
Small Vessel Impact on the Whistle Parameters of Two Ecotypes of Common Bottlenose Dolphin (Tursiops truncatus) in La Paz Bay, Mexico

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erbe, C.; Marley, S.A.; Schoeman, R.P.; Smith, J.N.; Trigg, L.E.; Embling, C.B. The effects of ship noise on marine mammals—A review. Front. Mar. Sci. 2019, 6, 606. [Google Scholar] [CrossRef]
- Van der Graaf, A.; Ainslie, M.; André, M.; Brensing, K.; Dalen, J.; Dekeling, R.; Robinson, S.; Tasker, M.; Thomsen, F.; Werner, S. European Marine Strategy Framework Directive-Good Environmental Status (msfd ges): Report of the Technical Subgroup on Underwater Noise and Other Forms of Energy; TSG Noise & Milieu Ltd: Brussels, Belgium, 2012. [Google Scholar]
- Hildebrand, J.A. Anthropogenic and natural sources of ambient noise in the ocean. Mar. Ecol. Prog. Ser. 2009, 395, 5–20. [Google Scholar] [CrossRef]
- Andrew, R.K.; Howe, B.M.; Mercer, J.A.; Dzieciuch, M.A. Ocean ambient sound: Comparing the 1960s with the 1990s for a receiver off the California coast. Acoust. Res. Lett. Online 2002, 3, 65–70. [Google Scholar] [CrossRef]
- Hermannsen, L.; Mikkelsen, L.; Tougaard, J.; Beedholm, K.; Johnson, M.; Madsen, P.T. Recreational vessels without Automatic Identification System (AIS) dominate anthropogenic noise contributions to a shallow water soundscape. Sci. Rep. 2019, 9, 15477. [Google Scholar] [CrossRef] [PubMed]
- Erbe, C. Underwater noise of whale-watching boats and potential effects on killer whales (Orcinus orca), based on an acoustic impact model. Mar. Mammal Sci. 2002, 18, 394–418. [Google Scholar] [CrossRef]
- Gordon, J.; Tyack, P.L. Sound and cetaceans. In Marine Mammals; Springer: Berlin/Heidelberg, Germany, 2002; pp. 139–196. [Google Scholar]
- Janik, V.M.; Sayigh, L.S. Communication in bottlenose dolphins: 50 years of signature whistle research. J. Comp. Physiol. A 2013, 199, 479–489. [Google Scholar] [CrossRef]
- King, S.L.; Janik, V.M. Bottlenose dolphins can use learned vocal labels to address each other. Proc. Natl. Acad. Sci. USA 2013, 110, 13216–13221. [Google Scholar] [CrossRef]
- Heiler, J.; Elwen, S.H.; Kriesell, H.; Gridley, T. Changes in bottlenose dolphin whistle parameters related to vessel presence, surface behaviour and group composition. Anim. Behav. 2016, 117, 167–177. [Google Scholar] [CrossRef]
- May-Collado, L.J.; Wartzok, D. A comparison of bottlenose dolphin whistles in the Atlantic Ocean: Factors promoting whistle variation. J. Mammal. 2008, 89, 1229–1240. [Google Scholar] [CrossRef]
- Richardson, W.J.; Greene, C.R., Jr.; Malme, C.I.; Thomson, D.H. Marine Mammals and Noise; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Oswald, J.N.; Rankin, S.; Barlow, J. To whistle or not to whistle? Geographic variation in the whistling behavior of small odontocetes. Aquat. Mamm. 2008, 34, 288–302. [Google Scholar] [CrossRef]
- Rako, N.; Fortuna, C.M.; Holcer, D.; Mackelworth, P.; Nimak-Wood, M.; Pleslić, G.; Sebastianutto, L.; Vilibić, I.; Wiemann, A.; Picciulin, M. Leisure boating noise as a trigger for the displacement of the bottlenose dolphins of the Cres–Lošinj archipelago (northern Adriatic Sea, Croatia). Mar. Pollut. Bull. 2013, 68, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Fouda, L.; Wingfield, J.E.; Fandel, A.D.; Garrod, A.; Hodge, K.B.; Rice, A.N.; Bailey, H. Dolphins simplify their vocal calls in response to increased ambient noise. Biol. Lett. 2018, 14, 20180484. [Google Scholar] [CrossRef] [PubMed]
