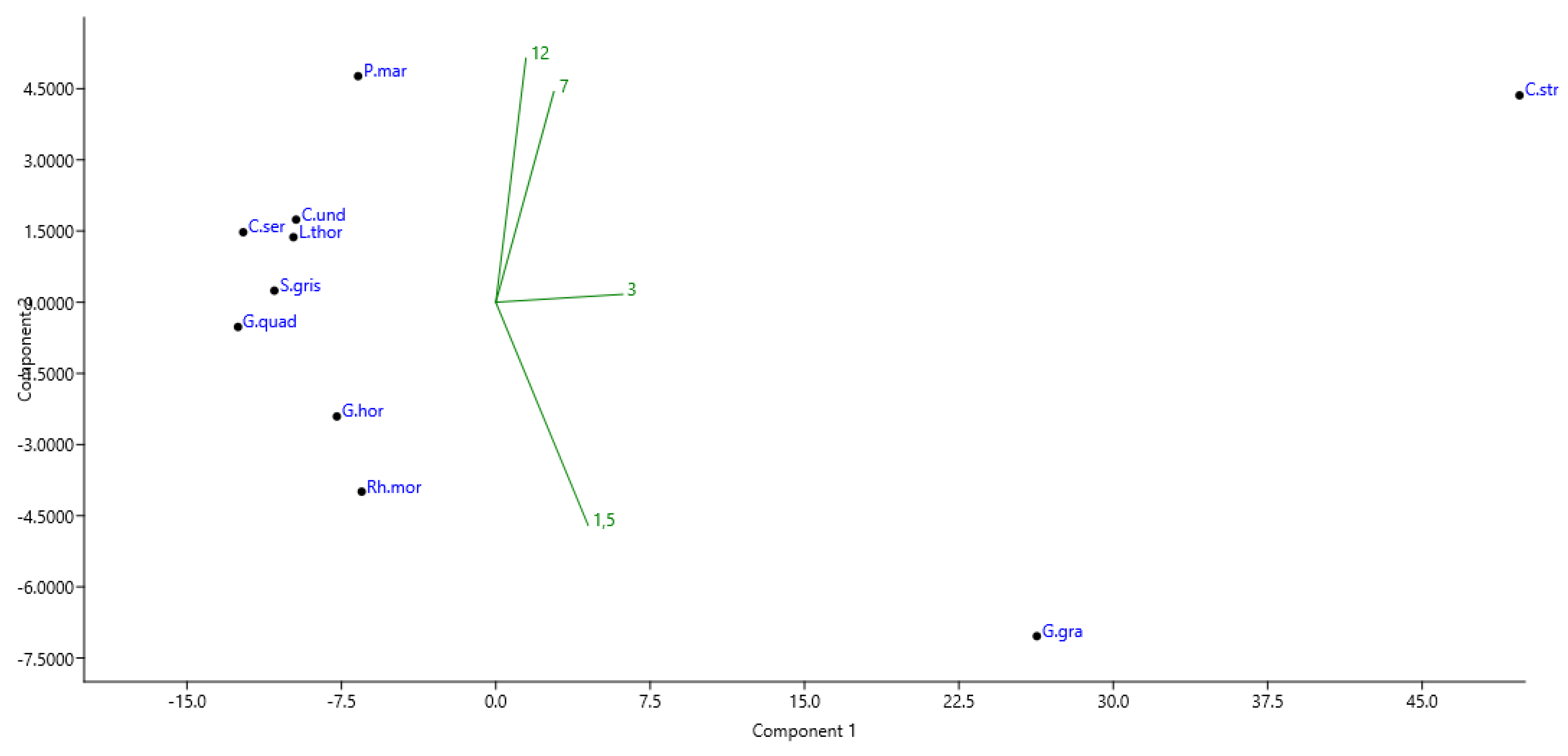
During the experiment, it was shown that only 11 Coleoptera species had a total number of more than 100 specimens (P. marmorata, C. serra, C. strigata, C. undata, G. grandis, G. hortensis, G. quadripunctatus, S. grisea, L. thoracica, Rh. mordax and A. dispar). We calculated the preferred heights for them.
The distant position of C. strigata and G. grandis along Principal Component 1 (PC1) in Figure 5 reflects their highest relative abundance in the studied pine forest. These species indeed had 3.5–92.5-fold and 1.6–47.8-fold, respectively, higher abundance at all heights in comparison with other species. According to the number of trapped specimens, C. strigata seems to prefer the 3 m height, followed by 1.5 m, while its abundance declined with an increase in the height of the trap location. The same tendency was found for another of the most-abundant species, G. grandis, which was characterized by higher abundance values at the heights of 3 m and 1.5 m; the abundance of this species was much lower at 7 m, falling to an average value of 1.32 specimens per trap at 12 m (Figure 5).
4. Discussion
This paper provides an idea of clusters of beetles captured using beer traps installed at various heights in pine forests of the temperate zone of European Russia. Previously, it was widely believed that temperate forests are less stratified than tropical forests because of the smaller structural differences and seasonal changes in climatic conditions that force many species to migrate between layers [60]. However, subsequent studies, as well as our research, have shown that beetle communities in the canopy differ from communities in the undergrowth. Longhorn and scarab beetles were studied by Touroult and Dalens [61] using bait traps placed at three various heights: 3–5 m (undergrowth), 10–15 m and 25–30 m (under canopy). It was found that, for longhorn beetles, the abundance is higher in the undergrowth, while the richness is much higher in the canopy. For scarab beetles, abundance and richness are the same in all three layers. In temperate deciduous forests in Japan, beetle abundance was higher in the canopy relative to the understory [62]. In Ontario (Canada), a higher abundance of Coleoptera was found in the undergrowth of coniferous forest [32].
In the European part of Russia, the species richness of Coleoptera falling into traps is significant. The number of species from this order actively flying to beer bait is extremely high and usually amounts to more than 90% of the total of Coleoptera specimens [63]. In these studies, we obtained similar results. As in other studies [37,63], this bait attracts many Coleoptera species. However, there are species that can accidentally be caught by the trap, which was demonstrated previously [63]. However, the number of such “accidental” species is small and usually does not exceed 10%.
Our data showed that the abundance and species richness of Coleoptera in pine forests are higher if the trap is set at heights of 1.5 m and 3 m. At the same time, the Shannon index was higher, and the Simpson index was lower at the ground layer than in tree crowns (at heights of 7 m and 12 m). The species richness at the height of 1.5 m was considerable. Similar results were obtained in the studies of Ulyshen and Hanula [64]. Therefore, in the crowns of trees of temperate pine forests, there is little species richness and the dominance of one or more species.
The found high richness is also consistent with the life habits of some Coleoptera families, for example, with their nutrition and reproduction, as well as with the interactions in which they participate, such as predation and competition [60]. There are some families and species that constantly fall into traps. For example, the families Elateridae, Cerambycidae, Staphylinidae and Curculionidae, by number, prevailed at the height of 1.5 and 3 m, i.e., closer to the soil and at the level of the undergrowth. In other studies of vertical distribution, they were also recorded in the same layer of tropical forest [65] and temperate deciduous forests [64,66]. It is likely that for some families this is due to the fact that the humidity in the undergrowth may be higher than in the canopy. For example, Staphylinidae are predominantly found in habitats with higher humidity; in addition, the understory also provides this family with resources for nutrition [67,68]. The species richness of Elateridae was almost the same at heights of 1.5, 3 and 7 m. These species are mainly saproxylic and often prefer a grassy layer and the level of undergrowth in forests [53,69]. Some species of Scolytinae (Curculionidae) were associated with heights of 7 m to 21 m, while other species are associated with the height of 1.2 m [70].
Representatives of such families as Chrysomelidae, Anthribidae, Oedemeridae, Mordellidae, Melandryidae, and Endomychidae were found only in the undergrowth. The species of these families are mainly phytophages (including anthophiles) and mycetophages [71,72,73]. Thus, herbaceous plants and woody fungi growing in the pine forest provide food resources for these species. For families such as Histeridae and Silphidae, the undergrowth also represents more favorable living conditions. Representatives of these families are necrophages, coprophages and sometimes nidicoles [73,74].
On the other hand, the species of the Cleridae family were distributed evenly over the entire vertical gradient. They are associated with woody plants and are predators of various species of xylophagous insects (Bostrichidae, Ptinidae, Scolitinae, Buprestidae and Cerambycidae); therefore, they act as important regulators of their populations [75,76]. The number and species richness of Dermestidae were almost the same at heights of 1.5, 3 and 7 m, while in the crowns of trees, these indicators decreased. The larvae of these beetles are necrophages, and imagoes are often found on flowering plants [73,77].
