
Spatial Patterns of Neutral and Functional Genetic Variations along Dendritic Networks of Riverscape in Brown Trout Populations

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
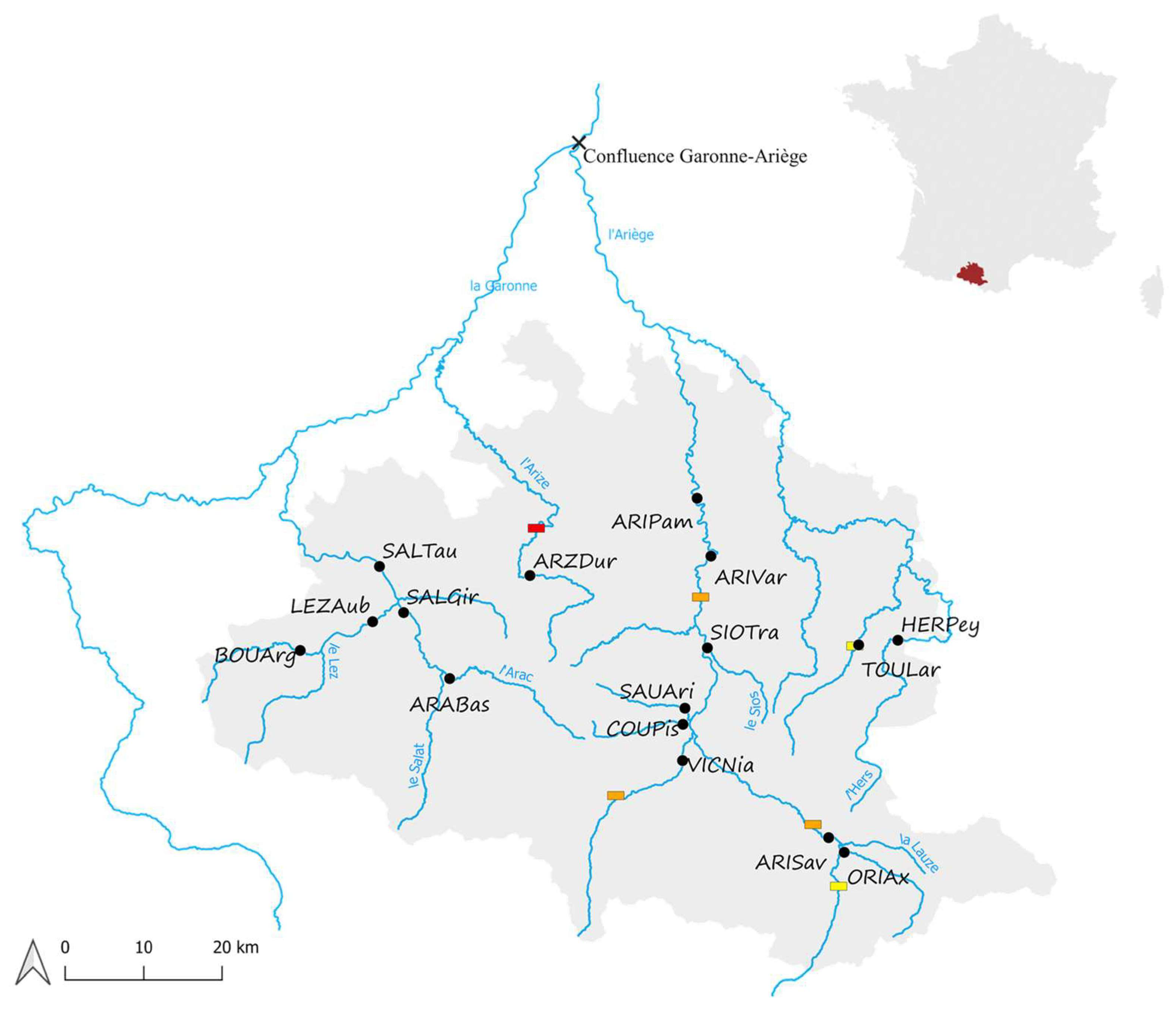
2.1. Study Areas and Fish Sampling
2.2. Spatial Data
2.3. Genotyping of Neutral and Functional Genetic Markers
2.4. Genetic Analyses
2.5. Statistical Analyses
3. Results
3.1. Robustness and Descriptive Genetic Statistics
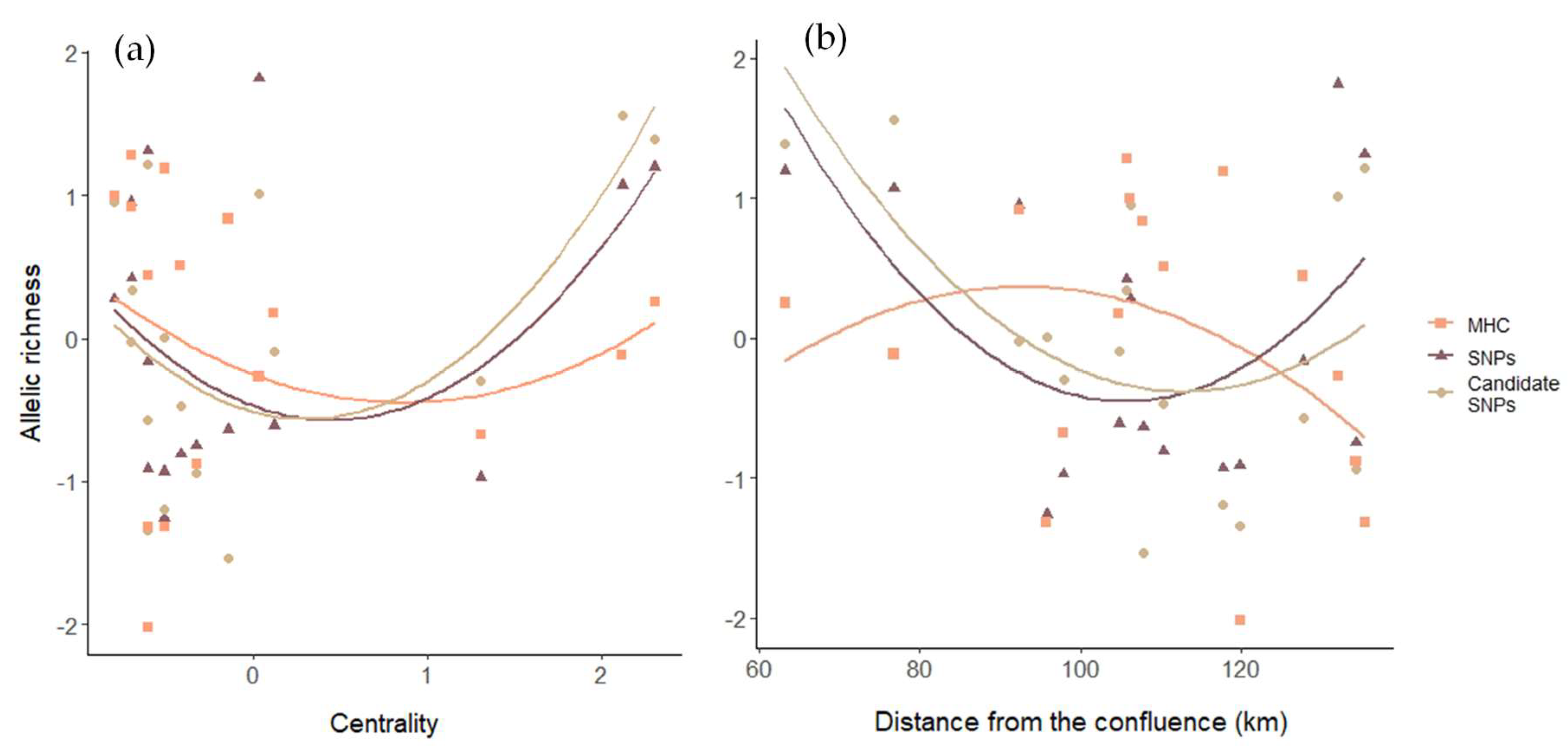
3.2. Genetic Diversity
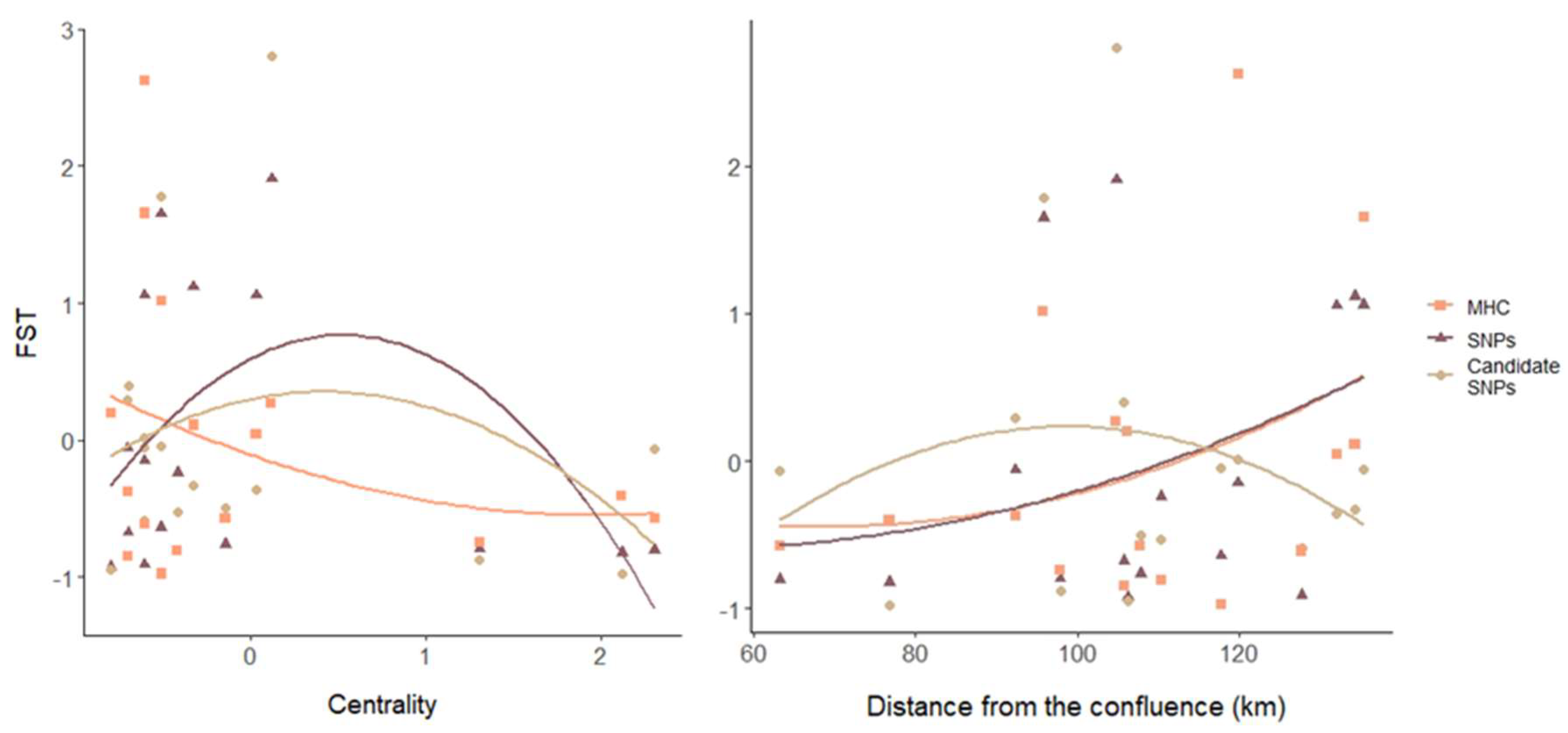
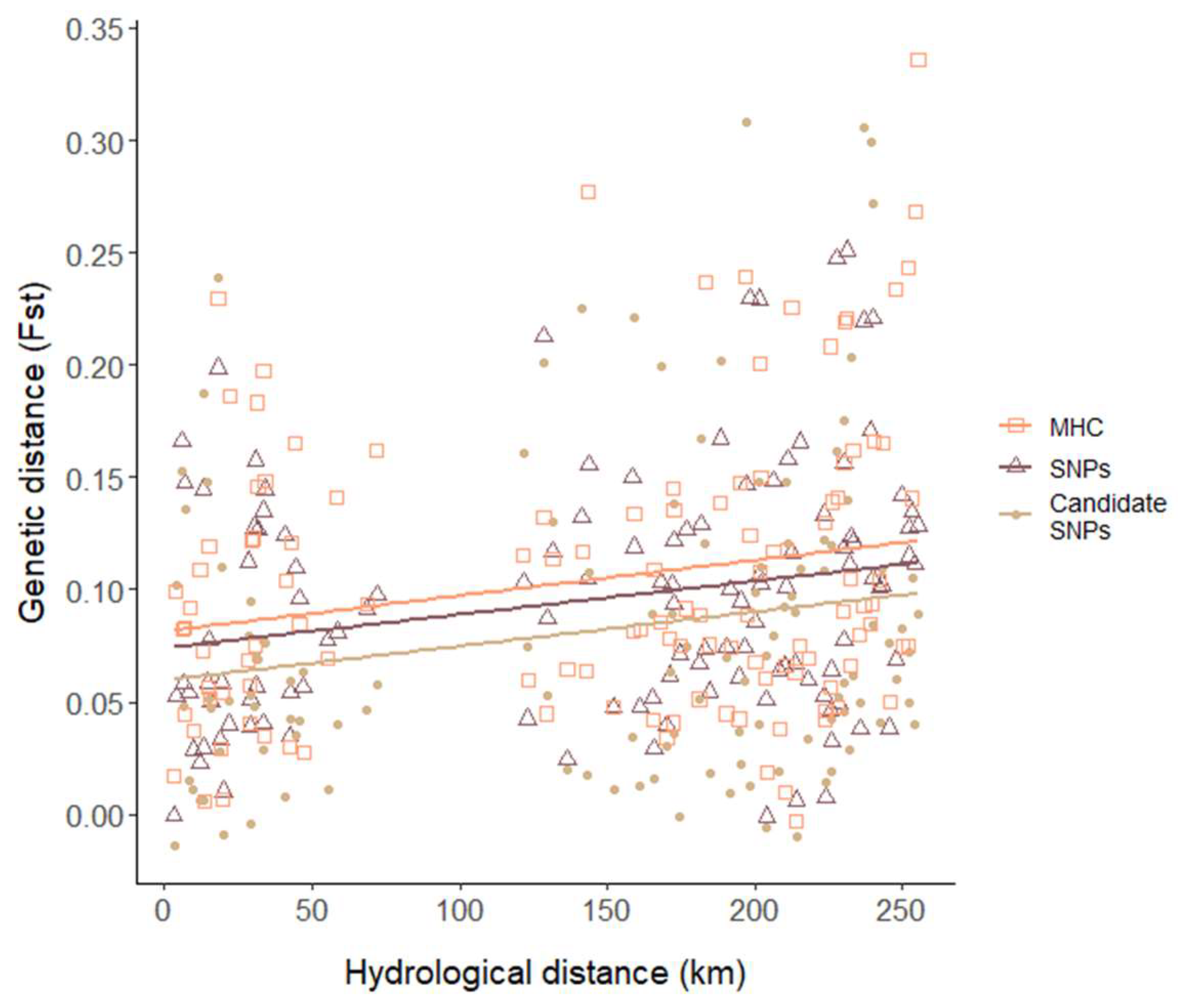
3.3. Genetic Differentiation
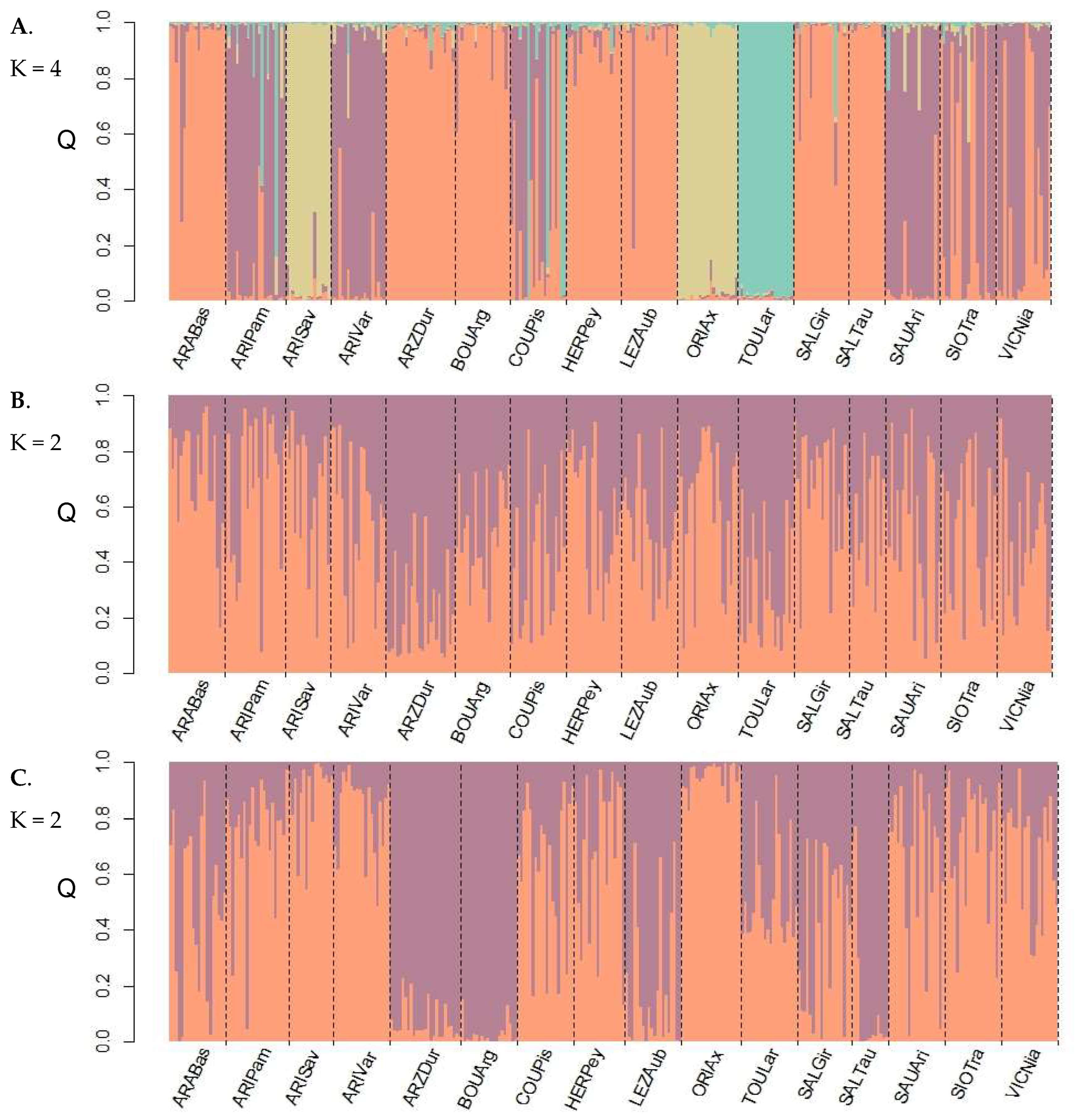
3.4. Population Genetic Structure
4. Discussion
4.1. Environmental Gradients Shape Spatial Patterns of Genetic Diversity and Differentiation
4.2. Different Genetic Markers Can Exhibit a Similar Pattern
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Legendre, P.; Fortin, M.J. Spatial Pattern and Ecological Analysis. Vegetatio 1989, 80, 107–138. [Google Scholar] [CrossRef]
- Ortego, J.; Riordan, E.C.; Gugger, P.F.; Sork, V.L. Influence of Environmental Heterogeneity on Genetic Diversity and Structure in an Endemic Southern Californian Oak. Mol. Ecol. 2012, 21, 3210–3223. [Google Scholar] [CrossRef]
