
Genetic Diversity of Chinese Longsnout Catfish (Leiocassis longirostris) in Four Farmed Populations Based on 20 New Microsatellite DNA Markers

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
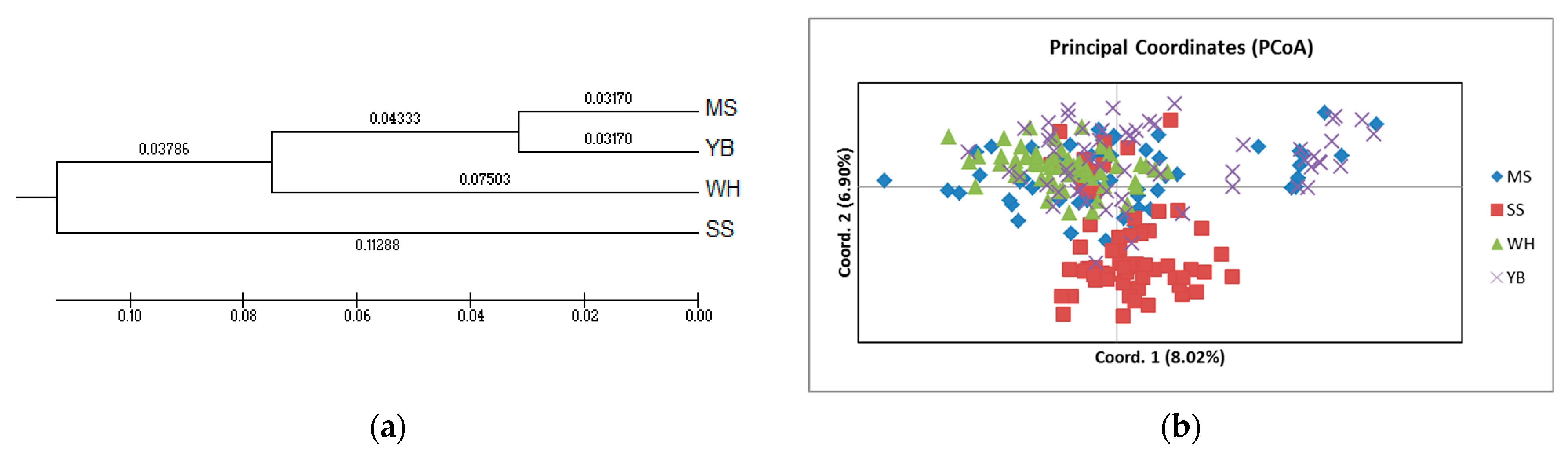
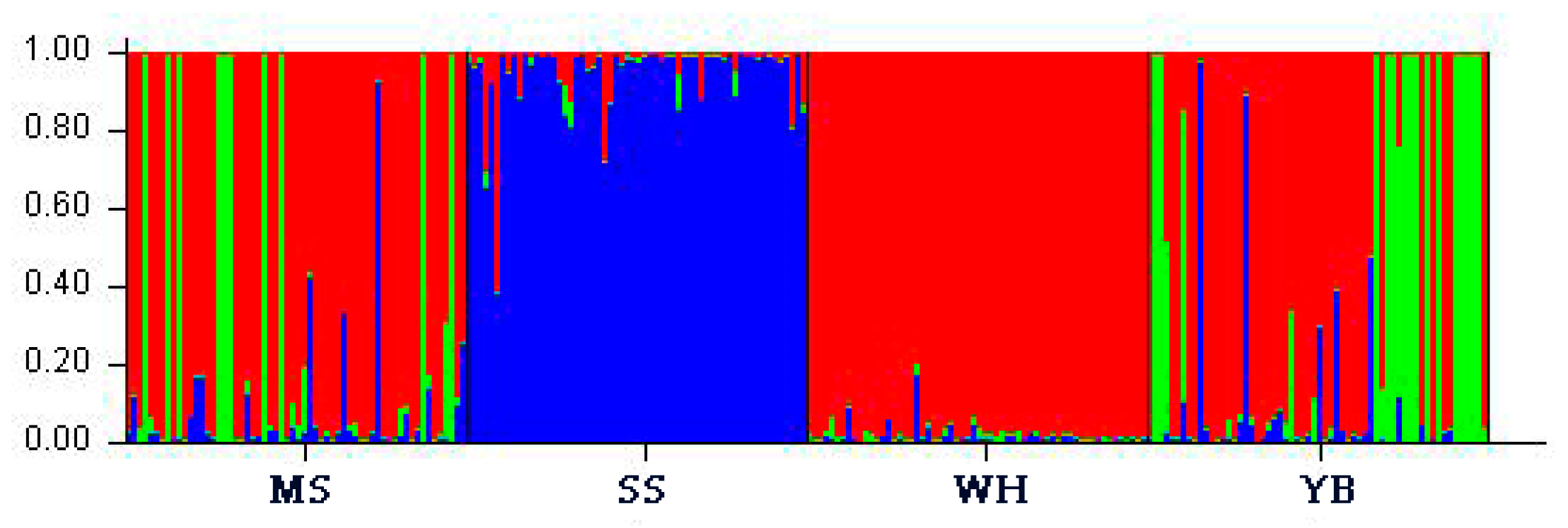
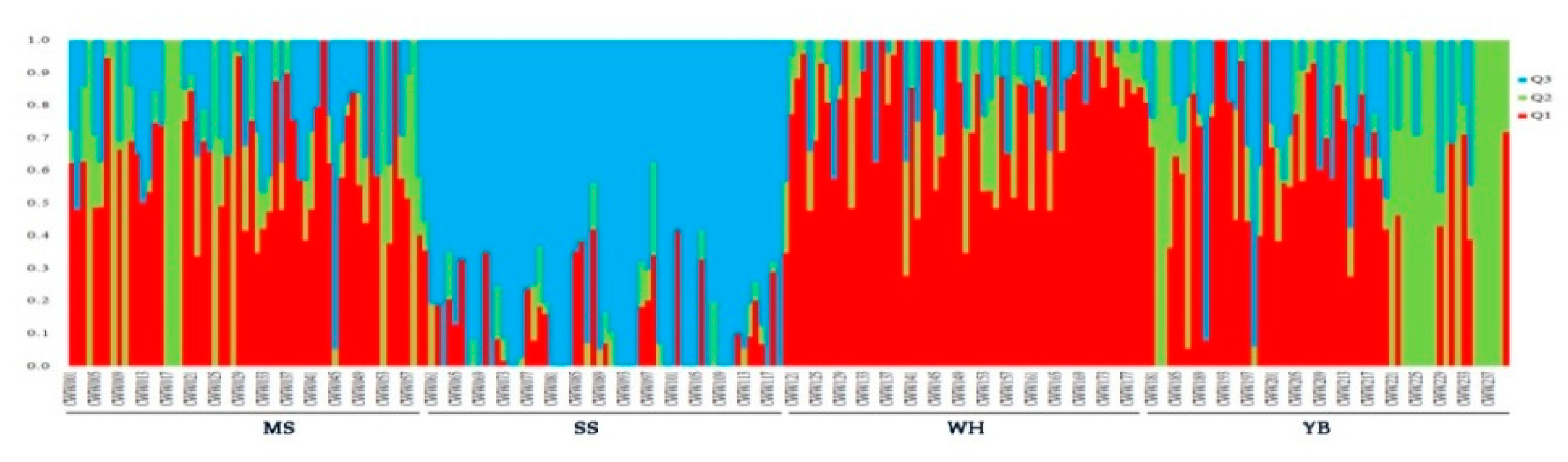
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- He, W.P.; Zhou, J.; Li, Z.; Jing, T.S.; Li, C.H.; Yang, Y.J.; Xiang, M.B.; Zhou, C.W.; Lv, G.J.; Xu, H.Y.; et al. Chromosome-level genome assembly of the Chinese longsnout catfish Leiocassis longirostris. Zool. Res. 2021, 42, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.H.; Wang, Y.J. Biological characteristics and artificial breeding techniques of Leiocassis longirostris. Fish. Guide Be Rich 2006, 24, 27–29. [Google Scholar]
- Zhang, S.L.; Sun, X.J.; Zhang, X.; Zhi, Q.J.; Su, J.T.; Liang, Y.J.; Yang, P. Analysis of meat content and muscle nutrient components in the Chinese longsnout catfish. J. Dalian Ocean Univ. 2013, 28, 335–336. [Google Scholar]
- Gao, Y.X.; Zhang, H.X.; Hu, Y.M.; Li, Z.M.; Shang, X.H.; Zhao, Y.F.; Wu, Y.N. Fatty acid content of freshwater fishes in Dongting Lake. J. Food Hyg. China 2015, 27, 6–9. [Google Scholar]
- Hubert, N.; Pepey, E.; Mortillaro, J.M.; Steinke, D.; Verdal, H.D. Mitochondrial Genetic Diversity among Farmed Stocks of Oreochromis spp. (Perciformes, Cichlidae) in Madagascar. Diversity 2021, 13, 281. [Google Scholar] [CrossRef]
- Xiao, M.; Hu, Q.; Zhao, Y.; Bao, F. Development of SNP markers in Leiocassis longirostris Günther using high-throughput sequencing. Conserv. Genet. Resour. 2020, 12, 173–176. [Google Scholar] [CrossRef]
- Zeng, S.Q. A 10-year ban on fishing in the Yangtze river has led to an ecological war. Rural. Work. Newsl. 2020, 16, 13–15. [Google Scholar]
- Wang, D.; Wu, F.X.; Song, D.D.; Gao, H.Q.; Wang, Y.L.; Wang, J.X.; Wang, S.H.; Wang, X.H.; Wei, J.X.; Zhu, M.; et al. Bureau of Fisheries, Ministry of Agriculture and Rural Affairs of the People’s Republic of China. China Fishery Statistical Yearbook, 1st ed.; China Agriculture Press: Bejing, China, 2021. [Google Scholar]