- Kragh, I.M.; McHugh, K.; Wells, R.S.; Sayigh, L.S.; Janik, V.M.; Tyack, P.L.; Jensen, F.H. Signal-specific amplitude adjustment to noise in common bottlenose dolphins (Tursiops truncatus). J. Exp. Biol. 2019, 222, jeb216606. [Google Scholar] [CrossRef] [PubMed]
- Perez-Ortega, B.; Daw, R.; Paradee, B.; Gimbrere, E.; May-Collado, L.J. Dolphin-Watching Boats Affect Whistle Frequency Modulation in Bottlenose Dolphins. Front. Mar. Sci. 2021, 8, 102. [Google Scholar] [CrossRef]
- Williams, R.; Lusseau, D.; Hammond, P.S. Estimating relative energetic costs of human disturbance to killer whales (Orcinus orca). Biol. Conserv. 2006, 133, 301–311. [Google Scholar] [CrossRef]
- Aguilar Soto, N.; Johnson, M.; Madsen, P.T.; Tyack, P.L.; Bocconcelli, A.; Fabrizio Borsani, J. Does intense ship noise disrupt foraging in deep-diving Cuvier’s beaked whales (Ziphius cavirostris)? Mar. Mammal Sci. 2006, 22, 690–699. [Google Scholar] [CrossRef]
- Wisniewska, D.M.; Johnson, M.; Teilmann, J.; Siebert, U.; Galatius, A.; Dietz, R.; Madsen, P.T. High rates of vessel noise disrupt foraging in wild harbour porpoises (Phocoena phocoena). Proc. R. Soc. B Biol. Sci. 2018, 285, 20172314. [Google Scholar] [CrossRef]
- Wright, A.J.; Soto, N.A.; Baldwin, A.L.; Bateson, M.; Beale, C.M.; Clark, C.; Deak, T.; Edwards, E.F.; Fernández, A.; Godinho, A. Do marine mammals experience stress related to anthropogenic noise? Int. J. Comp. Psychol. 2007, 20, 274–316. [Google Scholar]
- Segura, I.; Rocha-Olivares, A.; Flores-Ramírez, S.; Rojas-Bracho, L. Conservation implications of the genetic and ecological distinction of Tursiops truncatus ecotypes in the Gulf of California. Biol. Conserv. 2006, 133, 336–346. [Google Scholar] [CrossRef]
- Bearzi, M.; Saylan, C.A.; Hwang, A. Ecology and comparison of coastal and offshore bottlenose dolphins (Tursiops truncatus) in California. Mar. Freshw. Res. 2009, 60, 584–593. [Google Scholar] [CrossRef]
- Viloria-Gómora, L.; Medrano-González, L. Population ecological traits of Tursiops truncatus putative morphotypes in the transitional region of the Mexican Pacific Ocean. Therya 2015, 6, 351–369. [Google Scholar] [CrossRef] [Green Version]
- Gao, A.; Zhou, K.; Wang, Y. Geographical variation in morphology of bottlenosed dolphins (Tursiops sp.) in Chinese waters. Aquat. Mamm. 1995, 21, 121. [Google Scholar]
- Díaz-Gamboa, R. Diferenciación Entre Tursiones Tursiops Truncatus Costeros y Oceánicos en el Golfo de California por Medio de Análisis de Isótopos Estables de Carbono Y Nitrógeno. Master’s Thesis, Centro Interdisciplinario de Ciencias Marinas, La Paz, Mexico, 2003. [Google Scholar]
- Lowther-Thieleking, J.L.; Archer, F.I.; Lang, A.R.; Weller, D.W. Genetic differentiation among coastal and offshore common bottlenose dolphins, Tursiops truncatus, in the eastern North Pacific Ocean. Mar. Mammal Sci. 2015, 31, 1–20. [Google Scholar] [CrossRef]
- Peters, C.H. Context-Specific Signal Plasticity of Two Common Bottlenose Dolphin Ecotypes (Tursiops truncatus) in Far North Waters. Ph.D. Thesis, Massey University, Albany, New Zealand, 2018. [Google Scholar]
- Salinas Zacarías, M.A. Ecología de los Tursiones, Tursiops truncatus, en la Bahía de La Paz, BCS; Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas: La Paz, Mexico, 2005. [Google Scholar]
- Wells, R.; Natoli, A.; Braulik, G. Tursiops truncatus (Errata Version Published in 2019). The IUCN Red List of Threatened Species: E. T22563A156932432. 2019. Available online: https://www.iucnredlist.org/species/22563/156932432 (accessed on 10 August 2022).
