


Grassland Alterations Do Not Affect Breeding Success, but Can Explain Dietary Shifts of a Generalist Raptor Species


Abstract
:1. Introduction
2. Materials and Methods
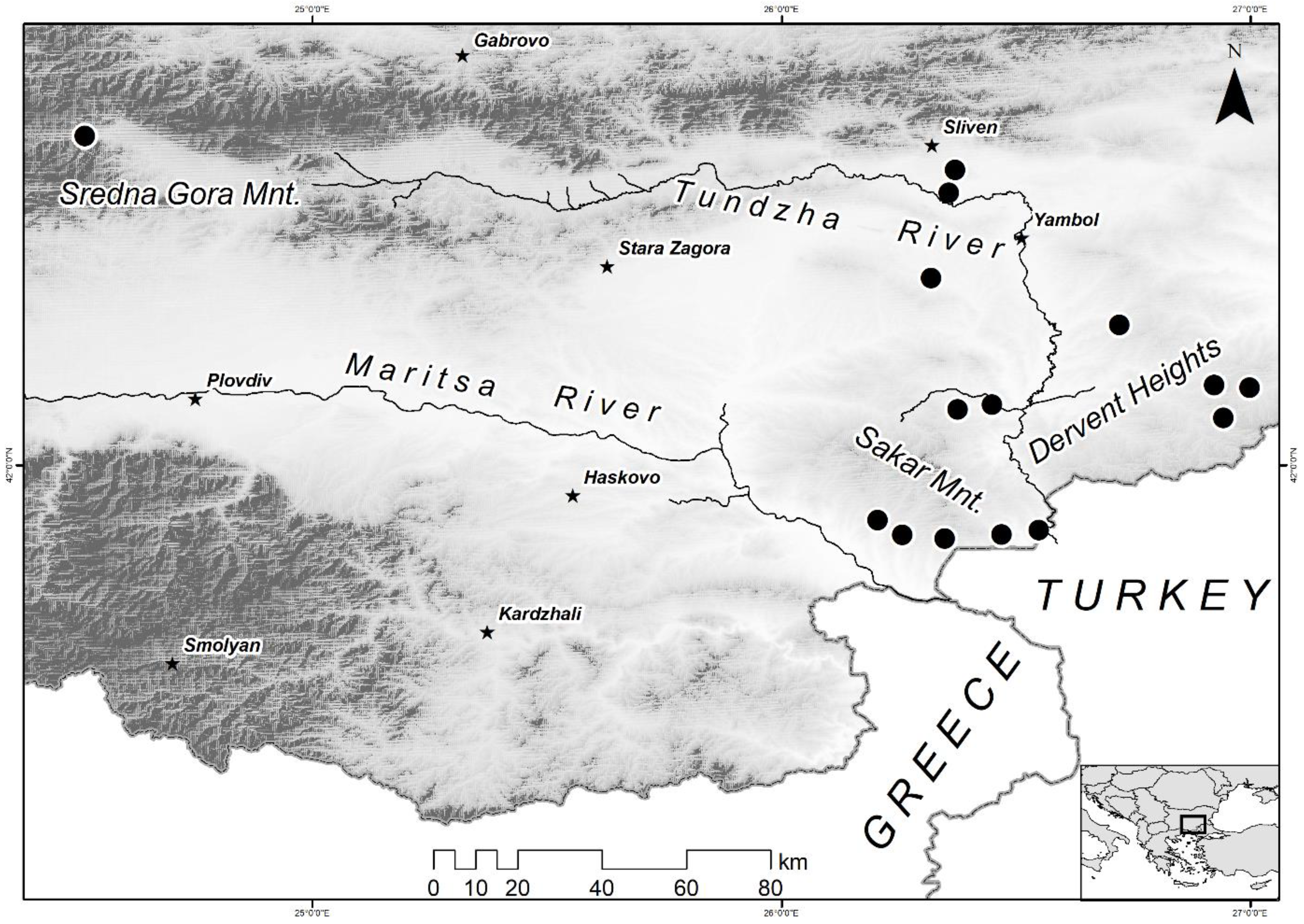
2.1. Habitat Associations
2.2. Dietary Data Analysis
2.3. Statistical Procedures
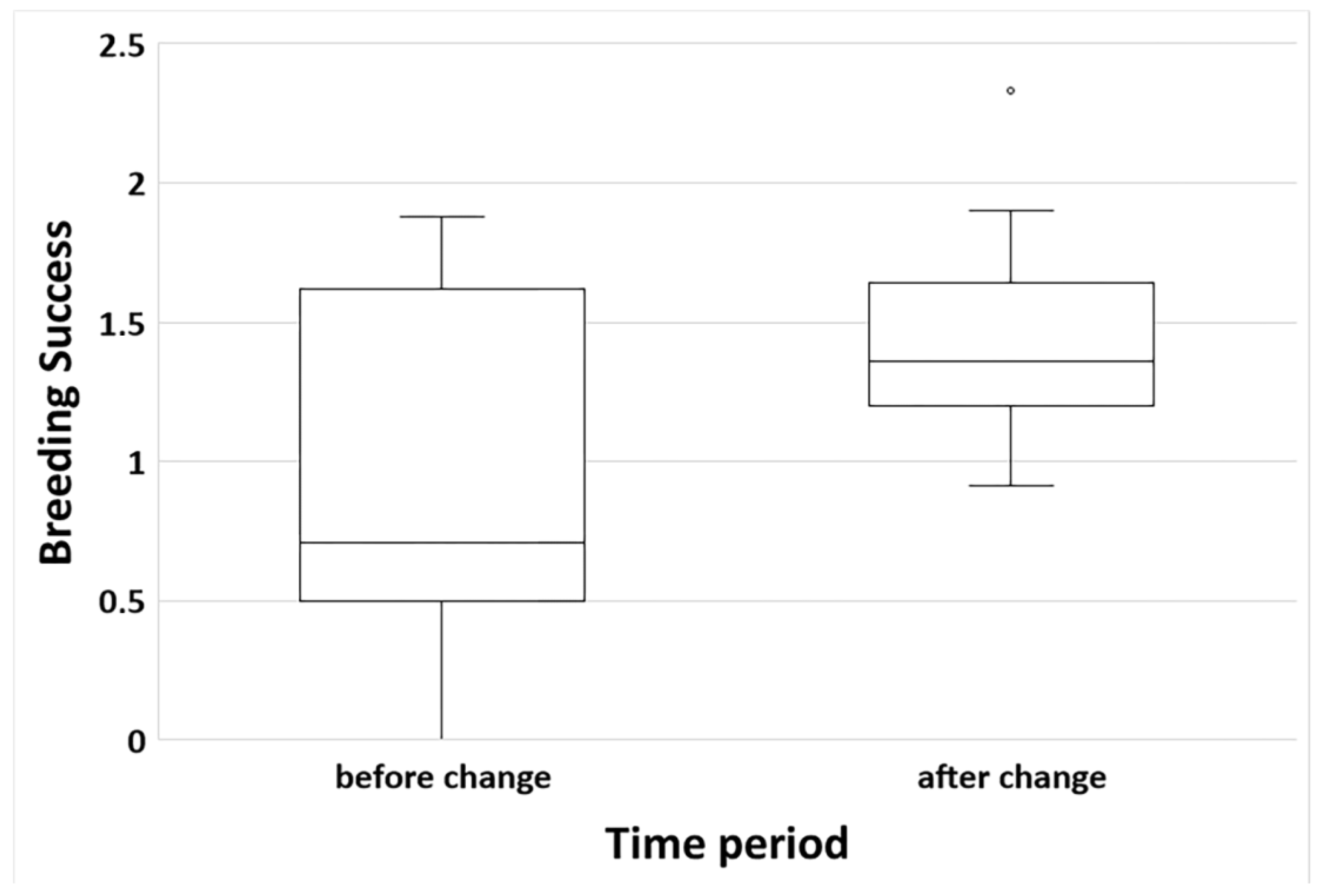
3. Results
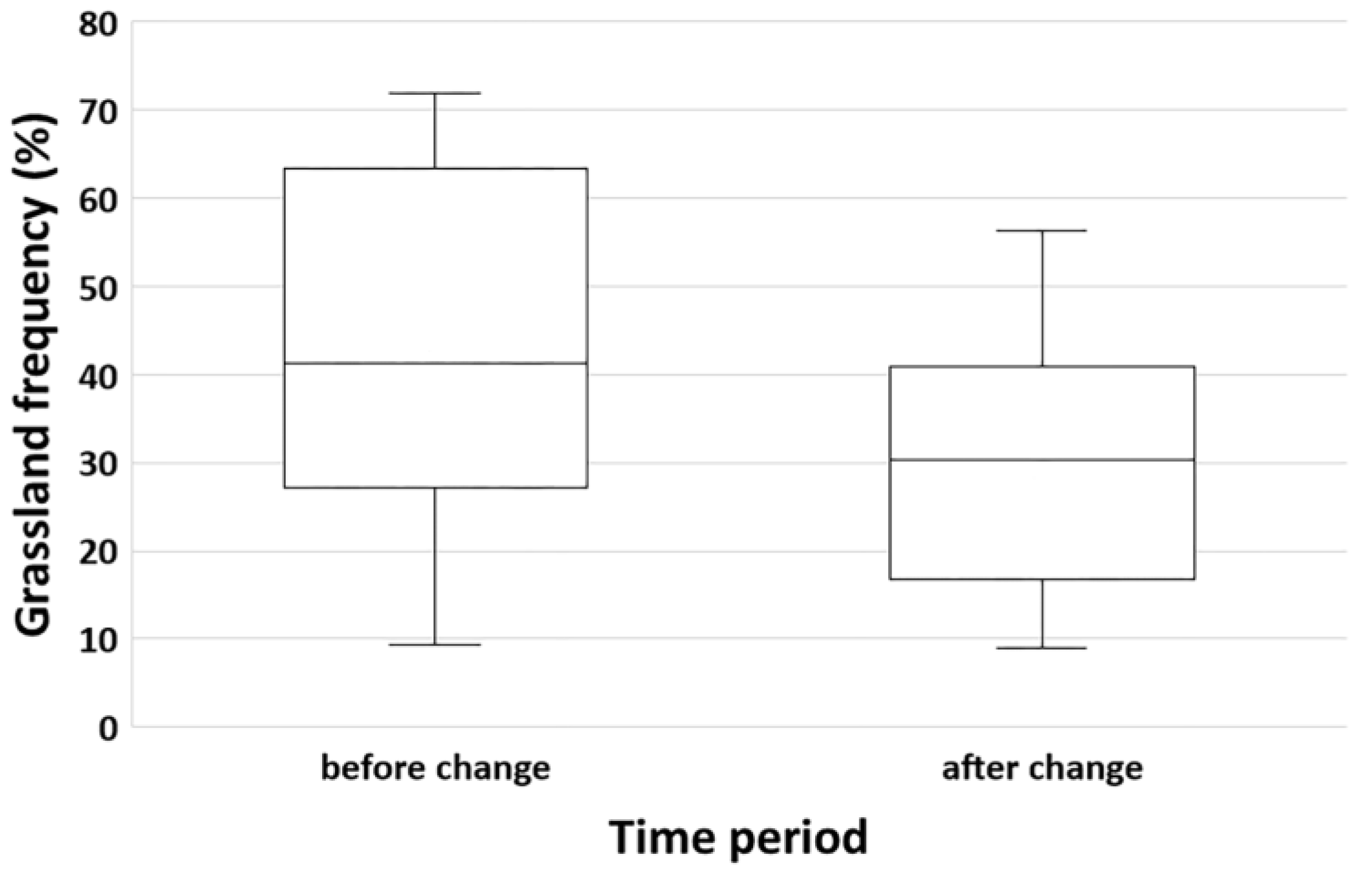
3.1. Grassland Alteration
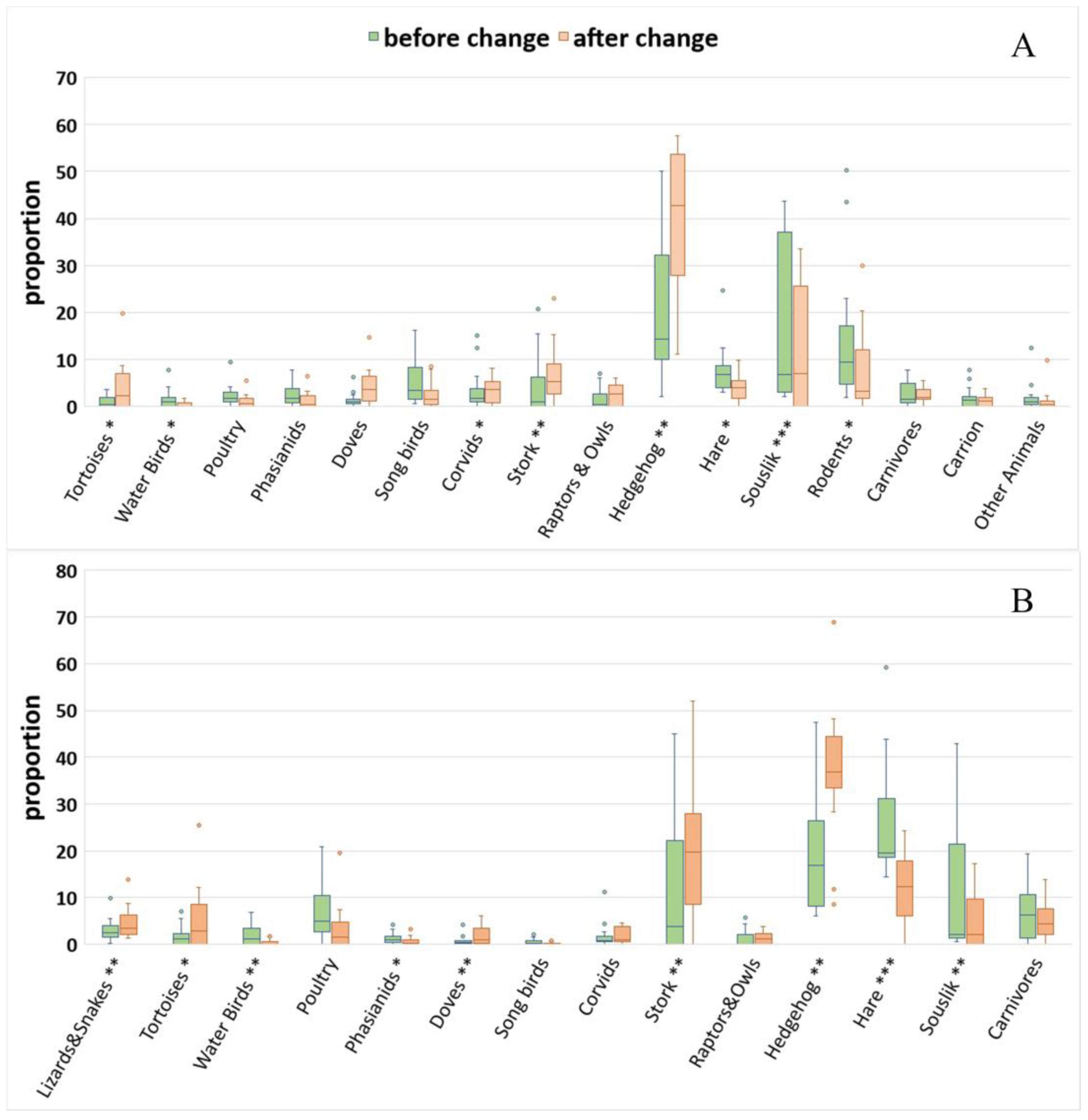
3.2. Dietary Shifts
3.3. Modeling General Effect of Dietary Alteration
3.4. Threshold Effect of Grassland Change in Individual Territory
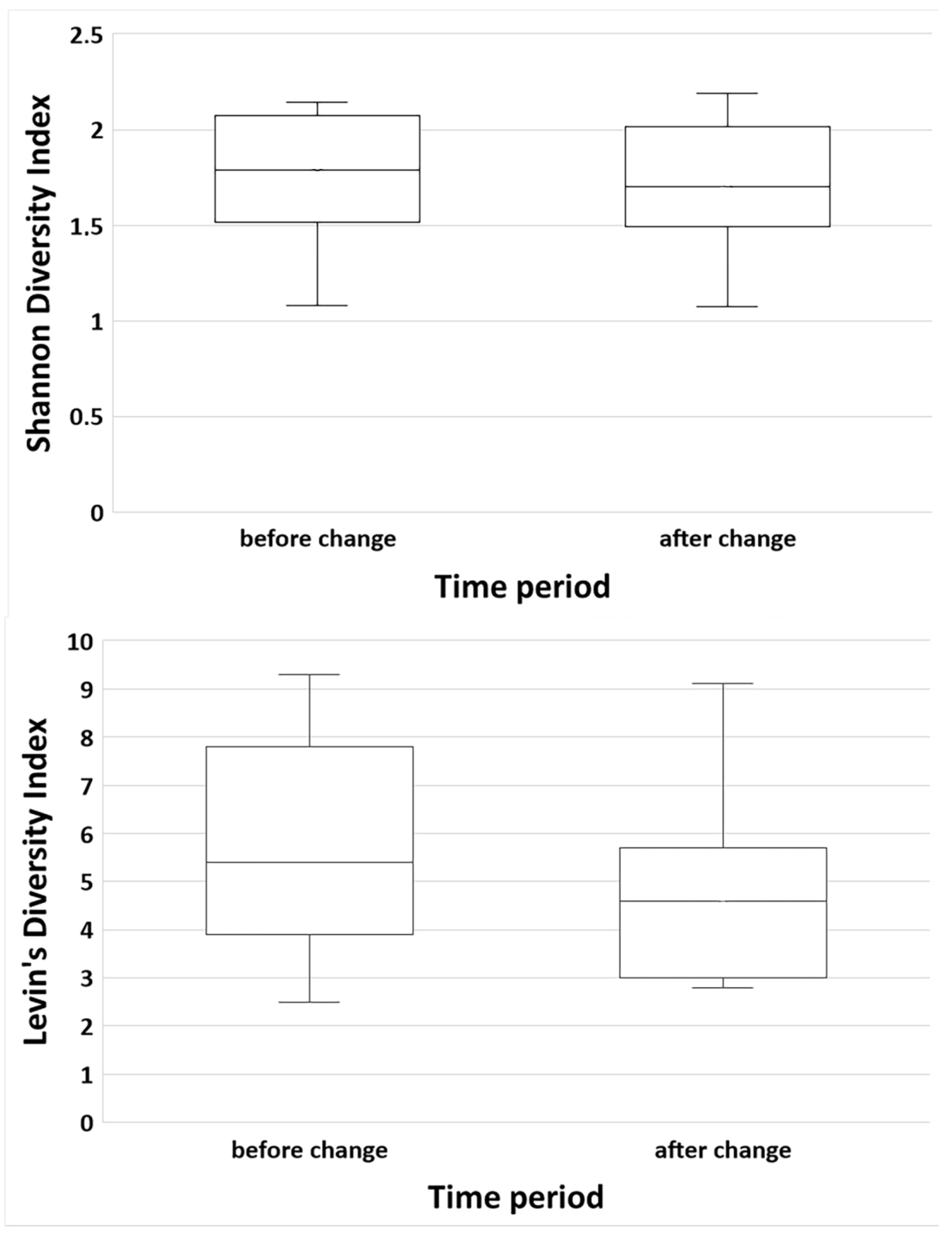
3.5. Habitat Diversity and Diet Diversity
3.6. Breeding Success and Effect of Grassland Transformation and Dietary Shifts
4. Discussion
Conservation Consequences
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Donald, P.F.; Green, R.E.; Heath, M.F. Agricultural intensification and the collapse of Europe’s farmland bird populations. Proc. R. Soc. Lond. B 2001, 268, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, I. The recent declines of farmland bird populations in Britain: An appraisal of causal factors and conservation actions. Ibis 2004, 146, 579–600. [Google Scholar] [CrossRef]
- Bowler, D.E.; Heldbjerg, H.; Fox, A.D.; O’Hara, R.; Böhning-Gaese, K. Disentangling the effects of multiple environmental drivers on population changes within communities. J. Anim. Ecol. 2018, 87, 1034–1045. [Google Scholar] [CrossRef] [PubMed]
- Sergio, F.; Newton, I.; Marchesi, L.; Pedrini, P. Ecologically justified charisma: Preservation of top predators delivers biodiversity conservation. J. Appl. Ecol. 2006, 43, 1049–1055. [Google Scholar] [CrossRef] [Green Version]
- Donàzar, J.A.; Cortés-Avizanda, A.; Fargallo, J.A.; Margalida, A.; Moleón, M.; Morales-Reyes, Z.; Moreno-Opo, R.; Pérez-García, J.M.; Sánchez-Zapata, J.A.; Zuberogoitia, I.; et al. Roles of raptors in a changing world: From flagships to providers of key ecosystem services. Ardeola 2016, 63, 181–234. [Google Scholar] [CrossRef] [Green Version]
- Sergio, F.; Caro, T.; Brown, D.; Clucas, B.; Hunter, J.; Ketchum, J.; Mchugh, K.; Hiraldo, F. Top predators as conservation tools: Ecological rationale, assumptions, and efficacy. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Flynn, D.F.B.; Gogol-Prokurat, M.; Nogeire, T.; Molinari, N.; Richers, B.T.; Lin, B.B.; Simpson, N.; Mayfield, M.M.; De Clerck, F. Loss of functional diversity under land use intensification across multiple taxa. Ecol. Lett. 2009, 12, 22–33. [Google Scholar] [CrossRef]