- Luo, Y.H.; Zhang, Y.Y. Studies on the storage and artificial propagation techniques of Leiocassis longirostris. Freshw. Fish. 1986, 4, 5–9. [Google Scholar]
- Liu, F.R. Discussion on the practical technology of intensive culture of Leiocassis longirostris. Mod. Fish. Inf. 2003, 18, 22–25. [Google Scholar]
- Zhu, L.; Li, Z.H.; Li, X.X.; Yang, Z.Q. Artificial breeding and culture techniques of Leiocassis longirostris. J. Aquac. 2020, 41, 54–55. [Google Scholar]
- Jenkins, S.F.; Ishengoma, E.; Rhode, C. A temporal assessment of family composition and genetic diversity in a commercial cohort of Dusky Kob, Argyrosomus japonicus, across the production cycle. Aquaculture 2020, 516, 734640. [Google Scholar] [CrossRef]
- Skaala, Ø.; Høyheim, B.; Glovera, K.; Dahlea, G. Microsatellite analysis in domesticated and wild Atlantic salmon (Salmo salar L). Allelic Divers. Identif. Individ. Aquac. 2004, 240, 131–143. [Google Scholar]
- Frost, L.A.; Evans, B.S.; Jerry, D.R. Loss of genetic diversity due to hatchery culture practices in barramundi (Lates calcarifer). Aquaculture 2007, 261, 1056–1064. [Google Scholar] [CrossRef]
- Le, W.; Shi, X.; Su, Y.; Meng, Z.; Lin, H. Loss of Genetic Diversity in the Cultured Stocks of the Large Yellow Croaker, Larimichthys crocea, Revealed by Microsatellites. Int. J. Mol. Sci. 2012, 13, 5584–5597. [Google Scholar]
- Santos, C.H.A.; Santana, G.X.; Leitão, C.S.S.; Paula-Silva, M.N.; Almeida-Val, V.M.F. Loss of genetic diversity in farmed populations of Colossoma macropomum estimated by microsatellites. Anim. Genet. 2016, 47, 373–376. [Google Scholar] [CrossRef]
- Shan, B.B.; Liu, Y.; Song, N.; Yang, C.P.; Liu, S.G.; Gao, T.X.; Sun, D.R. Parentage determination of black sea bream (Acanthopagrus schlegelii) for stock enhancement: Effectiveness and loss of genetic variation. Acta Oceanol. Sin. 2021, 40, 41–49. [Google Scholar] [CrossRef]
- Rao, F.X. Biological characteristics of Leiocassis longirostris and its artificial propagation. Henan Fish. 1994, 4, 31–32. [Google Scholar]
- Zhang, Y.G.; Luo, Q.S.; Zhong, M.C. Study on the nesting and sperm structure of Leiocassis longirostris. Acta Hydrobiol. Sin. 1993, 17, 246–250. [Google Scholar]
