
Replicate Studies Separated by 40 Years Reveal Changes in the Altitudinal Stratification of Montane Passalid Beetle Species (Passalidae) in Mesoamerica
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Data Collection
2.2. Faunistic Analysis
2.3. Change in Climatic Variable
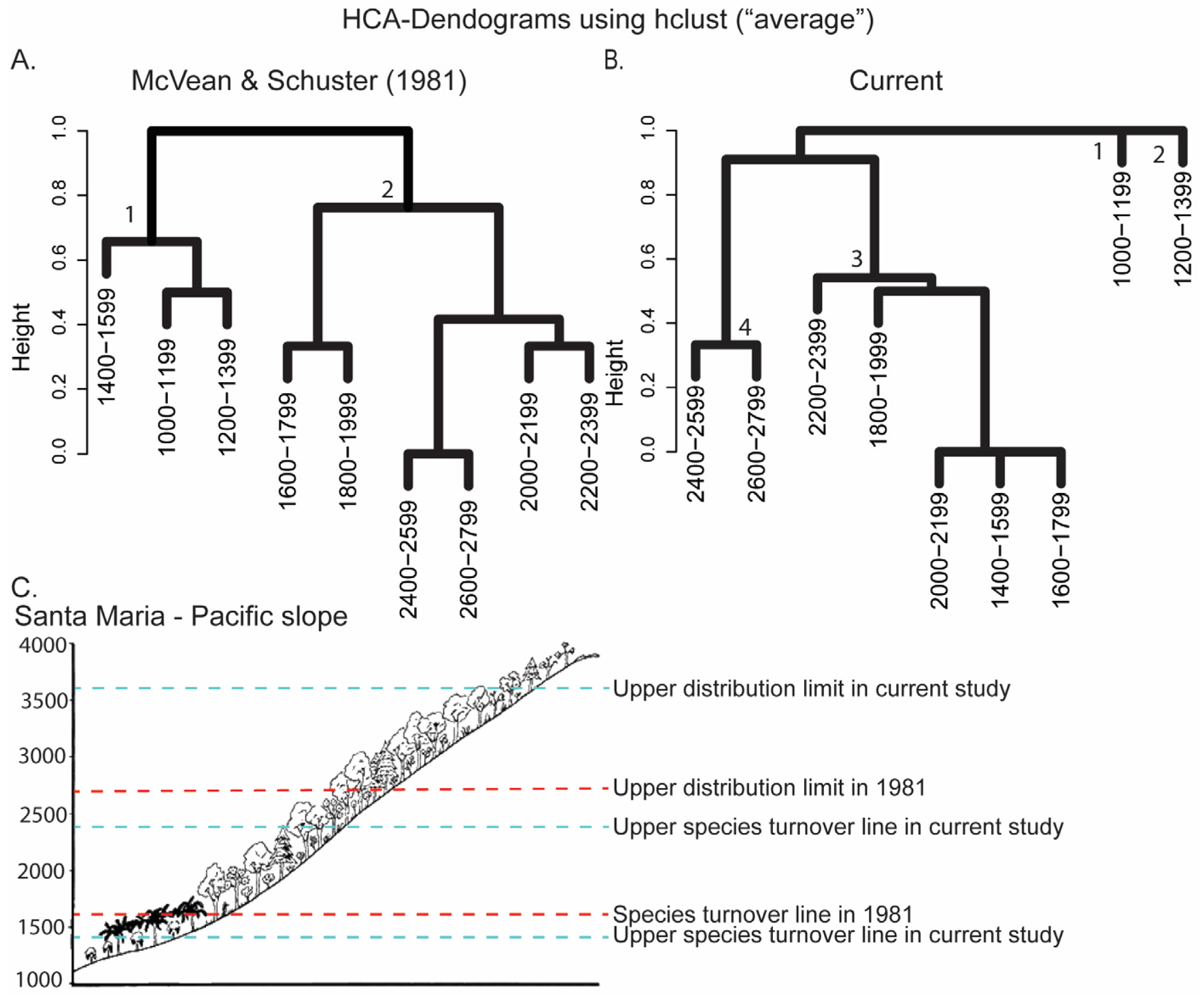
3. Results
3.1. Atitlan Volcano
3.2. Santa Maria Volcano
4. Discussion
- (1)
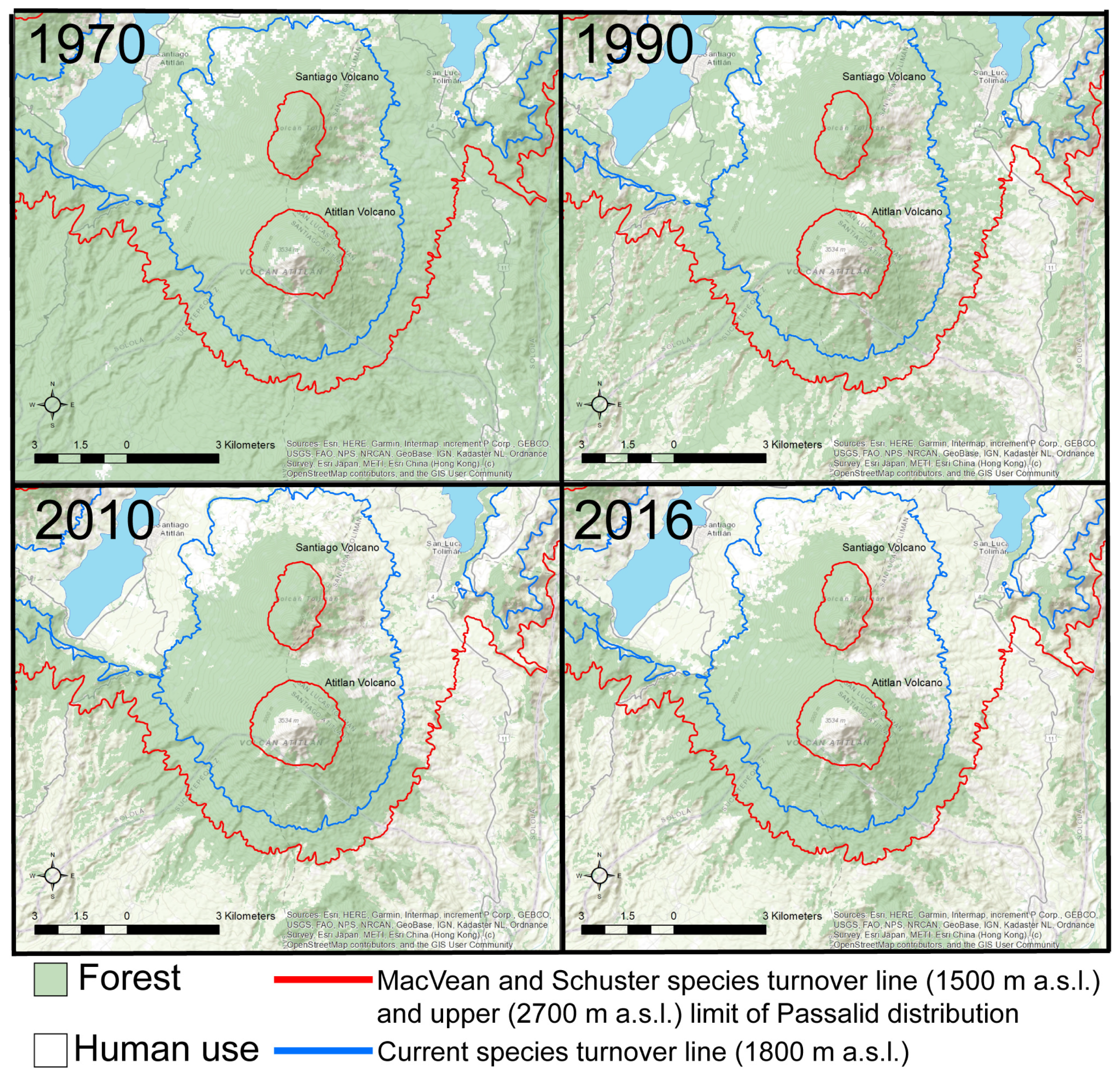
- At the Atitlan volcano, MacVean and Schuster [9] detected a turnover from lowland to montane species at 1500 m a.s.l. (1600 m a.s.l. in our analyses). However, hierarchical cluster analyses of our data indicated that the current species turnover was at 1800 m a.s.l. The 1500 m a.s.l. boundary for species turnover in Passalidae tends to be consistent across altitudinal gradient studies of Passalidae (e.g., Tingo Maria region in Peru [36] and Soconusco México [13], Parque Nacional el Trifinio in El Salvador [32]). In these locations, species turnover is correlated with a change in vegetation from lower montane forest to humid montane forest such as cloud forest. Mesoamerican forest cover declined from 72% in the 1980s to 42% today and humid montane habitats are one of the most threatened environments [14]. Although assessments of the structural composition of the forest are not available, land-use coverage (e.g., humid montane forest) between 1500 and 1800 m a.s.l. on the south side of the Atitlán volcano has not significantly changed in the last 50 years (Figure 4).