- Bittencourt, L.; Barbosa, M.; Santos-Neto, E.B.; Bisi, T.L.; Lailson-Brito, J.; Azevedo, A.F. Whistles of Atlantic spotted dolphin from a coastal area in the southwestern Atlantic Ocean. J. Acoust. Soc. Am. 2020, 148, EL420–EL426. [Google Scholar] [CrossRef]
- Kelly, C.; Glegg, G.; Speedie, C. Management of marine wildlife disturbance. Ocean. Coast. Manag. 2004, 47, 1–19. [Google Scholar] [CrossRef]
- Nowacek, S.M.; Wells, R.S.; Solow, A.R. Short-term effects of boat traffic on bottlenose dolphins, Tursiops truncatus, in Sarasota Bay, Florida. Mar. Mammal Sci. 2001, 17, 673–688. [Google Scholar] [CrossRef]
- Marley, S.A.; Kent, C.P.S.; Erbe, C.; Parnum, I.M. Effects of vessel traffic and underwater noise on the movement, behaviour and vocalisations of bottlenose dolphins in an urbanised estuary. Sci. Rep. 2017, 7, 13437. [Google Scholar] [CrossRef]
- Acevedo, A. Interactions between boats and bottlenose dolphins, Tursiops truneatus, in the entrance to Ensenada De La Paz, Mexico. Aquat. Mamm. 1991, 17, 120–124. [Google Scholar]
- Gregory, P.R.; Rowden, A.A. Behaviour patterns of bottlenose dolphins (Tursiops truncatus) relative to tidal state, time-of-day, and boat traffic in Cardigan Bay, West Wales. Aquat. Mamm. 2001, 27, 105–113. [Google Scholar]
- Mann, J. Behavioral sampling methods for cetaceans: A review and critique. Mar. Mammal Sci. 1999, 15, 102–122. [Google Scholar] [CrossRef]
- Lachlan, R.F. Luscinia: A bioacoustics Analysis Computer Program. Version 1.0 [Computer Program]. Available online: www.lusciniasound.org (accessed on 30 June 2007).
- Quick, N.J.; Janik, V.M. Whistle rates of wild bottlenose dolphins (Tursiops truncatus): Influences of group size and behavior. J. Comp. Psychol. 2008, 122, 305. [Google Scholar] [CrossRef] [PubMed]
- Buckstaff, K.C. Effects of watercraft noise on the acoustic behavior of bottlenose dolphins, Tursiops truncatus, in Sarasota Bay, Florida. Mar. Mammal Sci. 2004, 20, 709–725. [Google Scholar] [CrossRef]