- Wu, Y.J. Study on the simple method of fish sperm preservation. Inland Fish. 2003, 6, 31. [Google Scholar]
- Xiao, M.S.; Cui, F.; Jian, K.; Ma, Y.H. Analysis on sequence polymorphism of the mitochondrial DNA control region and population genetic diversity of the cultivated and natural chinese longsnout catfish (Leiocassis longirostris). Acta Hydrobiol. Sin. 2013, 37, 90–99. [Google Scholar]
- Yang, G.; Xiao, M.; Yu, Y.; Xu, S. Genetic variation at mtDNA and microsatellite loci in Chinese longsnout catfish (Leiocassis longirostris). Mol. Biol. Rep. 2012, 39, 4605–4617. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.X.; Wang, X.Q.; Mo, Y.L. RAPD analysis of genetic diversity of Leiocassis longirostris. Jiangxi Fish. Sci. Technol. 2010, 2, 13–16. [Google Scholar]
- Xiao, M.S.; Xia, H.W.; Ma, Y.H. Genetic variation of the Chinese longsnout catfish Leiocassis longirostris in the Yangtze River revealed using mitochondrial DNA cytochrome b sequences. Acta Ecol. Sin. 2012, 32, 305–313. [Google Scholar] [CrossRef]
- Wang, H.Y.; Huang, W.Q. Preliminary Analysis on the Genetic Diversity in Four Populations of Leiocassis longirostris by Using Microsatellite Markers. J. Henan Agric. Sci. 2011, 40, 146–148. [Google Scholar]
- Wang, Z.; Zhou, J.; Ye, Y.; Wei, Q.; Wu, Q. Genetic structure and low-genetic diversity suggesting the necessity for conservation of the Chinese Longsnout catfish, Leiocassis longirostris (Pisces: Bagriidae). Environ. Biol. Fishes 2006, 75, 455–463. [Google Scholar] [CrossRef]
- Guo, A.; Yuan, J.; Lian, Q.; Li, M.; Gu, Z. Isolation and characterization of 20 polymorphic microsatellites loci for Xenocypris davidi based on high-throughput sequencing. Mol. Biol. Rep. 2020, 47, 8305–8310. [Google Scholar] [CrossRef]
- Xiao, M.; Yang, G. Isolation and characterization of 17 microsatellite loci for the Chinese longsnout catfish (Leiocassis longirostris). Mol. Ecol. Resour. 2009, 9, 1039–1041. [Google Scholar] [CrossRef]
- Yu, Y.; Xiao, M.; Chen, L.; Yang, G. Isolation and characterization of microsatellite loci in the longsnout catfish (Leiocassis longirostris). Aquaculture Research,. 2009, 40, 246–248. [Google Scholar] [CrossRef]