- Chave, J. The Problem of Pattern and Scale in Ecology: What Have We Learned in 20 Years? Ecol. Lett. 2013, 16, 4–16. [Google Scholar] [CrossRef]
- Wright, S. Isolation by Distance. Genetics 1943, 28, 114–138. [Google Scholar] [CrossRef]
- Hillebrand, H. On the Generality of the Latitudinal Diversity Gradient. Am. Nat. 2004, 163, 192–211. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Monaghan, M.T.; Omura, T. Longitudinal Patterns of Genetic Diversity and Larval Density of the Riverine Caddisfly Hydropsyche orientalis (Trichoptera). Aquat. Sci. 2008, 70, 377–387. [Google Scholar] [CrossRef]
- Lawrence, E.R.; Fraser, D.J. Latitudinal Biodiversity Gradients at Three Levels: Linking Species Richness, Population Richness and Genetic Diversity. Glob. Ecol. Biogeogr. 2020, 29, 770–788. [Google Scholar] [CrossRef] [Green Version]
- Blanchet, S.; Prunier, J.G.; Paz-Vinas, I.; Saint-Pé, K.; Rey, O.; Raffard, A.; Mathieu-Bégné, E.; Loot, G.; Fourtune, L.; Dubut, V. A River Runs through It: The Causes, Consequences, and Management of Intraspecific Diversity in River Networks. Evol. Appl. 2020, 13, 1195–1213. [Google Scholar] [CrossRef] [Green Version]
- Fagan, W.F. Connectivity, Fragmentation, and Extinction Risk in Dendritic Metapopulations. Ecology 2002, 83, 3243–3249. [Google Scholar] [CrossRef]