- Merchant, N.D.; Fristrup, K.M.; Johnson, M.P.; Tyack, P.L.; Witt, M.J.; Blondel, P.; Parks, S.E. Measuring acoustic habitats. Methods Ecol. Evol. 2015, 6, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Barlett, M.L.; Wilson, G.R. Characteristics of small boat signatures. J. Acoust. Soc. Am. 2002, 112, 2221. [Google Scholar] [CrossRef]
- Amoser, S.; Wysocki, L.E.; Ladich, F. Noise emission during the first powerboat race in an Alpine lake and potential impact on fish communities. J. Acoust. Soc. Am. 2004, 116, 3789–3797. [Google Scholar] [CrossRef]
- Picciulin, M.; Sebastianutto, L.; Codarin, A.; Farina, A.; Ferrero, E.A. In situ behavioural responses to boat noise exposure of Gobius cruentatus (Gmelin, 1789; fam. Gobiidae) and Chromis chromis (Linnaeus, 1758; fam. Pomacentridae) living in a Marine Protected Area. J. Exp. Mar. Biol. Ecol. 2010, 386, 125–132. [Google Scholar] [CrossRef]
- Mensinger, A.F.; Putland, R.L.; Radford, C.A. The effect of motorboat sound on Australian snapper Pagrus auratus inside and outside a marine reserve. Ecol. Evol. 2018, 8, 6438–6448. [Google Scholar] [CrossRef]
- Herzing, D.L. Vocalizations and associated underwater behavior of free-ranging Atlantic spotted dolphins, Stenella frontalis and bottlenose dolphins, Tursiops truncatus. Aquat. Mamm. 1996, 22, 61–80. [Google Scholar]
- Gospić, N.R.; Picciulin, M. Changes in whistle structure of resident bottlenose dolphins in relation to underwater noise and boat traffic. Mar. Pollut. Bull. 2016, 105, 193–198. [Google Scholar] [CrossRef]
- Van Ginkel, C.; Becker, D.M.; Gowans, S.; Simard, P. Whistling in a noisy ocean: Bottlenose dolphins adjust whistle frequencies in response to real-time ambient noise levels. Bioacoustics 2018, 27, 391–405. [Google Scholar] [CrossRef]
- Kinsler, L.E.; Frey, A.R.; Coppens, A.B.; Sanders, J.V. Fundamentals of Acoustics; John Wiley & Sons: Hoboken, NJ, USA, 2000. [Google Scholar]
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Dinno, A. dunn. test: Dunn’s test of multiple comparisons using rank sums. R package version 2017, 1, 1. [Google Scholar]