- Herrera, J.M.; Costa, P.; Medinas, D.; Marques, J.T.; Mira, A. Community composition and activity of insectivorous bats in Mediterranean olive farms. Anim. Conserv. 2015, 18, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Cardador, L.; Carrete, M.; Mañosa, S. Can intensive agricultural landscapes favour some raptor species? The Marsh Harrier in north-eastern Spain. Anim. Conserv. 2011, 14, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Buij, R.; Kortekaas, K.; van Krimpen, R.R.D.; van Wijk, R.; van der Zanden, S.; De Iongh, H.H.; Heitkönig, I.M.A.; de Snoo, G.R.; Komdeur, J. Breeding performance of the Grasshopper Buzzard (Butastur rufipennis) in a natural and a humanmodified West African savanna. Condor 2013, 115, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Murgatroyd, M.; Avery, G.; Underhill, L.G.; Amar, A. Adaptability of a specialist predator: The effects of land use on diet diversification and breeding performance of Verreaux’s eagles. J. Avian Biol. 2016, 47, 1–12. [Google Scholar] [CrossRef]
- Thiollay, J.M.; Clobert, J. Comparative foraging adaptations of small raptors in a dense African savanna. Ibis 1990, 132, 42–57. [Google Scholar] [CrossRef]
- Sánchez-Zapata, J.A.; Carrete, M.; Gravilov, A.; Sklyarenko, S.; Ceballos, O.; Donázar, J.A.; Hiraldo, F. Land use changes and raptor conservation in steppe habitats of eastern Kazakhstan. Biol. Conserv. 2003, 111, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Demerdzhiev, D.; Dobrev, D.; Popgeorgiev, G.; Stoychev, S. Landscape alteration affects the demography of an endangered avian predator by reducing the habitat quality. Avian Res. 2022, 13, 100030. [Google Scholar] [CrossRef]
- Donázar, J.A.; Hiraldo, F.; Bustamante, J. Factors influencing nest site selection, breeding density and breeding success in the bearded vulture (Gypaetus barbatus). J. Appl. Ecol. 1993, 30, 504–514. [Google Scholar] [CrossRef]
- Jenkins, C.N.; Pimm, S.L.; Joppa, L.N. Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl. Acad. Sci. USA 2013, 110, 2602–2610. [Google Scholar] [CrossRef] [Green Version]
- Grande, J.M.; Orozco-Valor, P.M.; Liébana, M.S.; Sarasola, J.H. Birds of prey in agricultural landscapes: The role of agriculture expansion and intensification. In Birds of Prey: Biology and Conservation in the XXI Centuryology and Conservation in the XXI Century; Sarasola, J., Grande, J., Negro, J., Eds.; Springer: New York, NY, USA, 2018; pp. 197–228. [Google Scholar]
- Heath, J.A.; Kochert, M.N.; Steenhof, K. Golden Eagle dietary shifts following wildfire and shrub loss have negative consequences for nestling survivorship. Ornithol. Appl. 2021, 123, duab034. [Google Scholar] [CrossRef]
- Fuller, R.J.; Gough, S.J. Changes in sheep numbers in Britain: Implications for bird populations. Biol. Conserv. 1999, 91, 73–89. [Google Scholar] [CrossRef]