- Marshall, T.C.; Slate, J.; Kruuk LE, B.; Pemberton, J.M. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (Version 1.2): Population Genetics Software for Exact Tests and Ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol Ecol Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; Weiss, K.M. Admixture as a tool for finding linked genes and detecting that difference from allelic association between loci. Proc. Natl. Acad. Sci. USA 1988, 85, 9119–9123. [Google Scholar] [CrossRef]
- Yue, L.; Wang, Y.; Xian, W.; Zhang, H. Genetic Diversity and Population Structure of Portunus trituberculatus in Released and Wild Populations Based on Microsatellite DNA Markers from the Yangtze Estuary. Diversity 2022, 14, 374. [Google Scholar] [CrossRef]
- Sánchez-Velásqueza, J.J.; Pinedo-Bernalb, P.N.; Reyes-Floresa, L.E.; Yzásiga-Barreraa, C.; Zelada-Mázmela, E. Genetic diversity and relatedness inferred from microsatellite loci as a tool for broodstock management of fine flounder Paralichthys adspersus. Aquac. Fish. 2021, 7, 664–674. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Yang, M.O.; Zhang, F.T.; Wang, J.W. Study on the genetic structure of wild and hatchery populations of Procypris rabaudi Tchang, an endemic fish in the upper Yangtze River. Fish. Res. 2022, 245, 106–134. [Google Scholar] [CrossRef]
- Lindqvist, A.K.; Magnusson, P.K.; Balciuniene, J.; Wadelius, C.; Lindholm, E.; Alarcon-Riquelme, M.E.; Gyllensten, U.B. Chromosome-specific panels of tri- and tetranucleotide microsatellite markers for multiplex fluorescent detection and automated genotyping: Evaluation of their utility in pathology and forensics. Genome Res. 1996, 6, 1170–1176. [Google Scholar] [CrossRef]
- Souza-Shibatta, L.; Ricardo, P.C.; Pina, W.C.; Flaresso-Neto, V.; Freiria, G.A.; Kotelok-Diniz, T.; Gaglianone, M.C.; Arias, M.C.; Sofia, S.H. Isolation and characterization of microsatellite markers in two species of the neotropical Epicharis (Hymenoptera, Apidae, Centridini) genus and cross-amplification in related species. Mol. Biol. Rep. 2021, 48, 1977–1983. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Fisheries, C. Report on the development of Aquatic seed industry in China (From 1949 to 2019). China Fish. 2020, 9, 11–21. [Google Scholar]