During the experiment, 11 species were represented by more than 100 specimens (P. marmorata, C. serra, C. strigata, C. undata, G. grandis, G. hortensis, G. quadripunctatus, S. grisea, L. thoracica, Rh. mordax and A. dispar). The average occurrence frequency of these species ranged from 11 to 100%. These species have a wide range, and they are distributed in the most diverse types of forests, forest parks and squares [78,79,80,81,82]. For C. strigata, it is shown that the abundance of the species is maximal at heights of 1.5 and 3 m with the greatest occurrence frequency at the same heights. Imagoes of this species live near the flowing fermenting juice of Q. robur, where its larvae develop, and are rarely caught on the leaking juice of P. tremula. It is in such habitats that these species thrive the most [83]. G. grandis is common on the decaying tree sap of birches and oaks, where the larva develops. Imago was also found on polypores and rotting berries [84,85]. Previously, a high abundance of this species was recorded in pine forests with a well-developed undergrowth of deciduous trees [83]. G. hortensis prefers lower layers of the forest (1.5 m). P. marmorata was clearly confined to the highest layer of the forest.
Rh. mordax was captured in the highest number at the height of 1.5 m. Rh. mordax larvae develop under the bark of dead coniferous and deciduous trees, and imagoes also prefer small heights for habitat in the forest [66,86]. Scolytinae beetles prefer to inhabit lower parts of the canopy layer, so most of them are trapped at the height of 2 m and lower [87,88]. In our studies, among the Scolytinae, A. dispar had the largest number in traps; its individuals clearly preferred the level of undergrowth, including in deciduous forests [66]. The saproxylic species L. thoracica did not have a clear preference in the pine forest. However, it was previously noted that mass collections of this species occur in places with a predominance in the birch stand and in the crowns of trees of various forest types [89,90]. Larvae of this species are polyphages of deciduous trees and live in dead, rotten wood of thick trunks (e.g., Populus, Betula, Tilia, Salix and Fagus) [91,92].
In our study, the relative number of saproxylic beetle species was lower at the height of 1.5 m, whereas at other heights it increased. In temperate forests, saproxylic beetles include 30% of all Coleoptera taxa [93]. Moreover, the biodiversity of such species is higher in forests with an increase in dead wood, stumps, dead trees and coarse wood debris [53,55,94,95]. For example, Vodka and Cizek [78] found that the saproxylic species richness was higher in the undergrowth than in the canopy at the edge of the forest, while the opposite situation was observed in depths of the forest. Preisser et al. [96] revealed that most of the studied families are more numerous in traps at the ground level. Previous studies [97] showed that the number of saproxyl beetle species was higher in well-illuminated forests and correlated positively with temperature variability. Therefore, exposure to the sun (high temperature) promotes the saproxyl beetle diversity due to the higher microclimatic heterogeneity within and between dead wood logs. Of considerable importance for saproxylic species in the ground layer of the forest are the composition of tree species, dead wood, fallen trees and remains of stumps. Under specific conditions, for example, the increased temperature on the forest ground layer, this can lead to an increase in the insect species richness of this forest layer [98,99,100,101]. This is confirmed in studies by Weiss et al. [12], who proved that in temperate forests, an increase in the proportion of saprophages near the ground layer is caused by the accumulation of dead organic matter on the forest floor.
Our research confirmed earlier studies in deciduous forests [66]. Differences in the vertical distribution of saproxylic species may be due to differences in bait sensitivity, behavior, dispersal ability, microclimate, ecological interactions or spatial heterogeneity of the nutrition quality and quantity [94,102]. For example, the difference in the vertical structure of the longhorn species community correlated with a high number of flowering plants, a high leaf area index, as well as with a high level of light intensity and steep relief in pine forests [32]. At the same time, a higher density of snags and the amount of felled wood are often recorded in such areas, which are important variables for the development of Cerambycidae larvae [103]. Since most adult Cerambycidae are good fliers and spend a lot of time feeding on flowers away from sites, which they inhabit, it is possible that adults were more active in searching for flowering plants (almost all Cerambycidae we found were anthophiles). In our study, in absolute terms, the number of saproxylic species at heights of 1.5 and 3 m exceeded the analogous indicator at other heights. A good herbaceous cover of perennial grasses and a shrubby tier are a good food base for anthophilic species.
Earlier, using the example of five different forest areas with a predominance of individual species of trees, it was shown that the highest species richness is observed in pine forests, and the lowest one in oak forests [83]. When comparing the results obtained with other studies in deciduous forests [66], this pattern persists. As a result, 92 Coleoptera species belonging to 26 families were identified [66]. One hundred twenty-five species from 36 families were obtained in pine forests. It is possible that a more significant biodiversity of Coleoptera in pine forests is associated with the microclimate of temperate pine forests, characteristics of undergrowth and grassy layer, humidity, sunlight penetration and other conditions. For example, in the Mediterranean pine forests in summer, the light transmission of tree crowns is expressed to a greater extent. In addition, the soil moisture in the undergrowth was lower than outside the areas where shrubs grow. Despite the soil moisture decrease, in pine forests and in treeless areas, shrubs serve as safe places for plant development due to the buffering of the microclimate in the summer period [104]. In Western Hungary, the middle part and the shrub layer of the pine forest played a key role in maintaining a specific microclimate in forests having a close canopy. Such conditions are favorable for sensitive taxa of forest specialists (for example, some forest grasses, epiphytic mosses and lichens) [105]. At the same time, deciduous forests with high crown closeness contribute to the reduction in undergrowth and poor herbaceous cover, which ultimately reduces the biodiversity of Coleoptera [66].
5. Conclusions
The research of beetles at various heights in temperate pine forests of the European Russia allowed us to find 125 species from 36 families. The highest species richness was revealed for Nitidulidae and Cerambycidae (19 species each), Elateridae (13 species), Curculionidae (7 species) and Scarabaeidae (6 species). Eleven species had the maximal abundance of individuals in traps, while twenty-four beetle species were common in all studied heights. According to the absolute and average number of specimens, the number of families and species, superiority was observed at heights of 1.5 m and 3 m. The minimal number of specimens and the smallest species richness of beetles were found at the height of 12 m. The highest differences in the Jaccard similarity index were obtained between minimal, 1.5 m, and maximal, 12 m, heights. The values of the Shannon index were higher at the ground layer compared to the tree crown, while values of the Simpson index were characterized by the opposite trend. C. strigata and G. grandis had clear height preferences (3 and 1.5 m). The specimen number of these species decreased with increasing height of the trap position. C. undata and P. marmorata were more common at a height of 7 m with a frequency of 61.4–79.6% and 68.2–79.6%, respectively. The absolute number of saproxylic and anthophilic species was higher at the ground and undergrowth levels of the forest. This study showed that there are still little data on the vertical distribution of arthropod species inhabiting various forests of the temperate zone. To better understand such regularities, we propose using a set of various methods of data collection, expand the taxonomic composition of the studied arthropod groups, and increase the sampling efforts, including the number of studied sites per research.
Author Contributions
Conceptualization, A.B.R. and L.V.E.; methodology, A.B.R. and A.K; software, A.A.K.; validation, A.B.R. and L.V.E.; formal analysis, A.B.R. and L.V.E.; investigation, A.B.R.; resources, A.B.R.; data curation, L.V.E.; writing—original draft preparation, A.B.R. and A.K; writing—review and editing, L.V.E.; visualization, L.V.E.; supervision, A.B.R.; project administration, A.B.R.; funding acquisition, A.B.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Russian Science Foundation, grant number 22-14-00026.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
Not applicable.
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table A1.
The mean number (M), total number of species (T) and the occurrence frequency of beetle specimens (O, %) collected using beer traps at various heights in temperate forests, Republic of Mordovia, European Russia.
Table A1.