- Campbell Grant, E.H.; Lowe, W.H.; Fagan, W.F. Living in the Branches: Population Dynamics and Ecological Processes in Dendritic Networks. Ecol. Lett. 2007, 10, 165–175. [Google Scholar] [CrossRef]
- Morrissey, M.B.; de Kerckhove, D.T. The Maintenance of Genetic Variation Due to Asymmetric Gene Flow in Dendritic Metapopulations. Am. Nat. 2009, 174, 875–889. [Google Scholar] [CrossRef]
- Paz-Vinas, I.; Loot, G.; Stevens, V.M.; Blanchet, S. Evolutionary Processes Driving Spatial Patterns of Intraspecific Genetic Diversity in River Ecosystems. Mol. Ecol. 2015, 24, 4586–4604. [Google Scholar] [CrossRef] [Green Version]
- Paz-Vinas, I.; Blanchet, S. Dendritic Connectivity Shapes Spatial Patterns of Genetic Diversity: A Simulation-Based Study. J. Evol. Biol. 2015, 28, 986–994. [Google Scholar] [CrossRef]
- Cyr, F.; Angers, B. Historical Process Lead to False Genetic Signal of Current Connectivity among Populations. Genetica 2011, 139, 1417–1428. [Google Scholar] [CrossRef]
- Chaput-Bardy, A.; Fleurant, C.; Lemaire, C.; Secondi, J. Modelling the Effect of In-Stream and Overland Dispersal on Gene Flow in River Networks. Ecol. Model. 2009, 220, 3589–3598. [Google Scholar] [CrossRef] [Green Version]