- McClure, C.J.W.; Westrip, J.R.S.; Johnson, J.A.; Schulwitz, S.E.; Virani, M.Z.; Davies, R.; Symes, A.; Wheatley, H.; Thorstrom, R.; Amar, A.; et al. State of the world’s raptors: Distributions, threats, and conservation recommendations. Biol. Conser. 2018, 227, 390–402. [Google Scholar] [CrossRef]
- Tella, J.L.; Forero, M.G.; Hiraldo, F.; Donázar, J.A. Conflicts between Lesser Kestrel Conservation and European Agricultural Policies as Identified by Habitat Use Analyses. Conserv. Biol. 1998, 12, 593–604. [Google Scholar] [CrossRef] [Green Version]
- Catry, I.; Amano, T.; Franco, A.M.A.; Sutherland, W.J. Influence of spatial and, temporal dynamics of agricultural practices on the lesser kestrel. J. Appl. Ecol. 2012, 49, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Arroyo, B.; Garcia, J.T.; Bretagnolle, V. Conservation of the Montagu’s Harrier (Circus pygargus) in agricultural areas. Anim. Conser. 2002, 5, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Demerdzhiev, D.; Dobrev, V.; Popgeorgiev, G. Effects of habitat change on territory occupancy, breeding density and breeding success of Long-legged Buzzard (Buteo rufinus, Cretzschmar, 1927) in Besaparski riodove special protection area (Natura 2000), Southern Bulgaria. Acta Zool. Bulg. 2014, 5, 191–200. [Google Scholar]
- Demerdzhiev, D.; Angelov, I.; Dobrev, D. Foraging Patterns of Non-Territorial Eastern Imperial Eagle (Aquila heliaca): A Case of Successful Adaptation. Diversity 2022, 14, 1060. [Google Scholar] [CrossRef]
- Demerdzhiev, D.A.; Horváth, M.; Kovács, A.; Stoychev, S.A.; Karyakin, I.V. Status and population trend of the eastern imperial eagle (Aquila heliaca) in Europe in the period 2000–2010. Acta Zool. Bulg. 2011, 3, 5–14. [Google Scholar]
- Demerdzhiev, D.; Boev, Z.; Dobrev, D.; Nedyalkov, N.; Petrov, T. Does Temporal and Spatial Diet Alteration Lead to Successful Adaptation of the Eastern Imperial Eagle, a Top Predator? Diversity 2022, 14, 1000. [Google Scholar] [CrossRef]
- Demerdzhiev, D.; Stoychev, S.; Dobrev, D.; Spasov, S.; Terziev, N. Conservation measures undertaken to improve the population status of eastern imperial eagle (Aquila heliaca) in Bulgaria. Slovak Raptor J. 2014, 8, 27–39. [Google Scholar] [CrossRef]
- Lazarova, I.; Dobrev, D.; Gradev, G.; Petrov, R.; Stoychev, S.; Klisurov, I.; Demerdzhiev, D. Main mortality factors for the Eastern Imperial Eagle (Aquila heliaca Savigny, 1809) in Bulgaria. Ornis Hung. 2020, 28, 120–134. [Google Scholar] [CrossRef]
- Demerdzhiev, D.; Boev, Z.; Dobrev, D.; Terziev, N.; Nedyalkov, N.; Stoychev, S.; Petrov, T. Diet of Eastern Imperial Eagle (Aquila heliaca) in Bulgaria: Composition, distribution and variation. Biodivers. Data J. 2022, 10, e77746. [Google Scholar] [CrossRef]
- Pyke, G.; Pulliam, H.R.; Charnov, E.L. Optimal Foraging: A Selective Review of Theory and Tests. Q. Rev. Biol. 1977, 52, 137–154. [Google Scholar] [CrossRef] [Green Version]