The mean number (M), total number of species (T) and the occurrence frequency of beetle specimens (O, %) collected using beer traps at various heights in temperate forests, Republic of Mordovia, European Russia.
| Taxon Name | 1.5 m | 3 m | 7 m | 12 m | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T | M | O | T | M | O | T | M | O | T | M | O | |
| Carabidae | ||||||||||||
| Calathus micropterus (Duftschmid, 1812) | 1 | 0.02 | 2.27 | |||||||||
| Harpalus signaticornis (Duftschmid, 1812) | 1 | 0.02 | 2.27 | |||||||||
| Histeridae | ||||||||||||
| Gnathoncus buyssoni Auzat, 1917 | 22 | 0.50 | 36.36 | 16 | 0.36 | 29.55 | 16 | 0.36 | 18.18 | 8 | 0.18 | 11.36 |
| Margarinotus striola (C.R. Sahlberg, 1819) | 24 | 0.54 | 22.73 | 1 | 0.02 | 2.27 | ||||||
| Platysoma deplanatum (Gyllenhal, 1808) | 2 | 0.05 | 4.55 | 2 | 0.05 | 4.55 | ||||||
| Platysoma elongatum (Thunberg, 1787) | 17 | 0.39 | 27.27 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | |||
| Platysoma lineare Erichson, 1834 | 20 | 0.45 | 27.27 | 2 | 0.05 | 4.55 | ||||||
| Silphidae | ||||||||||||
| Necrodes littoralis (Linnaeus, 1758) | 7 | 0.16 | 13.64 | 6 | 0.14 | 9.09 | 4 | 0.09 | 9.09 | |||
| Nicrophorus vespilloides Herbst, 1783 | 3 | 0.07 | 2.27 | |||||||||
| Oiceoptoma thoracicum (Linnaeus, 1758) | 12 | 0.27 | 18.18 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | |||
| Staphylinidae | ||||||||||||
| Staphylinidae sp. | 775 | 17.61 | 95.45 | 291 | 6.61 | 79.55 | 151 | 3.43 | 75.00 | 22 | 0.50 | 29.55 |
| Euplectus sp. | 1 | 0.02 | 2.27 | |||||||||
| Quedius dilatatus (Fabricius, 1787) | 5 | 0.11 | 6.82 | 22 | 0.50 | 15.91 | 15 | 0.34 | 13.63 | 2 | 0.05 | 4.55 |
| Scarabaeidae | ||||||||||||
| Cetonia aurata (Linnaeus, 1758) | 3 | 0.07 | 6.82 | 4 | 0.09 | 9.09 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 |
| Gnorimus variabilis (Linnaeus, 1758) | 2 | 0.05 | 2.27 | 3 | 0.07 | 4.55 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 |
| Oxythyrea funesta (Poda von Neuhaus, 1761) | 1 | 0.02 | 2.27 | |||||||||
| Protaetia fieberi (Kraatz, 1880) | 4 | 0.09 | 9.09 | 4 | 0.09 | 9.09 | 6 | 0.14 | 9.09 | |||
| Protaetia marmorata (Fabricus, 1792) | 100 | 2.27 | 68.18 | 219 | 4.98 | 68.18 | 307 | 6.98 | 79.55 | 139 | 3.16 | 72.73 |
| Potosia cuprea volhyniensis (Gory & Percheron, 1833) | 14 | 0.32 | 22.73 | 12 | 0.27 | 25.00 | 5 | 0.11 | 11.36 | |||
| Scirtidae | ||||||||||||
| Contacyphon sp. | 1 | 0.02 | 2.27 | |||||||||
| Contacyphon padi (Linnaeus, 1758) | 1 | 0.02 | 2.27 | |||||||||
| Contacyphon pubescens (Fabricius, 1792) | 1 | 0.02 | 2.27 | |||||||||
| Throscidae | ||||||||||||
| Trixagus sp. | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | ||||||
| Elateridae | ||||||||||||
| Agrypnus murinus (Linnaeus, 1758) | 2 | 0.05 | 4.55 | |||||||||
| Ampedus balteatus (Linnaeus, 1758) | 5 | 0.11 | 9.09 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | |||
| Ampedus cinnabarinus (Eschscholtz, 1829) | 1 | 0.02 | 2.27 | |||||||||
| Ampedus nigroflavus (Goeze, 1777) | 2 | 0.05 | 4.55 | 2 | 0.05 | 4.55 | ||||||
| Ampedus pomorum (Herbst, 1784) | 2 | 0.05 | 4.55 | 3 | 0.07 | 6.82 | ||||||
| Ampedus praeustus (Fabricius, 1792) | 1 | 0.02 | 2.27 | |||||||||
| Athous subfuscus (O.F. Müller, 1764) | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | ||||||
| Dalopius marginatus (Linnaeus, 1758) | 1 | 0.02 | 2.27 | 4 | 0.09 | 4.55 | 3 | 0.07 | 4.55 | 1 | 0.02 | 2.27 |
| Denticollis borealis (Paykull, 1800) | 1 | 0.02 | 2.27 | |||||||||
| Elater ferrugineus Linnaeus, 1758 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | ||||||
| Melanotus castanipes (Paykull, 1800) | 3 | 0.07 | 6.82 | 1 | 0.02 | 2.27 | ||||||
| Prosternon tesselatum (Linnaeus, 1758) | 23 | 0.52 | 15.91 | 5 | 0.11 | 9.09 | 2 | 0.05 | 4.55 | |||
| Selatosomus aeneus (Linnaeus, 1758) | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | |||
| Buprestidae | ||||||||||||
| Dircaea quadriguttata (Paykull, 1798) | 1 | 0.02 | 2.27 | |||||||||
| Cantharidae | ||||||||||||
| Cantharis nigricans O.F. Müller, 1776 | 2 | 0.05 | 4.55 | 1 | 0.02 | 2.27 | ||||||
| Cantharis pellucida Fabricius, 1792 | 1 | 0.02 | 2.27 | |||||||||
| Rhagonycha nigripes (W. Redtenbacher, 1842) | 2 | 0.05 | 4.55 | |||||||||
| Dermestidae | ||||||||||||
| Attagenus schaefferi (Herbst, 1792) | 8 | 0.18 | 9.09 | 1 | 0.02 | 2.27 | 6 | 0.14 | 9.09 | 1 | 0.02 | 2.27 |
| Ctesias serra (Fabricius, 1792) | 24 | 0.54 | 15.91 | 73 | 1.66 | 22.73 | 81 | 1.84 | 20.45 | 37 | 0.84 | 22.73 |
| Dermestes lardarius Linnaeus, 1758 | 1 | 0.02 | 2.27 | |||||||||
| Megatoma undata (Linnaeus, 1758) | 4 | 0.09 | 9.09 | 3 | 0.07 | 6.82 | ||||||
| Trogoderma glabrum (Herbst, 1783) | 12 | 0.27 | 15.91 | 14 | 0.32 | 20.45 | 19 | 0.43 | 18.18 | 1 | 0.02 | 2.27 |
| Cleridae | ||||||||||||
| Thanasimus femoralis (Zetterstedt, 1828) | 1 | 0.02 | 2.27 | 2 | 0.05 | 4.55 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 |
| Thanasimus formicarius (Linnaeus, 1758) | 4 | 0.09 | 9.09 | |||||||||
| Melyridae | ||||||||||||
| Aplocnemus nigricornis (Fabricius, 1792) | 2 | 0.05 | 4.55 | 1 | 0.02 | 2.27 | ||||||
| Dasytes fusculus (Illiger, 1801) | 1 | 0.02 | 2.27 | |||||||||
| Dasytes niger (Linnaeus, 1761) | 1 | 0.02 | 2.27 | |||||||||
| Malachius bipustulatus (Linnaeus, 1758) | 2 | 0.05 | ||||||||||
| Erotylidae | ||||||||||||
| Dacne bipustulata (Thunberg, 1781) | 1 | 0.02 | 2.27 | |||||||||
| Triplax russica (Linnaeus, 1758) | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | ||||||
| Monotomidae | ||||||||||||
| Rhizophagus bipustulatus (Fabricius, 1792) | 1 | 0.02 | 2.27 | |||||||||
| Rhizophagus cribratus (Gyllenhal, 1827) | 1 | 0.02 | 2.27 | |||||||||
| Rhizophagus fenestralis (Linnaeus, 1758) | 8 | 0.18 | 9.09 | 9 | 0.20 | 13.63 | 6 | 0.14 | 4.55 | |||
| Nitidulidae | ||||||||||||
| Carpophilus hemipterus (Linnaeus, 1758) | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | ||||||
| Carpophilus marginellus Motschulsky, 1858 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | ||||||
| Cryptarcha strigata (Fabricius, 1787) | 1476 | 33.55 | 100 | 2074 | 47.14 | 97.72 | 1608 | 24.27 | 95.45 | 651 | 14.80 | 90.91 |
| Cryptarcha undata (G.-A. Olivier, 1790) | 75 | 1.70 | 61.36 | 162 | 3.68 | 77.27 | 123 | 2.80 | 79.55 | 63 | 1.43 | 68.18 |
| Cychramus luteus (Fabricius, 1787) | 22 | 0.50 | 18.18 | 9 | 0.20 | 11.36 | ||||||
| Cychramus variegatus (Herbst, 1792) | 2 | 0.05 | 4.55 | 1 | 0.02 | 2.27 | ||||||
| Epuraea sp. | 161 | 3.66 | 79.55 | 90 | 2.05 | 63.64 | 26 | 0.59 | 27.27 | 8 | 0.18 | 13.63 |
| Epuraea guttata (G.-A. Olivier, 1811) | 4 | 0.09 | 9.09 | 3 | 0.07 | 6.82 | 1 | 0.02 | 2.27 | |||
| Glischrochilus grandis (Tournier, 1872) | 1136 | 25.82 | 68.18 | 1345 | 30.57 | 54.55 | 489 | 11.11 | 43.18 | 58 | 1.32 | 38.64 |
| Glischrochilus hortensis (Geoffroy, 1785) | 277 | 6.30 | 56.82 | 173 | 3.93 | 56.82 | 62 | 1.41 | 27.27 | 7 | 0.16 | 13.63 |