- Alp, M.; Keller, I.; Westram, A.M.; Robinson, C.T. How River Structure and Biological Traits Influence Gene Flow: A Population Genetic Study of Two Stream Invertebrates with Differing Dispersal Abilities. Freshw. Biol. 2012, 57, 969–981. [Google Scholar] [CrossRef]
- Prunier, J.G.; Dubut, V.; Loot, G.; Tudesque, L.; Blanchet, S. The Relative Contribution of River Network Structure and Anthropogenic Stressors to Spatial Patterns of Genetic Diversity in Two Freshwater Fishes: A Multiple-Stressors Approach. Freshw. Biol. 2018, 63, 6–21. [Google Scholar] [CrossRef]
- Castric, V.; Bonney, F.; Bernatchez, L. Landscape Structure and Hierarchical Genetic Diversity in the Brook Charr, Salvelinus Fontinalis. Evolution 2001, 55, 1016–1028. [Google Scholar] [CrossRef]
- Finn, D.S.; Bonada, N.; Múrria, C.; Hughes, J.M. Small but Mighty: Headwaters Are Vital to Stream Network Biodiversity at Two Levels of Organization. J. N. Am. Benthol. Soc. 2011, 30, 963–980. [Google Scholar] [CrossRef] [Green Version]
- Prunier, J.G.; Dubut, V.; Chikhi, L.; Blanchet, S. Contribution of Spatial Heterogeneity in Effective Population Sizes to the Variance in Pairwise Measures of Genetic Differentiation. Methods Ecol. Evol. 2017, 8, 1866–1877. [Google Scholar] [CrossRef] [Green Version]