- Stephens, D.W.; Krebs, J.R. Foraging Theory; Princeton University Press: Princeton, NJ, USA, 1986. [Google Scholar]
- Simpson, E.H. Measure of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Katzner, T.; Bragin, E.; Knick, S.; Smith, A. Spatial structure in the diet of Eastern Imperial Eagle Aquila heliaca in Kazakhstan. J. Avian Biol. 2006, 37, 594–600. [Google Scholar] [CrossRef]
- Horváth, M.; Solti, B.; Fatér, I.; Juhász, T.; Haraszthy, L.; Szitta, T.; Ballók, Z.; Pásztory-Kovács, S. Temporal changes in the diet composition of the Eastern Imperial Eagle (Aquila heliaca) in Hungary. Ornis Hung. 2018, 26, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Horváth, M.; Szitta, T.; Firmánszky, G.; Solti, B.; Kovács, A.; Moskát, C. Spatial variation in prey composition and its possible effect on reproductive success in an expanding Eastern Imperial Eagle (Aquila heliaca) population. Acta Zool. Acad. Sci. Hung. 2010, 56, 187–200. [Google Scholar]
- Newton, I. Population Ecology of Raptors; T. And A.D. Poyser: Berkhamsted, UK, 1979. [Google Scholar]
- ESRI. ArcGIS Desktop: Release 10.4.1; Environmental Systems Research Institute: Redlands, CA, USA, 2016. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Katzner, T.; Bragin, E.; Knick, S.; Smith, A. Relationship between demographics and diet specificity of Eastern Imperial Eagle Aquila heliaca in Kazakhstan. Ibis 2005, 147, 576–586. [Google Scholar] [CrossRef]
- Marti, C.D.; Bechard, M.; Jaksic, F.M. Food habits. In Raptor Research and Management Techniques; Bird, D.M., Bildstein, K.L., Eds.; Hancock House: Blaine, WA, USA, 2007. [Google Scholar]
- Demerdzhiev, D. Eastern Imperial Eagle (Aquila heliaca heliaca Savigny, 1809) (Accipitridae—Aves) in Bulgaria—Distribution, Biology, Ecology, Numbers, and Conservation Measures. Ph.D. Thesis, BAS-NMNH, Sofia, Bulgaria, 2011. (In Bulgarian, with English Summary). [Google Scholar]
- Demerdzhiev, D.A.; Stoychev, S.S.; Dobrev, D.D.; Spasov, S.D.; Oppel, S. Studying the demographic drivers of an increasing Imperial Eagle population to inform conservation management. Biod. Conser. 2015, 24, 627–639. [Google Scholar] [CrossRef]
- Vili, N.; Szabó, K.; Kovács, S.; Kabai, P.; Kalmár, L.; Horváth, M. High turnover rate revealed by noninvasive genetic analyses in an expanding Eastern Imperial Eagle population. Acta Zool. Acad. Sc. Hung. 2013, 59, 279–295. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95364-7. [Google Scholar]
- Shapiro, S.; Wilk, M.; Chen, H. A comparative study of various tests of normality. J. Am. Stat. Assoc. 1968, 63, 1343–1372. [Google Scholar] [CrossRef]