- La Manna, G.; Manghi, M.; Pavan, G.; Lo Mascolo, F.; Sarà, G. Behavioural strategy of common bottlenose dolphins (Tursiops truncatus) in response to different kinds of boats in the waters of Lampedusa Island (Italy). Aquat. Conserv. Mar. Freshw. Ecosyst. 2013, 23, 745–757. [Google Scholar] [CrossRef]
- Morisaka, T.; Shinohara, M.; Nakahara, F.; Akamatsu, T. Effects of ambient noise on the whistles of Indo-Pacific bottlenose dolphin populations. J. Mammal. 2005, 86, 541–546. [Google Scholar] [CrossRef]
- Scarpaci, C.; Bigger, S.W.; Corkeron, P.J.; Nugegoda, D. Bottlenose dolphins (Tursiops truncatus) increase whistling in the presence ofswim-with-dolphin’tour operations. J. Cetacean Res. Manag. 2000, 2, 183–185. [Google Scholar]
- Urick, R.J. Principles of Underwater Sound, 2nd ed.; McGraw-Hill Book Company: New York, NY, USA, 1975. [Google Scholar]
- Leon-Lopez, B.; Romero-Vivas, E.; Viloria-Gomora, L. Reduction of roadway noise in a coastal city underwater soundscape during COVID-19 confinement. J. Acoust. Soc. Am. 2021, 149, 652–659. [Google Scholar] [CrossRef] [PubMed]
- Richardson, W.J.; Würsig, B. Influences of man-made noise and other human actions on cetacean behaviour. Mar. Freshw. Behav. Phy. 1997, 29, 183–209. [Google Scholar] [CrossRef]
- Lusseau, D. The hidden cost of tourism: Detecting long-term effects of tourism using behavioral information. Ecol. Soc. 2004, 9. [Google Scholar] [CrossRef]
- Sini, M.; Canning, S.J.; Stockin, K.; Pierce, G.J. Bottlenose dolphins around Aberdeen harbour, north-east Scotland: A short study of habitat utilization and the potential effects of boat traffic. J. Mar. Biol. Assoc. United Kingd. 2005, 85, 1547–1554. [Google Scholar] [CrossRef]
- Buckingham, C.A.; Lefebvre, L.W.; Schaefer, J.M.; Kochman, H.I. Manatee response to boating activity in a thermal refuge. Wildl. Soc. Bull. 1999, 27, 514–522. [Google Scholar]