- Blanchet, S.; Rey, O.; Etienne, R.; Lek, S.; Loot, G. Species-Specific Responses to Landscape Fragmentation: Implications for Management Strategies. Evol. Appl. 2010, 3, 291–304. [Google Scholar] [CrossRef]
- Allen, A.P.; Gillooly, J.F.; Savage, V.M.; Brown, J.H. Kinetic Effects of Temperature on Rates of Genetic Divergence and Speciation. Proc. Natl. Acad. Sci. USA 2006, 103, 9130–9135. [Google Scholar] [CrossRef] [Green Version]
- Willis, G.H.; McDowell, L.L. Pesticides in Agricultural Runoff and Their Effects on Downstream Water Quality. Environ. Toxicol. Chem. 1982, 1, 267–279. [Google Scholar] [CrossRef]
- Dionne, M.; Miller, K.M.; Dodson, J.J.; Caron, F.; Bernatchez, L. Clinal Variation In MHC Diversity With Temperature: Evidence For The Role Of Host? Pathogen Interaction On Local Adaptation In Atlantic Salmon. Evolution 2007, 61, 2154–2164. [Google Scholar] [CrossRef]
- Hari, R.E.; Livingstone, D.M.; Siber, R.; Burkhardt-Holm, P.; Güttinger, H. Consequences of Climatic Change for Water Temperature and Brown Trout Populations in Alpine Rivers and Streams. Glob. Chang. Biol. 2006, 12, 10–26. [Google Scholar] [CrossRef]