| Glischrochilus quadriguttatus (Fabricius, 1777) | 48 | 1.09 | 6.82 | 3 | 0.07 | 6.82 | 1 | 0.02 | 2.27 | |||
| Glischrochilus quadripunctatus (Linnaeus, 1758) | 95 | 2.16 | 40.91 | 41 | 0.93 | 27.27 | 15 | 0.34 | 13.63 | 19 | 0.43 | 6.82 |
| Glischrochilus quadrisignatus (Say, 1835) | 2 | 0.05 | 4.55 | 2 | 0.05 | 4.55 | 1 | 0.02 | 2.27 | |||
| Ipidia binotata Reitter, 1875 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | ||||||
| Nitidula bipunctata (Linnaeus, 1758) | 2 | 0.05 | 2.27 | |||||||||
| Omosita depressa (Linnaeus, 1758) | 3 | 0.07 | 6.82 | |||||||||
| Omosita discoidea (Fabricius, 1775) | 2 | 0.05 | 4.55 | |||||||||
| Pityophagus ferrugineus (Linnaeus, 1761) | 1 | 0.02 | 2.27 | |||||||||
| Soronia grisea (Linnaeus, 1758) | 85 | 1.93 | 50.00 | 131 | 2.98 | 54.55 | 60 | 1.36 | 40.91 | 22 | 0.50 | 31.82 |
| Soronia punctatissima (Illiger, 1794) | 6 | 0.14 | 9.09 | 1 | 0.02 | 2.27 | ||||||
| Silvanidae | ||||||||||||
| Silvanus bidentatus (Fabricius, 1792) | 2 | 0.05 | 4.55 | |||||||||
| Cucujidae | ||||||||||||
| Pediacus depressus (Herbst, 1797) | 31 | 0.70 | 27.27 | 9 | 0.20 | 20.45 | 1 | 0.02 | 2.27 | |||
| Laemophloeidae | ||||||||||||
| Cryptolestes sp. | 1 | 0.02 | 2.27 | |||||||||
| Endomychidae | ||||||||||||
| Mycetina cruciata (Schaller, 1783) | 1 | 0.02 | 2.27 | |||||||||
| Coccinellidae | ||||||||||||
| Anatis ocellata (Linnaeus, 1758) | 3 | 0.07 | 4.55 | 4 | 0.09 | 6.82 | ||||||
| Calvia quatuordecimguttata (Linnaeus, 1758) | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | |||
| Halyzia sedecimguttata (Linnaeus, 1758) | 1 | 0.02 | 2.27 | 7 | 0.16 | 15.91 | 5 | 0.11 | 11.36 | 1 | 0.02 | 2.27 |
| Harmonia quadripunctata (Pontoppidan, 1763) | 1 | 0.02 | 2.27 | |||||||||
| Mycetophagidae | ||||||||||||
| Litargus connexus (Geoffroy, 1785) | 5 | 0.11 | 9.09 | 6 | 0.14 | 11.36 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 |
| Mycetophagus piceus (Fabricius, 1777) | 1 | 0.02 | 2.27 | |||||||||
| Mycetophagus quadripustulatus (Linnaeus, 1761) | 1 | 0.02 | 2.27 | |||||||||
| Ciidae | ||||||||||||
| Orthocis alni (Gyllenhal, 1813) | 1 | 0.02 | 2.27 | |||||||||
| Melandryidae | ||||||||||||
| Dircaea quadriguttata (Paykull, 1798) | 1 | 0.02 | 2.27 | |||||||||
| Orchesia fasciata (Illiger, 1798) | 4 | 0.09 | 6.82 | |||||||||
| Mordellidae | ||||||||||||
| Mordellistena humeralis (Linnaeus, 1758) | 1 | 0.02 | 2.27 | |||||||||
| Tomoxia bucephala A. Costa, 1854 | 1 | 0.02 | 2.27 | |||||||||
| Tenebrionidae | ||||||||||||
| Bolitophagus reticulatus (Linnaeus, 1767) | 1 | 0.02 | 2.27 | |||||||||
| Pseudocistela ceramboides (Linnaeus, 1758) | 1 | 0.02 | 2.27 | |||||||||
| Oedemeridae | ||||||||||||
| Chrysanthia geniculata W.L.E. Schmidt, 1846 | 1 | 0.02 | 2.27 | |||||||||
| Chrysanthia viridissima (Linnaeus, 1758) | 5 | 0.11 | 6.82 | |||||||||
| Oedemera lurida (Marsham, 1802) | 1 | 0.02 | 2.27 | |||||||||
| Pythidae | ||||||||||||
| Pytho depressus (Linnaeus, 1767) | 1 | 0.02 | 2.27 | |||||||||
| Pyrochroidae | ||||||||||||
| Schizotus pectinicornis (Linnaeus, 1758) | 1 | 0.02 | 2.27 | |||||||||
| Anthicidae | ||||||||||||
| Notoxus monoceros (Linnaeus, 1761) | 1 | 0.02 | 2.27 | |||||||||
| Scraptiidae | ||||||||||||
| Anaspis sp. | 3 | 0.07 | 6.82 | |||||||||
| Cerambycidae | ||||||||||||
| Anastrangalia reyi (Heyden, 1889) | 1 | 0.02 | 2.27 | |||||||||
| Aromia moschata (Linnaeus, 1758) | 2 | 0.05 | 4.55 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | |||
| Cortodera femorata (Fabricius, 1787) | 1 | 0.02 | 2.27 | |||||||||
| Dinoptera collaris (Linnaeus, 1758) | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | ||||||
| Leptura quadrifasciata Linnaeus, 1758 | 34 | 0.77 | 31.82 | 17 | 0.39 | 20.45 | 9 | 0.20 | 6.82 | 1 | 0.02 | 2.27 |
| Leptura thoracica Creutzer, 1799 | 111 | 2.52 | 38.64 | 128 | 2.91 | 40.91 | 116 | 2.64 | 34.09 | 77 | 1.75 | 36.36 |
| Lepturalia nigripes (De Geer, 1775) | 5 | 0.11 | 9.09 | 2 | 0.05 | 4.55 | 2 | 0.05 | 4.55 | 1 | 0.02 | 2.27 |
| Molorchus minor (Linnaeus, 1758) | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | ||||||
| Necydalis major Linnaeus, 1758 | 5 | 0.11 | 6.82 | 3 | 0.07 | 4.55 | 4 | 0.09 | 9.09 | |||
| Obrium cantharinum (Linnaeus, 1767) | 1 | 0.02 | 2.27 | |||||||||
| Pachyta quadrimaculata (Linnaeus, 1758) | 4 | 0.09 | 9.09 | 8 | 0.18 | 13.63 | 4 | 0.09 | 6.82 | 1 | 0.02 | 2.27 |
| Phymatodes testaceus (Linnaeus, 1758) | 1 | 0.02 | 2.27 | |||||||||
| Plagionotus detritus (Linnaeus, 1758) | 1 | 0.02 | 2.27 | |||||||||
| Rhagium inquisitor (Linnaeus, 1758) | 10 | 0.23 | 9.09 | 6 | 0.14 | 9.09 | 1 | 0.02 | 2.27 | |||
| Rhagium mordax (De Geer, 1775) | 389 | 8.84 | 52.27 | 169 | 3.84 | 40.91 | 46 | 1.05 | 34.09 | 11 | 0.25 | 15.91 |
| Rutpela maculata (Poda von Neuhaus, 1761) | 1 | 0.02 | 2.27 | |||||||||
| Stenocorus meridianus (Linnaeus, 1758) | 1 | 0.02 | 2.27 | |||||||||
| Stenurella melanura (Linnaeus, 1758) | 1 | 0.02 | 2.27 | |||||||||
| Stictoleptura maculicornis (De Geer, 1775) | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | 1 | 0.02 | 2.27 | |||
| Chrysomelidae | ||||||||||||
| Cryptocephalus flavipes Fabricius, 1781 | 1 | 0.02 | 2.27 | |||||||||
| Phyllotreta atra (Fabricius, 1775) | 1 | 0.02 | 2.27 | |||||||||
| Anthribidae | ||||||||||||
| Gonotropis dorsalis (Gyllenhal, 1813) | 1 | 0.02 | 2.27 | |||||||||
| Tropideres albirostris (Schaller, 1783) | 2 | 0.05 | 4.55 | |||||||||
| Nemonychidae | ||||||||||||
| Cimberis attelaboides (Fabricius, 1787) | 1 | 0.02 | 2.27 | 2 | 0.05 | 4.55 | 6 | 0.14 | 11.36 | |||
| Curculionidae | ||||||||||||
| Anisandrus dispar (Fabricius, 1792) | 149 | 3.39 | 20.45 | 152 | 3.45 | 25.00 | 104 | 2.36 | 15.91 | 58 | 0.32 | 11.36 |
| Anthonomus phyllocola (Herbst, 1795) | 1 | 0.02 | 2.27 | |||||||||
| Hylastes brunneus (Erichson, 1836) | 1 | 0.02 | 2.27 | |||||||||
| Phyllobius argentatus (Linnaeus, 1758) | 1 | 0.02 | 2.27 | |||||||||
| Polydrusus cervinus (Linnaeus, 1758) | 1 | 0.02 | 2.27 | |||||||||
| Tomicus minor (Hartig, 1834) | 1 | 0.02 | 2.27 | |||||||||
| Xyleborinus saxesenii (Ratzeburg, 1837) | 1 | 0.02 | 2.27 | |||||||||
| Total of individuals | 5281 | 5273 | 3335 | 1228 | ||||||||
References
- Brygadyrenko, V.V. Effect of canopy density on litter invertebrate community structure in pine forests. Ekológia 2016, 35, 90–102. [Google Scholar] [CrossRef] [Green Version]
- Connor, S.E.; Araújo, J.; Boski, T.; Gomes, A.; Gomes, S.D.; Leira, M.; Freitas, M.C.; Andrade, C.; Morales-Molino, C.; Franco-Múgica, F.; et al. Drought, fire and grazing precursors to large-scale pine forest decline. Divers. Distrib. 2021, 27, 1138–1151. [Google Scholar] [CrossRef]
- Popkova, T.V.; Zryanin, V.A.; Ruchin, A.B. The ant fauna (Hymenoptera: Formicidae) of the Mordovia State Nature Reserve, Russia. Nat. Conserv. Res. 2021, 6, 45–57. [Google Scholar] [CrossRef]