- Bao, M.H.; Feng, J.; He, J.J.; He, X.P.; Ding, D.L. Main points of cultivation techniques for Leiocassis longirostris seedlings of the characteristic fish in the Yangtze River. China Fish. 2021, 8, 76–77. [Google Scholar]
- Shu, J.; Li, Q.; Yu, R.H.; Tian, C.Y. Microsatellites Analysis on Genetic Variation Between Wild and Cultured Populations of Pacific Abalone (Haliotis discus hannai). Period. Ocean Univ. China 2008, 38, 52–58. [Google Scholar]
- Zhang, L.X.; Ding, F.X.; Zhao, X.H.; Wang, J.Y.; Jin, C.F.; Zhang, L. Effects of sample size and gender on population genetic diversity in microsatellite analysis. Anim. Husb. Vet. Med. 2011, 43, 36–39. [Google Scholar]
- Seo, D.; Bhuiyan MS, A.; Sultana, H.; Heo, J.M.; Lee, J.H. Genetic Diversity Analysis of South and East Asian Duck Populations Using Highly Polymorphic Microsatellite Markers. Asian-Australas. J. Anim. Sci. 2016, 29, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Y.; Wang, W.; Pang, Y.Z.; Chen, W.G.; Yang, Y.B. Microsatellite marker analysis of genetic diversity in the two populations of Yellow River carps. Heilongjiang Anim. Sci. Vet. Med. 2016, 12, 22–26. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Locus Name | Accession Number | Primer Sequence (5′ - 3′) | Fluorescent Type | N | NA | Size (bp) | Repeat Motif | Ta (°C) | Ho | HE | PIC | PHWE | Null Alleles |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CLC01 | OM868049 | F:CCCATTCTGTCTTCAAATCTAAGC R:GCCAATGCTCTACTATGCTTGTC | 5′6-FAM | 240 | 10 | 108-144 | (AGAT)14 | 54 | 0.688 | 0.689 | 0.650 | 0.2951 | 0.0016 |
| CLC02 | OM868050 | F:GGAAGAGAAACCAGTGTGTAGCA R:CATGAGGTGCTGAAGTCCACTAT | 5′6-FAM | 240 | 7 | 106-124 | (AAG)16 | 56 | 0.596 | 0.582 | 0.545 | 0.5313 | −0.0138 |
| CLC03 | OM868051 | F:GGGTGAGAAGGATAGGAAAGAAA R:TTTAACAACCCATGAATTAAAAACC | 5′6-FAM | 240 | 12 | 88-140 | (AGAT)17 | 54 | 0.883 | 0.861 | 0.844 | 0.0069 | −0.0164 |
| CLC04 | OM868052 | F:TTTAATGGGAAAGTTTAATGGATCA R:CTGTACTGCTTCCACCTGATTG | 5′6-FAM | 240 | 9 | 103-135 | (TAGA)14 | 55 | 0.817 | 0.842 | 0.822 | 0.6146 | 0.0157 |