- Elliott, J.M.; Elliott, J.A. Temperature Requirements of Atlantic Salmon Salmo Salar, Brown Trout Salmo trutta and Arctic Charr Salvelinus Alpinus: Predicting the Effects of Climate Change. J. Fish Biol. 2010, 77, 1793–1817. [Google Scholar] [CrossRef]
- Ojanguren, A.F.; Reyes-Gavilán, F.G.; Braña, F. Thermal Sensitivity of Growth, Food Intake and Activity of Juvenile Brown Trout. J. Therm. Biol. 2001, 26, 165–170. [Google Scholar] [CrossRef]
- Réalis-Doyelle, E.; Pasquet, A.; Charleroy, D.D.; Fontaine, P.; Teletchea, F. Strong Effects of Temperature on the Early Life Stages of a Cold Stenothermal Fish Species, Brown Trout (Salmo trutta L.). PLoS ONE 2016, 11, e0155487. [Google Scholar] [CrossRef] [Green Version]
- Borgwardt, F.; Unfer, G.; Auer, S.; Waldner, K.; El-Matbouli, M.; Bechter, T. Direct and Indirect Climate Change Impacts on Brown Trout in Central Europe: How Thermal Regimes Reinforce Physiological Stress and Support the Emergence of Diseases. Front. Environ. Sci. 2020, 8, 59. [Google Scholar] [CrossRef]
- Delghandi, M.R.; Menanteau-Ledouble, S.; Waldner, K.; El-Matbouli, M. Renibacterium salmoninarum and Mycobacterium spp.: Two Bacterial Pathogens Present at Low Levels in Wild Brown Trout (Salmo trutta fario) Populations in Austrian Rivers. BMC Vet. Res. 2020, 16, 40. [Google Scholar] [CrossRef] [Green Version]