- Ivanova, N.V.; Shashkov, M.P.; Shanin, V.N. Study of pine forest stand structure in the Priosko-Terrasny State Nature Biosphere Reserve (Russia) based on aerial photography by quad-rocopter. Nat. Conserv. Res. 2021, 6, 1–14. [Google Scholar] [CrossRef]
- Ganopoulos, I.; Aravanopoulos, F.; Madesis, P.; Pasentsis, K.; Bosmali, I.; Ouzounis, C.; Tsaftaris, A. Taxonomic identification of Mediterranean pines and their hybrids based on the high resolution melting (HRM) and trnL approaches: From cytoplasmic inheritance to timber tracing. PLoS ONE 2013, 8, e60945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houston Durrant, T.; de Rigo, D.; Caudullo, G. Pinus sylvestrisin Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; pp. 132–133. [Google Scholar]
- Horák, J.; Materna, J.; Halda, J.P.; Mladenović, S.; Bogusch, P.; Pech, P. Biodiversity in remnants of naturalmountain forests under conservation-oriented management. Sci. Rep. 2019, 9, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lust, N.; Muys, B.; Nachtergale, L. Increase of biodiversity in homogeneous Scots pine stands by anecologically diversified management. Biodivers. Conserv. 1998, 7, 249–260. [Google Scholar] [CrossRef]
- Urbanavichus, G.P.; Urbanavichene, I.N.; Vondrák, J.; Ismailov, A.B. Epiphytic lichen biota of Prielbrusie National Park (Northern Caucasus, Russia). Nat. Conserv. Res. 2021, 6, 77–94. [Google Scholar] [CrossRef]
- Filimonova, L.V. Vegetation dynamics in the Kostomuksha State Nature Reserve (Russia) and surroundings against changes in the natural environment during the Holocene. Nat. Conserv. Res. 2021, 6 (Suppl. S1), 98–115. [Google Scholar] [CrossRef]
- Başkent, E.Z. Assessment and valuation of key ecosystem services provided by two forest ecosystems in Turkey. J. Environ. Manag. 2021, 285, 112135. [Google Scholar] [CrossRef]
- Weiss, M.; Didham, R.K.; Procházka, J.; Schlaghamerský, J.; Basset, Y.; Odegaard, F.; Tichechkin, A.; Schmidl, J.; Floren, A.; Curletti, G.; et al. Saproxylic beetles in tropical and temperate forests—A standardized comparison of vertical stratification patterns. For. Ecol. Manag. 2019, 444, 50–58. [Google Scholar] [CrossRef]
- Teshome, M.; Asfaw, Z.; Mohammed, M. Pattern of functional diversity along the elevation gradient in the dry evergreen Afromontane forest of Hararghe Highland, Southeast Ethiopia. Biosyst. Divers. 2020, 28, 257–264. [Google Scholar] [CrossRef]
- Sabattini, J.A.; Sabattini, R.A.; Cian, J.C.; Sabattini, I.A. Carbon stock in subtropical native forests in a South American Protected Area. Nat. Conserv. Res. 2021, 6, 66–79. [Google Scholar] [CrossRef]
- Ivanov, D.G.; Kotlov, I.P.; Minayeva, T.Y.; Kurbatova, J.A. Estimation of carbon dioxide fluxes on a ridge-hollow bog complex using a high resolution orthophotoplan. Nat. Conserv. Res. 2021, 6, 16–28. [Google Scholar] [CrossRef]
- Kim, J. Subdivision design and landscape structure: Case study of The Woodlands, Texas, US. Urban For. Urban Green. 2019, 38, 232–241. [Google Scholar] [CrossRef]
- Kharitonova, A.O.; Kharitonova, T.I. The effect of landscape pattern on the 2010 wildfire spread in the Mordovia State Nature Reserve, Russia. Nat. Conserv. Res. 2021, 6, 29–41. [Google Scholar] [CrossRef]
- Kirstová, M.; Pyszko, P.; Šipoš, J.; Drozd, P.; Kočárek, P. Vertical distribution of earwigs (Dermaptera: Forficulidae) in a temperate lowland forest, based on sampling with a mobile aerial lift platform. Entomol. Sci. 2017, 20, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Giovanni, F.; Mei, M.; Cerretti, P. Vertical stratification of selected Hymenoptera in a remnant forest of the Po Plain (Italy, Lombardy) (Hymenoptera: Ampulicidae, Crabronidae, Sphecidae). Fragm. Entomol. 2017, 49, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, L.; Dvořáková, K.; Oboňa, J.; Ruchin, A.B. Selected Diptera families caught with beer traps in the Republic of Mordovia (Russia). Nat. Conserv. Res. 2020, 5, 65–77. [Google Scholar] [CrossRef]
- Ruchin, A.B. Seasonal dynamics and spatial distribution of lepidopterans in selected locations in Mordovia, Russia. Biodiversitas 2021, 22, 2569–2575. [Google Scholar] [CrossRef]
- Urban-Mead, K.R.; Muñiz, P.; Gillung, J.; Espinoza, A.; Fordyce, R.; van Dyke, M.; McArt, S.H.; Danforth, B.N. Bees in the trees: Diverse spring fauna in temperate forest edge canopies. For. Ecol. Manag. 2021, 482, 118903. [Google Scholar] [CrossRef]
- Makarkin, V.N.; Ruchin, A.B. Materials on the Neuroptera and Raphidioptera fauna in Mordovia and adjacent regions of European Russia. Proc. Mordovia State Nat. Reserve 2020, 24, 161–181. [Google Scholar]
- Haack, N.; Borges, P.A.V.; Grimm-Seyfarth, A.; Schlegel, M.; Wirth, C.; Bernhard, D.; Brunk, I.; Henle, K.; Pereira, H.M. Response of common and rare beetle species to tree species and vertical stratification in a floodplain forest. Insects 2022, 13, 161. [Google Scholar] [CrossRef]
- Davis, A.J.; Sutton, S.L.; Brendell, M.J.D. Vertical distribution of beetles in a tropical rainforest in Sulawesi: The role of the canopy in contributing to biodiversity. Sepilok Bull. 2011, 13, 59–83. [Google Scholar]
- Puker, A.; Ferreira, K.R.; Correa, C.M.A. Sampling flower chafer beetles (coleoptera: Cetoniidae) in the Amazon rainforest: The role of bait types and trap installation heights get access arrow. Environ. Entomol. 2020, 49, 1096–1104. [Google Scholar] [CrossRef]
- Sweeney, J.; Hughes, C.; Webster, V.; Kostanowicz, C.; Webster, R.; Mayo, P.; Allison, J.D. Impact of horizontal edge–interior and vertical canopy–understory gradients on the abundance and diversity of bark and woodboring beetles in survey traps. Insects 2020, 11, 573. [Google Scholar] [CrossRef]
- Foit, J.; Čermák, V.; Gaar, V.; Hradil, K.; Nový, V.; Rolincová, P. New insights into the life history of Monochamus galloprovincialis can enhance surveillance strategies for the pinewood nematode. J. Pest Sci. 2019, 92, 1203–1215. [Google Scholar] [CrossRef]
- Francese, J.A.; Oliver, J.B.; Fraser, I.; Lance, D.R.; Youssef, N.; Sawyer, A.J.; Mastro, V.C. Influence of trap placement and design on capture of the emerald ash borer (Coleoptera: Buprestidae). J. Econ. Entomol. 2008, 101, 1831–1837. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, Q.H.; Wang, Y.; Liu, G.T.; Zhou, X.; Niu, J.; Schlyter, F. Catching Ips duplicatus (Sahlberg) (Coleoptera: Scolytidae) with pheromone-baited traps: Optimal trap type, colour, height and distance to infestation. Pest Manag. Sci. 2010, 66, 213–219. [Google Scholar] [CrossRef]