- (2)
- For both volcanoes, passalid beetles were present above 2700 m a.s.l. MacVean and Schuster [9] found passalid beetles between 550 and 2650 m a.s.l. (Figure 3C). They reported sampling 25 logs above 2700 m a.s.l. at Atitlan and Santa Maria, but these logs did not contain passalid colonies. In the current study, logs containing passalid beetles were found from 1043 to 3498 m a.s.l. at Santa Maria, and 850 to 2900 m a.s.l. at Atitlan (Table 1). The species found above 2700 m a.s.l. were C. granulifrons, Ps. junctistriatus, and Proculini sp. “incertae sedis”, which are all montane endemic species.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goulson, D. The Insect Apocalypse, and Why It Matters. Curr. Biol. 2019, 29, R967–R971. [Google Scholar] [CrossRef] [PubMed]
- La Sorte, F.A.; Jetz, W. Projected Range Contractions of Montane Biodiversity under Global Warming. Proc. R. Soc. B Biol. Sci. 2010, 277, 3401–3410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurance, W.F.; Useche, D.C.; Shoo, L.P.; Herzog, S.K.; Kessler, M.; Escobar, F.; Brehm, G.; Axmacher, J.C.; Chen, I.-C.; Gámez, L.A.; et al. Global Warming, Elevational Ranges and the Vulnerability of Tropical Biota. Biol. Conserv. 2011, 144, 548–557. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A Globally Coherent Fingerprint of Climate Change Impacts across Natural Systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Nufio, C.R.; McGuire, C.R.; Bowers, M.D.; Guralnick, R.P. Grasshopper Community Response to Climatic Change: Variation Along an Elevational Gradient. PLoS ONE 2010, 5, e12977. [Google Scholar] [CrossRef] [PubMed]
- Molina-Martínez, A.; León-Cortés, J.L.; Regan, H.M.; Lewis, O.T.; Navarrete, D.; Caballero, U.; Luis-Martínez, A. Changes in Butterfly Distributions and Species Assemblages on a Neotropical Mountain Range in Response to Global Warming and Anthropogenic Land Use. Divers. Distrib. 2016, 22, 1085–1098. [Google Scholar] [CrossRef]
- McCain, C.M.; Garfinkel, C.F. Climate Change and Elevational Range Shifts in Insects. Curr. Opin. Insect Sci. 2021, 47, 111–118. [Google Scholar] [CrossRef]
- Halsch, C.A.; Shapiro, A.M.; Fordyce, J.A.; Nice, C.C.; Thorne, J.H.; Waetjen, D.P.; Forister, M.L. Insects and Recent Climate Change. Proc. Natl. Acad. Sci. USA 2021, 118, e2002543117. [Google Scholar] [CrossRef]
- MacVean, C.; Schuster, J.C. Altitudinal Distribution of Passalid Beetles (Coleoptera, Passalidae) and Pleistocene Dispersal on the Volcanic Chain of Northern Central America. Biotropica 1981, 13, 29. [Google Scholar] [CrossRef]
- Reyes-Castillo, P.; Halffter, G. Análisis de La Distribución Geográfica de La Tribu Proculini (Coleoptera: Passalidae). Folia Entomológica Mex. 1978, 39–40, 222–226. [Google Scholar]
- Schuster, J.C.; Cano, E.B.; Cardona, C. Un Método Sencillo Para Priorizar La Conservación de Los Bosques Nubosos de Guatemala, Usando Passalidae (Coleoptera) Como Organismos Indicadores. ACTA Zool. Mex. NS 2000, 80, 197–209. [Google Scholar] [CrossRef]
- Kattan, G.H.; Murcia, C.; Galindo-Cardona, A. An Evaluation of Bess Beetles (Passalidae) and Their Resource Base in a Restored Andean Forest. Trop. Conserv. Sci. 2010, 3, 334–343. [Google Scholar] [CrossRef]
- Chamé Vázquez, E.R.; Reyes-Castillo, P.; Gómez-Gómez, B.; Ibarra-Núñez, G. Distribución de La Familia Passalidae (Coleoptera: Scarabaeoidea) En Un Gradiente Altitudinal En El Soconusco, Chiapas, México. Dugesiana 2018, 25, 115–124. [Google Scholar] [CrossRef]