- Sudhagar, A.; Kumar, G.; El-Matbouli, M. The Malacosporean Myxozoan Parasite Tetracapsuloides bryosalmonae: A Threat to Wild Salmonids. Pathogens 2020, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Hedrick, R.P.; MacConnell, E.; de Kinkelin, P. Proliferative Kidney Disease of Salmonid Fish. Annu. Rev. Fish Dis. 1993, 3, 277–290. [Google Scholar] [CrossRef]
- Bailey, C.; Segner, H.; Casanova-Nakayama, A.; Wahli, T. Who Needs the Hotspot? The Effect of Temperature on the Fish Host Immune Response to Tetracapsuloides bryosalmonae the Causative Agent of Proliferative Kidney Disease. Fish Shellfish Immunol. 2017, 63, 424–437. [Google Scholar] [CrossRef] [Green Version]
- Duval, E.; Quéméré, E.; Loot, G.; Jacquin, L.; Veyssière, C.; Blanchet, S. A Multifaceted Index of Population Health to Detect Risk-Prone Populations and Underlying Stressors in Wildlife. Biol. Conserv. 2022, 274, 109706. [Google Scholar] [CrossRef]
- Altermatt, F. Diversity in Riverine Metacommunities: A Network Perspective. Aquat Ecol 2013, 47, 365–377. [Google Scholar] [CrossRef] [Green Version]