- Miller, D.R.; Crowe, C.M.; Sweeney, J.D. Trap height affects catches of bark and woodboring beetles (Coleoptera: Curculionidae, Cerambycidae) in baited multiple-funnel traps in Southeastern United States. J. Econ. Entomol. 2020, 113, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Vance, C.C.; Kirby, K.R.; Malcolm, J.R.; Smith, S.M. Community composition of longhorned beetles (Coleoptera: Cerambycidae) in the canopy and understorey of sugar maple and white pine stands in south-central Ontario. Environ. Entomol. 2003, 32, 1066–1074. [Google Scholar] [CrossRef]
- Bouget, C.; Brin, A.; Brustel, H. Exploring the “last biotic frontier”: Are temperate forest canopies special for saproxylic beetles? For. Ecol. Manag. 2011, 261, 211–220. [Google Scholar] [CrossRef]
- Leksono, A.S.; Takada, K.; Koji, S.; Nakagoshi, N.; Anggraeni, T.; Nakamura, K. Vertical and seasonal distribution of flying beetles in a suburban temperate deciduous forest collected by water pan trap. Insect Sci. 2005, 12, 199–206. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Khapugin, A.A. Red Data Book Invertebrates in a Protected Area of European Russia. Acta Zool. Acad. Sci. Hung. 2019, 65, 349–370. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Makarkin, N.V. Neuroptera and Raphidioptera in the Mordovia State Nature Reserve. Nat. Conserv. Res. 2017, 2, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A.; Vikhrev, N.E.; Esin, M.N. The use of simple crown traps for the insects collection. Nat. Conserv. Res. 2020, 5, 87–108. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, P.; Bousquet, Y.; Davies, A.E.; Alonso-Zarazaga, M.A.; Lawrence, J.F.; Lyal, C.H.C.; Newton, A.F.; Ried, C.A.M.; Schmitt, M.; Ślipiński, S.A.; et al. Family-group names in Coleoptera (Insecta). ZooKeys 2011, 88, 1–972. [Google Scholar] [CrossRef] [Green Version]
- Bouchard, P.; Bousquet, Y. Additions and corrections to “Family-group names in Coleoptera (Insecta)”. ZooKeys 2020, 922, 65–139. [Google Scholar] [CrossRef]
- Löbl, I.; Smetana, A. (Eds.) Catalogue of Palaearctic Coleoptera. In Curculionoidea I; Apollo Books: Stenstrup, Denmark, 2011; Volume 7, p. 373. [Google Scholar]
- Löbl, I.; Smetana, A. (Eds.) Catalogue of Palaearctic Coleoptera. In Curculionoidea II; Apollo Books: Stenstrup, Denmark, 2013; Volume 8, p. 707. [Google Scholar]
- Löbl, I.; Löbl, D. (Eds.) Catalogue of Palaearctic Coleoptera. Revised and Updated Version. Hydrophiloidea–Staphylinoidea; Brill: Leiden, The Netherlands; Boston, MA, USA, 2015; Volume 2/1, p. 1702. [Google Scholar]
- Löbl, I.; Löbl, D. (Eds.) Catalogue of Palaearctic Coleoptera. Revised and Updated Version. Scarabaeoidea–Scirtoidea–Dascilloidea–Buprestoidea–Byrrhoidea; Brill: Leiden, The Netherlands; Boston, MA, USA, 2016; Volume 3, p. 983. [Google Scholar]
- Löbl, I.; Löbl, D. (Eds.) Catalogue of Palaearctic Coleoptera. Revised and Updated Version. Archostemata–Adephaga–Myxophaga; Brill: Leiden, The Netherlands; Boston, MA, USA, 2017; Volume 1, p. 1443. [Google Scholar]
- Iwan, D.; Löbl, I. (Eds.) Catalogue of Palaearctic Coleoptera. Revised and Updated Second Edition. Tenebrionoidea; Brill: Leiden, The Netherlands; Boston, MA, USA, 2020; Volume 5, p. 945. [Google Scholar]
- Danilevsky, M. (Ed.) Catalogue of Palaearctic Coleoptera. Updated and Revised Second Edition. Chrysomeloidea I (Vesperidae, Disteniidae, Cerambycidae); Brill: Leiden, The Netherlands; Boston, MA, USA, 2020; Volume 6/1, p. 712. [Google Scholar]
- Robertson, J.; Ślipiński, A.; Moulton, M.; Shockley, F.W.; Giorgi, A.; Lord, N.P.; McKenna, D.D.; Tomaszewska, W.; Forrester, J.; Miller, K.B.; et al. Phylogeny and classification of Cucujoidea and the recognition of a new superfamily Coccinelloidea (Coleoptera: Cucujiformia). Syst. Entomol. 2015, 40, 745–778. [Google Scholar] [CrossRef]
- Alonso-Zarazaga, M.A.; Barrios, H.; Borovec, R.; Bouchard, P.; Caldara, R.; Colonnelli, E.; Gültekin, L.; Hlaváč, P.; Korotyaev, B.; Lyal, C.H.C.; et al. Cooperative Catalogue of Palaearctic Coleoptera Curculionoidea. Monogr. Electrón. SEA 2017, 8, 1–729. [Google Scholar]
- Löbl, I.; Smetana, A. (Eds.) Catalogue of Palaearctic Coleoptera. Elateroidea–Derodontoidea–Bos-trichoidea–Lymexyloidea–Cleroidea–Cucujoidea; Apollo Books: Stenstrup, Denmark, 2007; Volume 4, p. 935. [Google Scholar]
- Löbl, I.; Smetana, A. (Eds.) Catalogue of Palaearctic Coleoptera. Chrysomeloidae; Apollo Books: Stenstrup, Denmark, 2010; Volume 6, p. 924. [Google Scholar]
- Bousquet, Y. Litteratura Coleopterologica (1758–1900): A guide to selected books related to the taxonomy of Coleoptera with publication dates and notes. ZooKeys 2016, 583, 1–776. [Google Scholar] [CrossRef] [Green Version]
- Speight, M.C.D. Saproxylic invertebrates and their conservation. In Nature and Environment Series 42; Council of Europe: Strasbourg, France, 1989; pp. 1–79. [Google Scholar]
- Lachat, T.; Wermelinger, B.; Gossner, M.M.; Bussler, H.; Isacsson, G.; Müller, J. Saproxylic beetles as indicator species for dead-wood amount and temperature in European beech forests. Ecol. Indic. 2012, 23, 323–331. [Google Scholar] [CrossRef]
- Carpaneto, G.M.; Baviera, C.; Biscaccianti, A.B.; Brandmayr, P.; Mazzei, A.; Mason, F.; Battistoni, A.; Teofili, C.; Rondinini, C.; Fattorini, S.; et al. A red list of Italian saproxylic beetles: Taxonomic overview, ecological features and conservation issues (Coleoptera). Fragm. Entomol. 2015, 47, 53–126. [Google Scholar] [CrossRef]
- Papis, M.; Mokrzycki, T. Saproxylic beetles (Coleoptera) of the strictly protected area Bukowa Góra in the Roztoczański National Park. For. Res. Pap. 2015, 76, 229–239. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423, 623–659. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Ecological Diversity and Its Measurement; Chapman & Hall: London, UK, 1996; p. 179. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics soft-ware package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Moore, D.S.; Notz, W.I.; Flinger, M.A. The Basic Practice of Statistics, 6th ed.; W. H. Freeman and Company: New York, NY, USA, 2013; p. 745. [Google Scholar]
- Ulyshen, M.D. Arthropod vertical stratification in temperate deciduous forests: Implications for conservation-oriented management. For. Ecol. Manag. 2011, 261, 1479–1489. [Google Scholar] [CrossRef]
- Touroult, J.; Dalens, P.H. Beetles vertical stratification in French Guiana’ forests: Study using aerial fruit traps. In Coléoptères de Guyane; ACOREP-France: Paris, France, 2012; pp. 16–24. [Google Scholar]
- Hirao, T.; Murakami, M.; Kashizaki, A. Importance of the understory stratum to entomofaunal diversity in a temperate deciduous forest. Ecol. Res. 2009, 24, 263–272. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A. Usage of fermental traps for studying the species diversity of Coleoptera. Insects 2021, 12, 407. [Google Scholar] [CrossRef]
- Ulyshen, M.D.; Hanula, J.L. A comparison of the beetle (Coleoptera) fauna captured at two heights above the ground in a North American temperate deciduous forest. Am. Midl. Nat. 2007, 158, 260–278. [Google Scholar] [CrossRef]