| CLC05 | OM868053 | F:AATAAACAAGGAAAATAATTGCTGG R:ATTGATGGCTAATTTTGCTGGTA | 5′6-FAM | 240 | 9 | 100-127 | (TAG)18 | 54 | 0.813 | 0.840 | 0.817 | 0.0011 * | 0.0158 |
| CLC06 | OM868054 | F:ATGTTGGTATATGAAGCCTGGAT R:TGACAGTATTTCCTCCATCATCA | 5′6-FAM | 240 | 11 | 106-136 | (AAG)15 | 54 | 0.854 | 0.834 | 0.811 | 0.2034 | −0.0133 |
| CLC07 | OM868055 | F:TCTGAAGTTGACCGTATGCTTTT R:CTTTCTTTCTCCATTGTCACCAC | 5′6-FAM | 240 | 12 | 153-193 | (AGAT)15 | 56 | 0.879 | 0.859 | 0.840 | 0.1704 | −0.0128 |
| CLC08 | OM868056 | F:CGTAGTGCTATTTGGGGTATTGA R:TTGTCCTACAATATTCCATGTTTGTT | 5′6-FAM | 240 | 8 | 156-188 | (ATAG)15 | 55 | 0.829 | 0.810 | 0.782 | 0.1714 | −0.0112 |
| CLC09 | OM868057 | F:GAACCACTTGCAGAATAAACACC R:TCATGATCAAAGTTCCTGACTTAAA | 5′6-FAM | 240 | 8 | 150-182 | (TAGA)15 | 56 | 0.825 | 0.820 | 0.794 | 0.5680 | −0.004 |
| CLC10 | OM868058 | F:CGCTCTGAGAAAGAAAAACTCAT R:GAACTTTAGATTCTCGGAAGGAAA | 5′6-FAM | 240 | 8 | 159-191 | (TAGA)15 | 55 | 0.779 | 0.835 | 0.814 | 0.5609 | 0.0312 |
| CLC11 | OM868059 | F:TGAGCAAACATGATTTGAATTTG R:TGTTCAAACATTTGCATCATTTC | 5′-HEX | 240 | 9 | 168-207 | (AAG)17 | 54 | 0.725 | 0.741 | 0.707 | 0.3128 | 0.0088 |
| CLC12 | OM868060 | F:GCAATCCTCCAAAGATATTCCTC R:CATGTTTTTGAGGATGAGACTTTTT | 5′-HEX | 240 | 17 | 194-258 | (TAGA)21 | 54 | 0.863 | 0.900 | 0.890 | 0.0003 ** | 0.022 |
| CLC13 | OM868061 | F:TCCCAGGTTATGAGTTATGGTGT R:GCTTTACTTCTCTAAAACAGCTCTGA | 5′-HEX | 240 | 9 | 221-245 | (AAG)17 | 55 | 0.813 | 0.803 | 0.779 | 0.0217 | −0.0098 |
| CLC14 | OM868062 | F:TTACTGGGGATAGATAGATGGCT R:TCTTTGTCTGTCTATGTATCTGCCT | 5′-HEX | 240 | 17 | 138-238 | (TAGA)22 | 54 | 0.804 | 0.836 | 0.817 | 0.4236 | 0.0207 |
| CLC15 | OM868063 | F:TGAGGTTGAGGTATAAAGGGAAAC R:CACTATCTTTTTCCATCCTTTCCA | 5′-HEX | 240 | 10 | 206-246 | (TAGA)16 | 53 | 0.867 | 0.802 | 0.781 | 0.0053 | −0.0474 |
| CLC16 | OM868064 | F:GGAGAAACGTCTCAATTCACTGT R:CGTGCACATAGTTTATGCTGAGA | 5′-HEX | 240 | 13 | 202-259 | (AGAT)15 | 56 | 0.825 | 0.838 | 0.820 | 0.2843 | 0.0067 |
| CLC17 | OM868065 | F:AAATACCGTATACACATGGGGGT R:CACAAGGACAAAAATGGTGTTTT | 5′-HEX | 240 | 8 | 218-246 | (TAGA)15 | 56 | 0.800 | 0.833 | 0.808 | 0.0296 | 0.0199 |