- Holt, M.M.; Noren, D.P.; Dunkin, R.C.; Williams, T.M. Vocal performance affects metabolic rate in dolphins: Implications for animals communicating in noisy environments. J. Exp. Biol. 2015, 218, 1647–1654. [Google Scholar] [CrossRef]
- Wartzok, D.; Poppper, A.; Gordon, J.; Merrill, J. Factors affecting the responses of marine mammals to acoustic disturbance. Mar. Technol. Soc. J. 2004, 37, 6–15. [Google Scholar] [CrossRef]
- Jensen, F.H.; Bejder, L.; Wahlberg, M.; Soto, N.A.; Johnson, M.; Madsen, P.T. Vessel noise effects on delphinid communication. Mar. Ecol. Prog. Ser. 2009, 395, 161–175. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
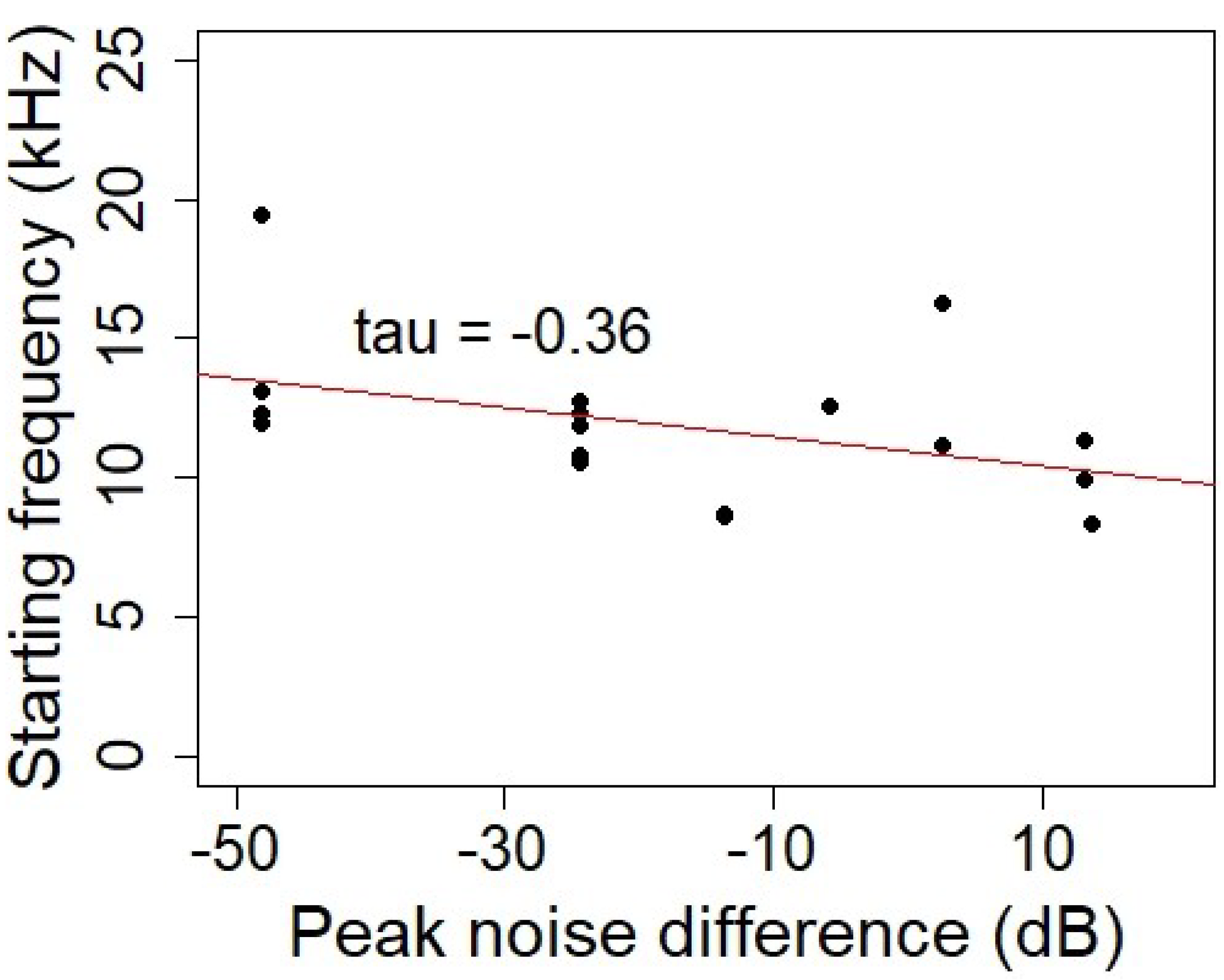{kind=link}
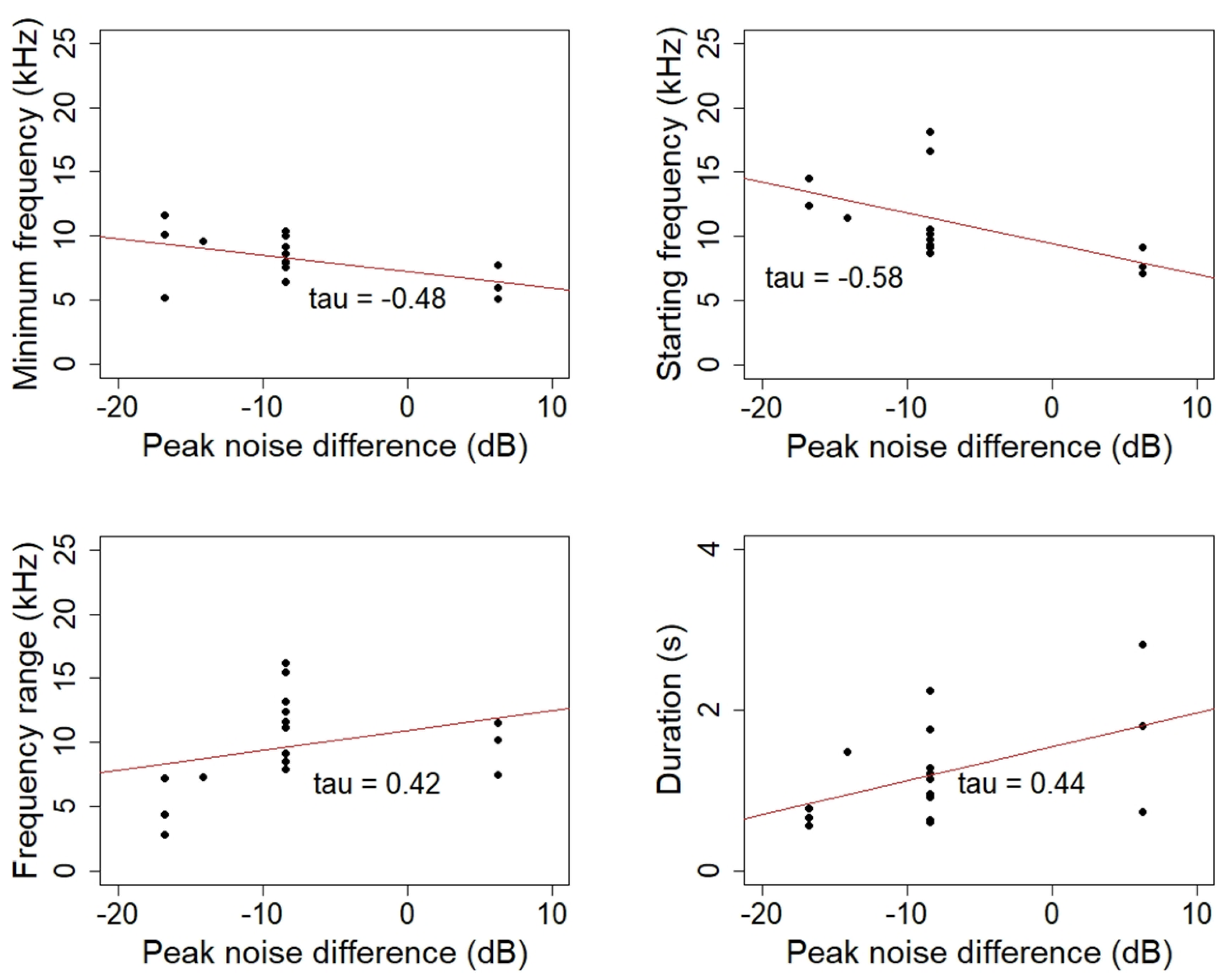{kind=link}
| Vessel Type | Definition | Length (m) |
|---|---|---|
| Fishing panga | Panga * vessel used for fishing activities. | 5–10 |
| Touristic panga | Panga vessel used for touristic activities. | 5–10 |
| Passenger panga | Panga vessel used for carrying passengers from the shore to other vessels or vice versa. | 5–10 |
| Ferry | Large-sized vessel used for carrying people and goods as a regular service from one port to another. | 150–200 |
| Yacht | Vessel used for recreational activities. | 10–30 |
| Sailing boat | Any vessel propelled entirely or partly by sails. | 5–30 |
| Cargo | Merchant vessel carrying goods and materials from one port to another. | 150–200 |
| Jet ski | Personal watercraft used for recreational activities | 2–4 |
| Absence of Vessels | Presence of a Single Small Vessel | Total | |
|---|---|---|---|
| Oceanic ecotype | |||
| N° groups recorded | 5 | 4 | 5 |
| N° recordings | 17 | 5 | 22 |
| Recording duration (min) | 167 | 55 | 222 |
| N° whistles | 190 | 105 | 295 |
| Coastal ecotype | |||
| N° groups recorded | 13 | 10 | 16 |
| N° recordings | 35 | 11 | 46 |
| Recording duration (min) | 252 | 83 | 335 |
| N° whistles | 185 | 78 | 263 |
| Oceanic Ecotype | Absence of Vessels (n = 190) | Presence of a Single Small Vessel (n = 105) | ||||
| Mean | sd | Median | Mean | sd | Median | |