- White, H.A. Heteroskedasticity-consistent covariance matrix estimator and a direct test for heteroskedasticity. Econometrica 1980, 48, 817–838. [Google Scholar] [CrossRef]
- StatSoft Inc. STATISTICA (Data Analysis Software System), Version 12. 2013. Available online: www.statsoft.com (accessed on 24 October 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics software package for education and data analysis. Paleont. Elec. 2001, 4, 9. [Google Scholar]
- Dobrev, V.D.; Popgeorgiev, G.S.; Plachiyski, D.G. Effects of the Common Agricultural Policy on the coverage of grassland habitats in Besaparski Ridove Special Protection Area (Natura 2000), southern Bulgaria. Acta Zool. Bulg. 2014, 5, 147–155. [Google Scholar]
- Coates, P.S.; Howe, K.B.; Casazza, M.L.; Delehanty, D.J. Landscape alterations influence differential habitat use of nesting buteos and ravens within sagebrush ecosystem: Implications for transmission line development. Condor 2014, 116, 341–356. [Google Scholar] [CrossRef]
- Zhelev, C.H.; Ninov, N.; Mihaylov, H.; Gruychev, G.; Stoyanov, S.; Mirchev, R. Density of Brown Hare (Lepus europaeus Pallas, 1778) in the plain habitats of Bulgaria. In Proceedings of the 2nd International Symposium on Hunting, Modern Aspects of Sustainable Management of Game Populations, Novi Sad, Serbia, 17–20 October 2013. [Google Scholar]
- Milchev, B. Diet shifting of tortoise-eating Golden Eagles (Aquila chrysaetos) in southeastern Bulgaria. Ornis Fenn. 2022, 99, 60–70. [Google Scholar] [CrossRef]
- Milchev, B.; Georgiev, V. Temporal and spatial dietary shifts of a generalist top predator: Long-term study of an Eagle Owl (Bubo bubo) population. For. Ideas 2020, 26, 366–379. [Google Scholar]
- Svanbäck, R.; Bolnick, D.I. Intraspecific competition affects the strength of individual specialization: An optimal diet theory method. Evol. Ecol. Res. 2005, 7, 993–1012. [Google Scholar]
- Moleón, M.; Sánchez-Zapata, J.A.; Gil-Sánchez, J.M.; Ballesteros-Duperón, E.; Barea-Azcón, J.M.; Virgós, E. Predator-prey relationships in a Mediterranean vertebrate system: Bonelli’s eagles, rabbits and partridges. Oecologia 2012, 168, 679–689. [Google Scholar] [CrossRef]
- Penteriani, V.; Fortuna, M.A.; Melian, C.J.; Otalora, F.; Ferrer, M. Can prey behaviour induce spatially synchronic aggregation of solitary predators? Oikos 2006, 113, 497–505. [Google Scholar] [CrossRef]
- Lourenço, R.; Delgado, M.D.M.; Campioni, L.; Korpimäki, E.; Penteriani, V. Evaluating the influence of diet-related variables on breeding performance and home range behaviour of a top predator. Popul. Ecol. 2015, 57, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Clouet, M.; Gerard, J.-F.; Goar, J.-L.; Goulard, M.; González, L.; Rebours, I.; Faure, C. Diet and Breeding Performance of the Golden Eagle Aquila chrysaetos at the Eastern and Western Extremities of the Pyrenees: An Example of Intra-Population Variability. Ardeola 2017, 64, 347–361. [Google Scholar] [CrossRef]