| CLC18 | OM868066 | F:TTTACAAGCCAAGCTGAAAGAAT R:CCACTCTCATAATGTCTCTGTTTCA | 5′-HEX | 240 | 12 | 197-243 | (ATAG)20 | 56 | 0.771 | 0.833 | 0.816 | 0.0044 | 0.044 |
| CLC19 | OM868067 | F:GCCTGAAAAATGTGTTCCTTTTA R:TGCCTCTTATTCAAAGGCTTTAC | 5′-HEX | 240 | 9 | 223-256 | (AGAT)15 | 54 | 0.846 | 0.800 | 0.773 | 0.0296 | −0.0273 |
| CLC20 | OM868068 | F:AAAATTGGTTTGAAGATGAAGCA R:TATGGCAATGTGGTGCAAAT | 5′-HEX | 240 | 12 | 216-256 | (TAGA)18 | 54 | 0.867 | 0.850 | 0.833 | 0.2817 | −0.0132 |
| Population | N | NA | NE | I | HO | HE | PIC |
|---|---|---|---|---|---|---|---|
| MS | 60 | 9.750 ± 0.542 | 5.771 ± 0.384 | 1.897 ± 0.067 | 0.791 ± 0.019 | 0.813 ± 0.020 | 0.784 ± 0.021 |
| WH | 60 | 8.700 ± 0.449 | 4.624 ± 0.304 | 1.695 ± 0.066 | 0.828 ± 0.021 | 0.768 ± 0.019 | 0.730 ± 0.022 |
| SS | 60 | 7.850 ± 0.519 | 4.065 ± 0.259 | 1.588 ± 0.055 | 0.806 ± 0.022 | 0.740 ± 0.018 | 0.701 ± 0.018 |
| YB | 60 | 9.750 ± 0.542 | 5.683 ± 0.277 | 1.915 ± 0.050 | 0.803 ± 0.019 | 0.821 ± 0.012 | 0.792 ± 0.013 |
| Population | MS | SS | WH | YB |
|---|---|---|---|---|
| MS | 0.000 | |||
| SS | 0.028 | 0.000 | ||
| WH | 0.019 | 0.039 | 0.000 | |
| YB | 0.009 | 0.031 | 0.023 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Mou, C.; Zhou, J.; Ye, H.; Wei, Z.; Ke, H.; Huang, Z.; Duan, Y.; Zhao, Z.; Zhao, H.; et al. Genetic Diversity of Chinese Longsnout Catfish (Leiocassis longirostris) in Four Farmed Populations Based on 20 New Microsatellite DNA Markers. Diversity 2022, 14, 654. https://0-doi-org.brum.beds.ac.uk/10.3390/d14080654
Zhang L, Mou C, Zhou J, Ye H, Wei Z, Ke H, Huang Z, Duan Y, Zhao Z, Zhao H, et al. Genetic Diversity of Chinese Longsnout Catfish (Leiocassis longirostris) in Four Farmed Populations Based on 20 New Microsatellite DNA Markers. Diversity. 2022; 14(8):654. https://0-doi-org.brum.beds.ac.uk/10.3390/d14080654
Chicago/Turabian StyleZhang, Lu, Chenyan Mou, Jian Zhou, Hua Ye, Zhen Wei, Hongyu Ke, Zhipeng Huang, Yuanliang Duan, Zhongmeng Zhao, Han Zhao, and et al. 2022. "Genetic Diversity of Chinese Longsnout Catfish (Leiocassis longirostris) in Four Farmed Populations Based on 20 New Microsatellite DNA Markers" Diversity 14, no. 8: 654. https://0-doi-org.brum.beds.ac.uk/10.3390/d14080654