| Duration (s) | 1.13 | 0.60 | 1.08 | 1.07 | 0.56 | 0.92 |
| Starting frequency (kHz) | 12.10 | 5.76 | 9.95 | 10.75 | 4.40 | 9.46 |
| Ending frequency (kHz) | 8.97 | 3.72 | 7.82 | 9.31 | 4.55 | 8.29 |
| Minimum frequency (kHz) | 7.32 | 2.14 | 7.01 | 7.07 | 1.96 | 6.71 |
| Maximum frequency (kHz) * | 18.57 | 4.37 | 18.53 | 17.01 | 3.52 | 16.96 |
| Frequency range (kHz) * | 11.26 | 4.33 | 11.12 | 9.93 | 3.07 | 9.79 |
| Peak frequency (kHz) | 11.76 | 3.58 | 11.13 | 11.11 | 2.69 | 10.80 |
| Whistle rate | 0.02 | 0.01 | 0.02 | 0.04 | 0.04 | 0.03 |
| Coastal Ecotype | Absence of Vessels (n = 185) | Presence of a Single Small Vessel (n = 78) | ||||
| Mean | sd | Median | Mean | sd | Median | |
| Duration (s) | 1.08 | 0.70 | 0.99 | 1.23 | 0.80 | 1.12 |
| Starting frequency (kHz) | 10.69 | 4.60 | 10.18 | 11.62 | 3.85 | 11.00 |
| Ending frequency (kHz) * | 9.11 | 3.31 | 8.66 | 10.02 | 2.97 | 9.88 |
| Minimum frequency (kHz) * | 6.95 | 2.03 | 6.65 | 7.89 | 2.00 | 7.95 |
| Maximum frequency (kHz) | 15.28 | 3.90 | 14.95 | 16.07 | 3.32 | 15.46 |
| Frequency range (kHz) | 8.33 | 3.82 | 7.86 | 8.17 | 3.06 | 8.17 |
| Peak frequency (kHz) * | 10.00 | 2.75 | 9.68 | 10.82 | 2.63 | 9.95 |
| Whistle rate | 0.05 | 0.08 | 0.03 | 0.04 | 0.05 | 0.02 |
| Oceanic Ecotype | Before Approach (n = 45) | During Approach (n = 16) | After Approach (n = 44) | ||||||
| Mean | sd | Median | Mean | sd | Median | Mean | sd | Median | |
| Duration (s) | 1.06 | 0.62 | 0.81 | 1.22 | 0.65 | 1.05 | 1.03 | 0.44 | 0.96 |
| Starting frequency (kHz) | 10.83 | 4.51 | 9.74 | 11.24 | 3.19 | 10.39 | 10.49 | 4.72 | 9.05 |
| Ending frequency (kHz) * | 10.80 | 5.73 | 9.10 | 9.81 | 4.28 | 8.61 | 7.60 | 2.17 | 7.00 |
| Minimum frequency (kHz) * | 7.31 | 2.08 | 7.11 | 8.22 | 1.90 | 7.97 | 6.42 | 1.62 | 6.29 |
| Maximum frequency (kHz) | 16.90 | 4.29 | 16.81 | 18.00 | 3.33 | 17.06 | 16.76 | 2.61 | 16.91 |
| Frequency range (kHz) | 9.59 | 3.37 | 8.69 | 9.79 | 3.66 | 9.68 | 10.34 | 2.51 | 10.71 |
| Peak frequency (kHz) | 11.63 | 2.74 | 11.34 | 11.86 | 2.99 | 12.11 | 10.32 | 2.34 | 10.27 |