- Rzedowski, J. Vegetación de México; 1ra. Edicion Digital; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Mexico city, Mexico, 2006. [Google Scholar]
- Arbeláez-Cortés, E.; Nyári, Á.S.; Navarro-Sigüenza, A.G. The Differential Effect of Lowlands on the Phylogeographic Pattern of a Mesoamerican Montane Species (Lepidocolaptes Affinis, Aves: Furnariidae). Mol. Phylogenet. Evol. 2010, 57, 658–668. [Google Scholar] [CrossRef]
- Beza-Beza, C.; Cano, E.; Schuster, J.; Archila, D.; Palmieri, M. Biogeografía Molecular de Escarabajos (Coleoptera: Passalidae) Del Género Ogyges Grupo “Laevissimus” y Sus Implicaciones En La Conservación de Los Bosques Nubosos de Guatemala. Rev. Univ. Val. Guatem. 2011, 23, 18–22. [Google Scholar]
- Schuster, J.C. Biotic Areas and the Distribution of Passalid Beetles (Coleoptera) in Northern Central America: Post-Pleistocene Montane Refuges. In Proceedings of the Tulane Studies in Zoology and Botany Supplementary Publication Number 1; Mérida: Yucatán, Mexico, 1992. [Google Scholar]
- Schuster, J.C. Passalidae (Coleoptera) de Mesoamérica: Diversidad y Biogeografía. In Biodiversidad de Guatemala; Cano, E.B., Ed.; Universidad del Valle de Guatemala: Guatemala City, Guatemala, 2006; Volume 1. [Google Scholar]
- Bunn, C.; Läderach, P.; Ovalle Rivera, O.; Kirschke, D. A Bitter Cup: Climate Change Profile of Global Production of Arabica and Robusta Coffee. Clim. Change 2015, 129, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Laderach, P.; Lundy, M.; Jarvis, A.; Ramirez, J.; Portilla, E.P.; Schepp, K.; Eitzinger, A. Predicted Impact of Climate Change on Coffee Supply Chains. In The Economic, Social and Political Elements of Climate Change; Climate Change Management; Leal Filho, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 703–723. ISBN 978-3-642-14775-3. [Google Scholar]
- Rovito, S.M.; Parra-Olea, G.; Vásquez-Almazán, C.R.; Papenfuss, T.J.; Wake, D.B. Dramatic Declines in Neotropical Salamander Populations Are an Important Part of the Global Amphibian Crisis. Proc. Natl. Acad. Sci. USA 2009, 106, 3231–3236. [Google Scholar] [CrossRef] [Green Version]
- Marshall, S.J. The Geomorphology and Physiographic Provinces of Central America. In Central America: Geology, Resources and Hazards; Bundschuh, J., Alvarado, G., Eds.; Taylor and Frances: London, UK, 2007; Volume 1. [Google Scholar]
- Global Volcanism Program. [Database] Volcanoes of the World (v. 5.0.0; 1 Nov 2022); Smithsonian Institution: Washington, DC, USA, 2022. [Google Scholar]
- Holdridge, L.R. Life Zone Ecology; Tropical Science Center: San Jose, Costa Rica, 1967. [Google Scholar]
- Illescas, V. Plan de Manejo Turístico Del Volcán y Laguna Chikab’al San Martín Scatepequez Quetzaltenango (Strategy Report); Centro de Estudios Ambientales y Biodiversidad: Guatemala City, Guatemala, 2011. [Google Scholar]
- Schuster, J.C.; Cano, E.B. Key to American Genera of Passalidae. Available online: https://unsm-ento.unl.edu/Guide/Scarabaeoidea/Passalidae/Passalidae-Key/Passalidae-key.pdf (accessed on 1 January 2019).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundations Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. Vegan: Community Ecology Package. Ordination Methods, Diversity Analysis and Other Functions for Community and Vegetation Ecologists. R Package Version 2.5. 2019. Available online: https://cran.r-project.org/package=vegan (accessed on 13 December 2021).