- Bragin, E. On the demography of the Imperial Eagle Aquila heliaca in Kazakhstan. In Raptors at Risk: City Berlin State; Chancellor, R., Meyburg, B.U., Eds.; WWGBP/Hancock House: Berlin, Germany, 2000; pp. 409–413. [Google Scholar]
- Karyakin, I.; Nikolenko, E.; Levin, A.; Kovalenko, A. Imperial Eagle in Russia and Kazakhstan: Population Status and Trends. Raptors Conserv. 2008, 14, 19–27. [Google Scholar]
- Väli, Ü. Factors limiting reproductive performance and nestling sex ratio in the Lesser Spotted Eagle Aquila pomarina at the northern limit of its range: The impact of weather and prey abundance. Acta Ornithol. 2012, 47, 157–168. [Google Scholar] [CrossRef]
- Demerdzhiev, D. Breeding parameters and factors infuencing the reproduction of an expanding Long-legged Buzzard (Buteo rufnus) population under high breeding density conditions. J. Ornithol. 2022, 163, 405–415. [Google Scholar] [CrossRef]
- López-López, P.; García-Ripollés, C.; Soutullo, A.; Cadahía, L.; Urios, V. Identifying potentially suitable nesting habitat for Golden Eagles applied to ‘important bird areas’ design. Anim. Conserv. 2007, 10, 208–218. [Google Scholar] [CrossRef]
- Horváth, M.; Szitta, T.; Bagyura, J.; Fatér, I.; Firmánszky, G.; Moskát, C. Simultaneous effect of habitat and age on reproductive success of Imperial Eagles (Aquila heliaca) in Hungary. Or. Hung. 2014, 22, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Balbontín, J.; Penteriani, V.; Ferrer, M. Variation in the age of mates as an early warning signal of changes in population trends? The case of Bonelli’s Eagle in Andalusia. Biol. Conser. 2003, 109, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, M.; Bisson, I. Age and territory quality effects on fecundity in Spanish imperial eagle (Aquila adalberti). Auk 2003, 120, 180–186. [Google Scholar] [CrossRef]
- Margalida, A.; Mañosa, S.; Gonzalez, L.M.; Ortega, E.; Sanchez, R.; Oria, J. Breeding of non-adults and effect of age on productivity in the Spanish Imperial Eagle Aquila adalberti. Ardea 2008, 96, 173–180. [Google Scholar] [CrossRef] [Green Version]
- MacArthur, R.H.; Pianka, E.R. On optimal use of a patchy environment. Am. Nat. 1966, 100, 603–609. [Google Scholar] [CrossRef] [Green Version]
- Kramer, D.L. ‘Foraging Behavior’. In Evolutionary Ecology: Concepts and Case Studies; Fox, C.W., Roff, D.A., Fairbairn, D.J., Eds.; Oxford Academic: Chenango, NY, USA, 2001. [Google Scholar]
- Dunning, J. (Ed.) Handbook of Avian Body Masses; CRC Press: Abingdon, UK, 2008. [Google Scholar]
- Aulagner, S.; Haffner, R.; Mitchell-Jones, A.; Moutou, F.; Zima, J. Mammals of Europe, North Africa and the Middle East; Christopher Helm: London, UK, 2009. [Google Scholar]