| Whistle rate | 0.04 | 0.04 | 0.02 | 0.03 | 0.04 | 0.02 | 0.06 | 0.08 | 0.03 |
| Coastal Ecotype | Before Approach (n = 23) | During Approach (n = 17) | After Approach (n = 38) | ||||||
| Mean | sd | Median | Mean | sd | Median | Mean | sd | Median | |
| Duration (s) | 1.07 | 0.75 | 0.91 | 1.51 | 1.04 | 1.26 | 1.20 | 0.69 | 1.17 |
| Starting frequency (kHz) | 11.17 | 4.71 | 10.18 | 11.90 | 2.74 | 11.89 | 11.76 | 3.77 | 10.86 |
| Ending frequency (kHz) | 9.99 | 3.69 | 10.98 | 9.93 | 2.58 | 9.68 | 10.08 | 2.71 | 9.88 |
| Minimum frequency (kHz) | 7.38 | 2.93 | 7.01 | 7.74 | 1.06 | 7.90 | 8.00 | 1.64 | 8.27 |
| Maximum frequency (kHz) | 16.24 | 3.85 | 15.58 | 15.88 | 3.03 | 15.53 | 16.04 | 3.17 | 15.06 |
| Frequency range (kHz) | 8.41 | 3.17 | 7.91 | 8.14 | 3.15 | 8.92 | 8.04 | 3.03 | 8.17 |
| Peak frequency (kHz) | 11.48 | 3.05 | 10.69 | 10.16 | 2.21 | 9.40 | 10.71 | 2.50 | 10.35 |
| Whistle rate | 0.03 | 0.04 | 0.01 | 0.05 | 0.05 | 0.04 | 0.04 | 0.05 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antichi, S.; Jaramillo-Legorreta, A.M.; Urbán R., J.; Martínez-Aguilar, S.; Viloria-Gómora, L. Small Vessel Impact on the Whistle Parameters of Two Ecotypes of Common Bottlenose Dolphin (Tursiops truncatus) in La Paz Bay, Mexico. Diversity 2022, 14, 712. https://0-doi-org.brum.beds.ac.uk/10.3390/d14090712
Antichi S, Jaramillo-Legorreta AM, Urbán R. J, Martínez-Aguilar S, Viloria-Gómora L. Small Vessel Impact on the Whistle Parameters of Two Ecotypes of Common Bottlenose Dolphin (Tursiops truncatus) in La Paz Bay, Mexico. Diversity. 2022; 14(9):712. https://0-doi-org.brum.beds.ac.uk/10.3390/d14090712
Chicago/Turabian StyleAntichi, Simone, Armando M. Jaramillo-Legorreta, Jorge Urbán R., Sergio Martínez-Aguilar, and Lorena Viloria-Gómora. 2022. "Small Vessel Impact on the Whistle Parameters of Two Ecotypes of Common Bottlenose Dolphin (Tursiops truncatus) in La Paz Bay, Mexico" Diversity 14, no. 9: 712. https://0-doi-org.brum.beds.ac.uk/10.3390/d14090712