- Harris, I.; Osborn, T.J.; Jones, P.; Lister, D. Version 4 of the CRU TS Monthly High-Resolution Gridded Multivariate Climate Dataset. Sci. Data 2020, 7, 109. [Google Scholar] [CrossRef] [Green Version]
- Funk, C.C.; Peterson, P.J.; Landsfeld, M.F.; Pedreros, D.H.; Verdin, J.P.; Rowland, J.D.; Romero, B.E.; Husak, G.J.; Michaelsen, J.C.; Verdin, A.P. A Quasi-Global Precipitation Time Series for Drought Monitoring; Data Series; US Geological Survey: Reston, VA, USA, 2014; p. 12. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio Team: Boston, MA, USA, 2021. [Google Scholar]
- Serrano-Peraza, F.; Serrano Cervantes, L.; Chaves, L.F. Bess Beetle (Coleoptera: Passalidae) Species Composition Across the Altitudinal Gradient of Montecristo National Park, El Salvador. Environ. Entomol. 2022, 51, 557–563. [Google Scholar] [CrossRef]
- McKenna, D.D.; Farrell, B.D. Molecular Phylogenetics and Evolution of Host Plant Use in the Neotropical Rolled Leaf ‘Hispine’ Beetle Genus Cephaloleia (Chevrolat) (Chrysomelidae: Cassidinae). Mol. Phylogenet. Evol. 2005, 37, 117–131. [Google Scholar] [CrossRef]
- McKenna, D.D.; Farrell, B.D. Tropical Forests Are Both Evolutionary Cradles and Museums of Leaf Beetle Diversity. Proc. Natl. Acad. Sci. USA 2006, 103, 10947–10951. [Google Scholar] [CrossRef] [Green Version]
- Royle, J.A.; Nichols, J.D. Estimating Abundance from Repeated Presence-Absence Data of Point Counts. Ecology 2003, 84, 777–790. [Google Scholar] [CrossRef]
- Schuster, J.C. Biogeographical and Ecological Limits of New World Passalidae (Coleoptera). Coleopt. Bull. 1978, 32, 21–28. [Google Scholar]
- Pons, D.A.; Castellanos, E.; Conde, D.; Brincker, J.; Incer, D.; Lopez, A. Escenarios de Aridez Para Guatemala Para Los Años 2030, 2050 y 2070 Utilizando Modelos de Cambio Climático. Rev. Yu’am 2018, 2, 4–16. [Google Scholar]
- Jiménez-Ferbans, L.; Amat García, G.; Reyes-Castillo, P. Diversity and Distribution Patterns of Passalidae (Coleoptera Scarabaeoidea) in the Caribbean Region of Colombia. Trop. Zool. 2010, 23, 147–164. [Google Scholar]
- Universidad del Valle de Guatemala; Instituto Nacional de Bosques (INAB); Consejo Nacional de Areas Protegidas (CONAP); Universidad Rafael Landivar (URL). Dinámica de La Cobertura Forestal de Guatemala Durante Los Años 1991, 1996 y 2001 y Mapa de Cobertura Forestal 2001; UVG/INB/CNAP: Guatemala City, Guatemala, 2006. [Google Scholar]
- Regalado, O.; Villagrán, X.; Pérez, G.; Castellanos, E.; Martínez, G.; Incer, D.; Ramos, V.H.; Molina, O.; Beltetón, C.; Gómez, J.M. Mapa de Cobertura Forestal de Guatemala 2010 y Dinámica de la Cobertura Forestal 2006–2010.; Universidad Rafael Landívar: Guatemala City, Guatemala, 2012. [Google Scholar] [CrossRef]
- Raxworthy, C.J.; Pearson, R.G.; Rabibisoa, N.; Rakotondrazafy, A.M.; Ramanamanjato, J.-B.; Raselimanana, A.P.; Wu, S.; Nussbaum, R.A.; Stone, D.A. Extinction Vulnerability of Tropical Montane Endemism from Warming and Upslope Displacement: A Preliminary Appraisal for the Highest Massif in Madagascar: Madagascan warming and upslope distribution shifts. Glob. Chang. Biol. 2008, 14, 1703–1720. [Google Scholar] [CrossRef] [Green Version]
- Herzog, S.K.; Hamel-Leigue, A.C.; Larsen, T.H.; Mann, D.J.; Soria-Auza, R.W.; Gill, B.D.; Edmonds, W.D.; Spector, S. Elevational Distribution and Conservation Biogeography of Phanaeine Dung Beetles (Coleoptera: Scarabaeinae) in Bolivia. PLoS ONE 2013, 8, e64963. [Google Scholar] [CrossRef] [Green Version]