- Petrov, P. Variability in the body mass of European Hare (Lepus europaeus, Pall.) in Bulgaria. For. Sci. 1964, 1, 73–81. (In Bulgarian) [Google Scholar]
- Real, J.; Mañosa, S.; Muñoz, E. Trichomoniasis in a Bonelli’s eagle population in Spain. J. Wildl. Dis. 2000, 36, 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudek, B.M.; Kochert, M.N.; Barnes, J.G.; Bloom, P.H.; Papp, J.M.; Gerhold, R.W.; Purple, K.E.; Jacobson, K.V.; Preston, C.R.; Vennum, C.R.; et al. Prevalence and risk factors of Trichomonas gallinae and Trichomonosis in Golden Eagle (Aquila chrysaetos) nestlings in Western North America. J. Wildl. Dis. 2018, 54, 755–764. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Factor | Effect | df | F | p |
|---|---|---|---|---|---|
| Grassland abundance | Territory | Random | 14 | 10.56 | <0.001 |
| Grassland abundance | Period | Fixed | 1 | 20.69 | <0.001 |
| H | Territory | Random | 14 | 43.88 | <0.001 |
| H | Period | Fixed | 1 | 4.88 | 0.04 |
| FNB | Territory | Random | 14 | 0.50 | 0.90 |
| FNB | Period | Fixed | 1 | 1.61 | 0.23 |
| Breeding success | Territory | Random | 14 | 0.96 | 0.53 |
| Breeding success | Period | Fixed | 1 | 7.73 | 0.01 |
| Categories | Frequency of Prey Contribution | Biomass of Prey Contribution | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Factor | Effect | df | F | p | Factor | Effect | df | F | p | |
| Tortoises | Territory | Random | 14 | 2.81 | 0.03 | Territory | Random | 14 | 2.73 | 0.04 |
| Tortoises | Period | Fixed | 1 | 7.53 | 0.02 | Period | Fixed | 1 | 5.43 | 0.04 |
| Water birds | Territory | Random | 14 | 2.33 | 0.06 | Territory | Random | 14 | 3.24 | 0.02 |
| Water birds | Period | Fixed | 1 | 9.53 | 0.008 | Period | Fixed | 1 | 16.02 | 0.001 |
| Poultry | Territory | Random | 14 | 0.43 | 0.94 | Territory | Random | 14 | 0.51 | 0.89 |
| Poultry | Period | Fixed | 1 | 3.22 | 0.09 | Period | Fixed | 1 | 2.48 | 0.14 |
| Phasianids | Territory | Random | 14 | 1.70 | 0.17 | Territory | Random | 14 | 2.65 | 0.04 |
| Phasianids | Period | Fixed | 1 | 6.93 | 0.02 | Period | Fixed | 1 | 8.51 | 0.01 |
| Doves | Territory | Random | 14 | 1.85 | 0.13 | Territory | Random | 14 | 3.75 | 0.009 |
| Doves | Period | Fixed | 1 | 6.39 | 0.02 | Period | Fixed | 1 | 5.62 | 0.03 |
| Songbirds | Territory | Random | 14 | 1.37 | 0.28 | Territory | Random | 14 | 1.85 | 0.13 |
| Songbirds | Period | Fixed | 1 | 6.52 | 0.02 | Period | Fixed | 1 | 4.37 | 0.055 |
| Corvids | Territory | Random | 14 | 2.71 | 0.04 | Territory | Random | 14 | 2.16 | 0.08 |
| Corvids | Period | Fixed | 1 | 0.002 | 0.97 | Period | Fixed | 1 | 0.007 | 0.94 |
| Stork | Territory | Random | 14 | 4.62 | 0.004 | Territory | Random | 14 | 4.14 | 0.006 |