- Kato, M.; Inque, T.; Hamid, A.A.; Nagamitsu, T.; Merdek, M.B.; Nona, A.R.; Itino, T.; Yamane, S.; Yumoto, T. Seasonality and vertical structure of light-attracted insect communities in a dipterocarp forest in Sarawak. Res. Popul. Ecol. 1995, 37, 59–79. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V. Vertical stratification of beetles in deciduous forest communities in the Centre of European Russia. Diversity 2021, 13, 508. [Google Scholar] [CrossRef]
- Lange, M.; Türke, M.; Pašalić, E.; Boch, S.; Hessenmöller, D.; Müller, J.; Prati, D.; Socher, S.A.; Fischer, M.; Weisser, W.W.; et al. Effects of forest management on ground-dwelling beetles (Coleoptera; Carabidae, Staphylinidae) in Central Europe are mainly mediated by changes in forest structure. For. Ecol. Manag. 2014, 329, 166–176. [Google Scholar] [CrossRef]
- Porhajašová, J.I.; Babošová, M.; Noskovič, J.; Ondrišík, P. Long-term developments and biodiversity in carabid and staphylinid (Coleoptera: Carabidae and Staphylinidae) fauna during the application of organic fertilizers under agroecosystem conditions. Pol. J. Environ. Stud. 2018, 27, 2229–2235. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Semishin, G.B. Fauna of click beetles (Coleoptera: Elateridae) in the interfluve of Rivers Moksha and Sura, Republic of Mordovia, Russia. Biodiversitas 2018, 19, 1352–1365. [Google Scholar] [CrossRef]
- Procházka, J.; Cizek, L.; Schlaghamerský, J. Vertical stratification of scolytine beetles in temperate forests. Insect Conserv. Divers. 2018, 11, 534–544. [Google Scholar] [CrossRef]
- Nikitsky, N.B.; Legalov, A.A. The ant-like stone beetles (Scydmaenidae) and the fungus weevils (Anthribidae) of the Moscow region. Euroasian Entomol. J. 2016, 15, 219–227. [Google Scholar]
- Zemoglyadchuk, A.V.; Ruchin, A.B.; Egorov, L.V. An annotated checklist of the tumbling flower beetles (Coleoptera, Mordellidae) of the Republic of Mordovia, with a short review of the family in European Russia. Entomol. Rev. 2020, 100, 771–787. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Polumordvinov, O.A. Coleoptera of the Penza region, Russia based on fermental crown trap. Biodiversitas 2021, 22, 1946–1960. [Google Scholar] [CrossRef]
- Bajerlein, D.; Matuszewski, S.; Konwersk, S. Insect succession on carrion: Seasonality, habitat preference and residency of histerid beetles (Coleoptera: Histeridae) visiting pig carrion exposed in various Forests (Western Poland). Pol. J. Ecol. 2011, 59, 787–797. [Google Scholar]
- Marković, Č.; Stojanović, A. Fauna of phloemo-xylophagous insects, their parasitoids and predators on Ulmus minor in Serbia. Biologia 2012, 67, 584–589. [Google Scholar] [CrossRef]
- Krivets, S.A.; Kerchev, I.A. Insects inhabiting the galleries of the four-eyed fir bark beetle Polygraphus proximus Blandf. (Coleoptera, Curculionidae: Scolytinae) in Siberia. Entomol. Rev. 2016, 96, 545–558. [Google Scholar] [CrossRef]
- Kadej, M. Larva and pupa of Ctesias (s. str.) serra (Fabricius, 1792) with remarks on biology and economic importance, and larval comparison of co-occurring genera (Coleoptera, Dermestidae). ZooKeys 2018, 758, 115–135. [Google Scholar] [CrossRef] [Green Version]
- Vodka, Š.; Cizek, L. The effects of edge-interior and understorey-canopy gradients on the distribution of saproxylic beetles in a temperate lowland forest. For. Ecol. Manag. 2013, 304, 33–41. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V. Fauna of longicorn beetles (Coleoptera: Cerambycidae) of Mordovia. Russ. Entomol. J. 2018, 27, 161–177. [Google Scholar] [CrossRef] [Green Version]
- Egorov, L.V.; Ruchin, A.B.; Semenov, V.B.; Semionenkov, O.I.; Semishin, G.B. Checklist of the Coleoptera of Mordovia State Nature Reserve, Russia. ZooKeys 2020, 962, 13–122. [Google Scholar] [CrossRef]
- Egorov, L.V.; Ruchin, A.B.; Semishin, G.B. Some data on the Coleoptera fauna of the Mordovia State Nature Reserve. Report 10. Proc. Mordovia State Nat. Reserve 2021, 26, 96–128. [Google Scholar]
- Ruzzier, E.; Galli, A.; Bani, L. Monitoring exotic beetles with inexpensive attractants: A case study. Insects 2021, 12, 462. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A. Seasonal activity of Coleoptera attracted by fermental crown traps in forest ecosystems of Central Russia. Ecol. Quest. 2021, 32, 37–53. [Google Scholar] [CrossRef]
- Alekseev, V.I.; Nikitsky, N.B. Rare and new for the fauna of the Baltic States beetles (Coleoptera) from the Kaliningrad Region. Acta Zool. Litu. 2008, 18, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Lasoń, A.; Holly, M. Glischrochilus grandis Tournier, 1872—New species of beetle for the Polish fauna and new data on the occurrence of genus Glischrochilus Reitter, 1873 (Coleoptera: Nitidulidae: Cryptarchinae). Acta Entomol. Sil. 2015, 23, 1–4. [Google Scholar]
- Gorshkova, V.P.; Volodchenko, A.N. The specific assemblage structure of longhorn beetles (Coleoptera, Cerambycidae) in floodplain forests of the western part of Saratov oblast. Biol. Bull. 2016, 43, 1416–1421. [Google Scholar] [CrossRef]
- Sheehan, T.N.; Ulyshen, M.D.; Horn, S.; Hoebeke, E.R. Vertical and horizontal distribution of bark and woodboring beetles by feeding guild: Is there an optimal trap location for detection? J. Pest Sci. 2019, 92, 327–341. [Google Scholar] [CrossRef]
- Holuša, J.; Fiala, T.; Foit, J. Ambrosia beetles prefer closed canopies: A case study in oak forests in Central Europe. Forests 2021, 12, 1223. [Google Scholar] [CrossRef]
- Danilevsky, M.L.; Ruchin, A.B.; Egorov, L.V. Mass collection of two rare longicorn-species (Coleoptera, Cerambycidae) in Central Russia. Humanit. Space 2019, 8, 1179–1183. [Google Scholar]
- Ruchin, A.B.; Egorov, L.V.; MacGowan, I.; Makarkin, V.N.; Antropov, A.V.; Gornostaev, N.G.; Khapugin, A.A.; Dvořák, L.; Esin, M.N. Post-fire insect fauna explored by crown fermental traps in forests of the European Russia. Sci. Rep. 2021, 11, 21334. [Google Scholar] [CrossRef]
- Sama, G. Atlas of the Cerambycidae of Europe and the Mediterranean Area. Northern, Western, Central and Eastern Europe British Isles and Continental Europe from France (Excl. Corsica) to Scandinavia and Urals. Kabourek, Zlín; Coronet Books Incorporated: London, UK, 2002; Volume 1, p. 173. [Google Scholar]
- Karpiński, L.; Szczepański, W.T.; Boldgiv, B.; Walczak, M. New data on the longhorn bee-tles of Mongolia with particular emphasis on the genus Eodorcadion Breuning, 1947 (Coleoptera, Cerambycidae). ZooKeys 2018, 739, 107–150. [Google Scholar] [CrossRef] [Green Version]
- Haeler, E.; Bergamini, A.; Blaser, S.; Ginzler, C.; Hindenlang, K.; Keller, C.; Kiebacher, T.; Kormann, U.G.; Scheidegger, C.; Schmidt, R.; et al. Saproxylic species are linked to the amount and isolation of dead wood across spatial scales in a beech forest. Landsc. Ecol. 2021, 36, 89–104. [Google Scholar] [CrossRef]