- Freeman, B.G.; Scholer, M.N.; Ruiz-Gutierrez, V.; Fitzpatrick, J.W. Climate Change Causes Upslope Shifts and Mountaintop Extirpations in a Tropical Bird Community. Proc. Natl. Acad. Sci. USA 2018, 115, 11982–11987. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Atitlan | Santa Maria | |||
|---|---|---|---|---|
| 1981 | Current | 1981 | Current | |
| Paxillus leachi | -- | 850 | -- | -- |
| Passalus punctiger | -- | -- | 800 | 1016 |
| Rhodocanthopus caelatus | 1500 | 850–1668 | 1150 | -- |
| Passalus punctatostriatus | -- | 1100–1300 | 1150–1550 | -- |
| Odontotaenius striatopunctatus | 1400 | 850–1300 | 1150 | 1230–1377 |
| Verres hageni | 1400 | 850 | 1550 | 1043 |
| Spurius bicornis | 1350–1500 | 1100–1668 | 1550 | -- |
| Oileus sargi | -- | 1700–1750 | 1550 | 1423–2274 |
| Arrox agassizi | -- | -- | -- | 2214 |
| Chondrocephalus debilis | 1550–2500 | -- | 1750 | -- |
| Chondrocephalus purulensis | 1950–2300 | 2049–2300 | 1750–2300 | -- |
| Ogyges laevissimus | 2500 | 2300–2350 | 2100–2600 | -- |
| Chondrocephalus granulifrons | 2500–2700 | 1900–2900 | 2200–2650 | 2700–3300 |
| Pseudacanthus subopacus | -- | -- | 2300 | 1814 |
| Vindex c.f. sculptilis | 2500 | -- | -- | 1978 |
| Pseudacanthus junctistriatus | -- | -- | -- | 2395–3124 |
| Proculini sp. | -- | -- | -- | 3498 |
| Total # Species | 9 | 10 | 12 | 10 |
| Volcano | Sampling Period | Cluster # | Species Composition |
| Atitlan | 1982 | Cluster 1 (1200–1599) | R. caelatus, C. debilis, Od. striatopunctatus, S. bicornis, V. hageni |
| Cluster 2 (1600–2399) | C. debilis, C. purulensis | ||
| Cluster 3 (2400–2799) | C. debilis, C. granulifrons, O. laevissimus, V. sp. | ||
| Current | Cluster 1 (1200–1799) | R. caelatus, P. punctatostriatus, O. striatopunctatus, Oi. sargi, S. bicornis | |
| Cluster 2 (1800–1799) | C. granulifrons, C. purulensis, O. laevissimus | ||
| Santa Maria | 1982 | Cluster 1 (1200–1599) | R. caelatus, P. punctatostriatus, Od. striatopunctatus, Oi. sargi, S. bicornis, V. hageni |
| Cluster 2 (1600–2799) | C. debilis, C. granulifrons, C. purulensis, Ps. subopacus, O. laevissimus | ||
| Current | Cluster 1 (1000–1199) | P. punctiger, V. hageni | |
| Cluster 2 (1200–1399) | O. striatopunctatus | ||
| Cluster 1 (1400–2199) | Oi. sargi, Ps. subopacus, V. sp | ||
| Cluster 2 (2200–2799) | Oi. sargi, C. granulifrons, Ps. junctistriatus, A. agassizi |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beza-Beza, C.F.; Rivera, C.; Pons, D.; McKenna, D.; Schuster, J.C. Replicate Studies Separated by 40 Years Reveal Changes in the Altitudinal Stratification of Montane Passalid Beetle Species (Passalidae) in Mesoamerica. Diversity 2023, 15, 315. https://0-doi-org.brum.beds.ac.uk/10.3390/d15030315
Beza-Beza CF, Rivera C, Pons D, McKenna D, Schuster JC. Replicate Studies Separated by 40 Years Reveal Changes in the Altitudinal Stratification of Montane Passalid Beetle Species (Passalidae) in Mesoamerica. Diversity. 2023; 15(3):315. https://0-doi-org.brum.beds.ac.uk/10.3390/d15030315
Chicago/Turabian StyleBeza-Beza, Cristian Fernando, Camilo Rivera, Diego Pons, Duane McKenna, and Jack C. Schuster. 2023. "Replicate Studies Separated by 40 Years Reveal Changes in the Altitudinal Stratification of Montane Passalid Beetle Species (Passalidae) in Mesoamerica" Diversity 15, no. 3: 315. https://0-doi-org.brum.beds.ac.uk/10.3390/d15030315