| Stork | Period | Fixed | 1 | 7.79 | 0.01 | Period | Fixed | 1 | 9.61 | 0.008 |
| Raptors and owls | Territory | Random | 14 | 1.56 | 0.21 | Territory | Random | 14 | 2.58 | 0.04 |
| Raptors and owls | Period | Fixed | 1 | 1.48 | 0.24 | Period | Fixed | 1 | 0.50 | 0.49 |
| Hedgehog | Territory | Random | 14 | 3.35 | 0.02 | Territory | Random | 14 | 2.93 | 0.03 |
| Hedgehog | Period | Fixed | 1 | 23.48 | <0.001 | Period | Fixed | 1 | 22.25 | <0.001 |
| Hare | Territory | Random | 14 | 2.44 | 0.054 | Territory | Random | 14 | 4.88 | 0.003 |
| Hare | Period | Fixed | 1 | 14.09 | 0.002 | Period | Fixed | 1 | 43.58 | <0.001 |
| Souslik | Territory | Random | 14 | 5.76 | 0.001 | Territory | Random | 14 | 4.64 | 0.003 |
| Souslik | Period | Fixed | 1 | 6.39 | 0.02 | Period | Fixed | 1 | 7.31 | 0.02 |
| Carnivores | Territory | Random | 14 | 0.69 | 0.75 | Territory | Random | 14 | 0.47 | 0.91 |
| Carnivores | Period | Fixed | 1 | 0.04 | 0.84 | Period | Fixed | 1 | 0.16 | 0.70 |
| Carrion | Territory | Random | 14 | 1.11 | 0.42 | Not evaluated | ||||
| Carrion | Period | Fixed | 1 | 0.27 | 0.61 | Not evaluated | ||||
| Rodents | Territory | Random | 14 | 2.92 | 0.03 | Excluded due the heteroscedasticity | ||||
| Rodents | Period | Fixed | 1 | 6.87 | 0.02 | Excluded due the heteroscedasticity | ||||
| Other animals | Territory | Random | 14 | 1.27 | 0.33 | Excluded due the heteroscedasticity | ||||
| Other animals | Period | Fixed | 1 | 2.41 | 0.14 | Excluded due the heteroscedasticity | ||||
| Lizards and snakes | Excluded due the heteroscedasticity | Territory | Random | 14 | 3.76 | 0.009 | ||||
| Lizards and snakes | Excluded due the heteroscedasticity | Period | Fixed | 1 | 2.18 | 0.16 | ||||
| Gulls | Excluded due the heteroscedasticity | Excluded due the heteroscedasticity | ||||||||
| Gulls | Excluded due the heteroscedasticity | Excluded due the heteroscedasticity | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demerdzhiev, D.A.; Dobrev, D.D.; Boev, Z.N. Grassland Alterations Do Not Affect Breeding Success, but Can Explain Dietary Shifts of a Generalist Raptor Species. Diversity 2023, 15, 422. https://0-doi-org.brum.beds.ac.uk/10.3390/d15030422
Demerdzhiev DA, Dobrev DD, Boev ZN. Grassland Alterations Do Not Affect Breeding Success, but Can Explain Dietary Shifts of a Generalist Raptor Species. Diversity. 2023; 15(3):422. https://0-doi-org.brum.beds.ac.uk/10.3390/d15030422
Chicago/Turabian StyleDemerdzhiev, Dimitar Atanasov, Dobromir Damyanov Dobrev, and Zlatozar Nikolaev Boev. 2023. "Grassland Alterations Do Not Affect Breeding Success, but Can Explain Dietary Shifts of a Generalist Raptor Species" Diversity 15, no. 3: 422. https://0-doi-org.brum.beds.ac.uk/10.3390/d15030422