- Ekström, A.L.; Bergmark, P.; Hekkala, A.M. Can multifunctional forest landscapes sustain a high diversity of saproxylic beetles? For. Ecol. Manag. 2021, 490, 119107. [Google Scholar] [CrossRef]
- Parisi, F.; Innangi, M.; Tognetti, R.; Lombardi, F.; Chirici, G.; Marchetti, M. Forest stand structure and coarse woody debris determine the biodiversity of beetle communities in Mediterranean mountain beech forests. Glob. Ecol. Conserv. 2021, 28, e01637. [Google Scholar] [CrossRef]
- Preisser, E.; Smith, D.C.; Lowman, M.D. Canopy and ground level insect distribution in a temperate forest. Selbyana 1998, 19, 141–146. [Google Scholar]
- Lettenmaier, L.; Seibold, S.; Bässler, C.; Brandl, R.; Gruppe, A.; Müller, J.; Hagge, J. Beetle diversity is higher in sunny forests due to higher microclimatic heterogeneity in deadwood. Oecologia 2022, 198, 825–834. [Google Scholar] [CrossRef]
- Pakeman, R.J.; Stockan, J.A. Drivers of carabid functional diversity: Abiotic environment, plant functional traits, or plant functional diversity? Ecology 2014, 95, 1213–1224. [Google Scholar] [CrossRef]
- Penone, C.; Allan, E.; Soliveres, S.; Felipe-Lucia, M.R.; Gossner, M.M.; Seibold, S.; Simons, N.K.; Schall, P.; Van Der Plas, F.; Manning, P.; et al. Specialisation and diversity of multiple trophic groups are promoted by different forest features. Ecol. Lett. 2019, 22, 170–180. [Google Scholar] [CrossRef]
- Schneider, A.; Blick, T.; Köhler, F.; Pauls, S.U.; Römbke, J.; Zub, P.; Dorow, W.H.O. Animal diversity in beech forests—An analysis of 30 years of intense faunistic research in Hessian strict forest reserves. For. Ecol. Manag. 2021, 499, 119564. [Google Scholar] [CrossRef]
- Tinya, F.; Kovács, B.; Bidló, A.; Dima, B.; Király, I.; Kutszegi, G.; Lakatos, F.; Mag, Z.; Márialigeti, S.; Nascimbene, J.; et al. Environmental drivers of forest biodiversity in temperate mixed forests—A multi-taxon approach. Sci. Total Environ. 2021, 795, 148720. [Google Scholar] [CrossRef]
- Maguire, D.Y.; Robert, K.; Brochu, K.; Larrivée, M.; Buddle, C.M.; Wheeler, T.A. Vertical stratification of beetles (Coleoptera) and flies (Diptera) in temperate forest canopies. Environ. Entomol. 2014, 43, 9–17. [Google Scholar] [CrossRef]
- Gosling, D.C.L.; Gosling, N.M. An annotated list of the Cerambycidae of Michigan (Coleoptera) Part II, the subfamilies Lepturinae and Lamiinae. Great Lakes Entomol. 1977, 10, 1–37. [Google Scholar]
- Prévosto, B.; Helluy, M.; Gavinet, J.; Fernandez, C.; Balandier, P. Microclimate in Mediterranean pine forests: What is the influence of the shrub layer? Agric. For. Meteorol. 2020, 282–283, 107856. [Google Scholar] [CrossRef]
- Kovács, B.; Tinya, F.; Ódor, P. Stand structural drivers of microclimate in mature temperate mixed forests. Agric. For. Meteorol. 2017, 234–235, 11–21. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Study sites in the Mordovia State Nature Reserve, European Russia.

Figure 2.
The proportion of species in five Coleoptera families collected using beer traps at various heights (see Appendix A).
Figure 2.
The proportion of species in five Coleoptera families collected using beer traps at various heights (see Appendix A).

Figure 3.
The proportion of specimens in five Coleoptera families collected using beer traps at various heights.
Figure 3.
The proportion of specimens in five Coleoptera families collected using beer traps at various heights.

Figure 4.
The similarity of beetle species composition between four studied heights based on the Jaccard index. We applied Ward’s method and Euclidean distance (cophenetic correlation coefficient = 0.771).
Figure 4.
The similarity of beetle species composition between four studied heights based on the Jaccard index. We applied Ward’s method and Euclidean distance (cophenetic correlation coefficient = 0.771).

Figure 5.
The diagram of the principal component analysis ordination of the selected beetle species in the Republic of Mordovia (European Russia) based on the number of specimens collected at various heights of trap positions (1.5 m, 3 m, 7 m, 12 m). Designations: P.mar.—Protaetia marmorata; C.ser.—Ctesias serra; C.str.—Cryptarcha strigata; C.und.—Cryptarcha undata; G.gra.—Glischrochilus grandis; G.hort.—Glischrochilus hortensis; G.quad.—Glischrochilus quadripunctatus; S.gris.—Soronia grisea; L.thor.—Leptura thoracica; Rh.mor.—Rhagium mordax; A.dis.—Anisandrus dispar. Groups of heights of trap positions were plotted onto the ordination diagram as supplementary environmental variables.
Figure 5.
The diagram of the principal component analysis ordination of the selected beetle species in the Republic of Mordovia (European Russia) based on the number of specimens collected at various heights of trap positions (1.5 m, 3 m, 7 m, 12 m). Designations: P.mar.—Protaetia marmorata; C.ser.—Ctesias serra; C.str.—Cryptarcha strigata; C.und.—Cryptarcha undata; G.gra.—Glischrochilus grandis; G.hort.—Glischrochilus hortensis; G.quad.—Glischrochilus quadripunctatus; S.gris.—Soronia grisea; L.thor.—Leptura thoracica; Rh.mor.—Rhagium mordax; A.dis.—Anisandrus dispar. Groups of heights of trap positions were plotted onto the ordination diagram as supplementary environmental variables.

Table 1.
The main parameters of Coleoptera individuals collected using beer traps at various heights.
Table 1.
The main parameters of Coleoptera individuals collected using beer traps at various heights.
| Parameters | Height (m) | |||
|---|---|---|---|---|
| 1.5 | 3 | 7 | 12 | |
| Total of families | 28 | 24 | 17 | 18 |
| Total of individuals | 5281 | 5273 | 3335 | 1228 |
| Mean number of individuals per trap | 120 | 120 | 76 | 28 |
| Number of species (excluding unidentified insects) | 84 | 69 | 59 | 44 |
| Number of saproxylic species | 56 | 52 | 48 | 33 |
| Proportion (%) of saproxylic species | 66.7 | 75.4 | 81.4 | 75.0 |
| Number of anthophilic species | 34 | 34 | 26 | 24 |
| Proportion (%) of anthophilic species | 40.5 | 49.3 | 44.1 | 54.5 |
| Shannon index | 2.24 | 1.91 | 1.85 | 1.81 |
| Simpson index | 0.20 | 0.26 | 0.30 | 0.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A. Vertical Distribution of Beetles (Coleoptera) in Pine Forests in Central European Russia. Diversity 2022, 14, 622. https://0-doi-org.brum.beds.ac.uk/10.3390/d14080622
AMA Style
Ruchin AB, Egorov LV, Khapugin AA. Vertical Distribution of Beetles (Coleoptera) in Pine Forests in Central European Russia. Diversity. 2022; 14(8):622. https://0-doi-org.brum.beds.ac.uk/10.3390/d14080622
Chicago/Turabian StyleRuchin, Alexander B., Leonid V. Egorov, and Anatoliy A. Khapugin. 2022. "Vertical Distribution of Beetles (Coleoptera) in Pine Forests in Central European Russia" Diversity 14, no. 8: 622. https://0-doi-org.brum.beds.ac.uk/10.3390/d14080622
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.