- Saint-Pé, K.; Leitwein, M.; Tissot, L.; Poulet, N.; Guinand, B.; Berrebi, P.; Marselli, G.; Lascaux, J.-M.; Gagnaire, P.-A.; Blanchet, S. Development of a Large SNPs Resource and a Low-Density SNP Array for Brown Trout (Salmo trutta) Population Genetics. BMC Genom. 2019, 20, 582. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F.; Debes, P.V.; Palomar, G.; Vasemägi, A. Association Mapping Reveals Candidate Loci for Resistance and Anaemic Response to an Emerging Temperature-Driven Parasitic Disease in a Wild Salmonid Fish. Mol. Ecol. 2018, 27, 1385–1401. [Google Scholar] [CrossRef]
- Piertney, S.B.; Oliver, M.K. The Evolutionary Ecology of the Major Histocompatibility Complex. Heredity 2006, 96, 7–21. [Google Scholar] [CrossRef] [Green Version]
- Klein, J. Natural History of the Major Histocompatibility Complex; Wiley: Hoboken, NJ, USA, 1986; ISBN 978-0-471-80953-1. [Google Scholar]
- Portanier, E.; Garel, M.; Devillard, S.; Maillard, D.; Poissant, J.; Galan, M.; Benabed, S.; Poirel, M.-T.; Duhayer, J.; Itty, C.; et al. Both Candidate Gene and Neutral Genetic Diversity Correlate with Parasite Resistance in Female Mediterranean Mouflon. BMC Ecol. 2019, 19, 12. [Google Scholar] [CrossRef] [Green Version]
- Langefors, Å.; Lohm, J.; Von Schantz, T.; Grahn, M. Screening of Mhc Variation in Atlantic Salmon (Salmo salar): A Comparison of Restriction Fragment Length Polymorphism (RFLP), Denaturing Gradient Gel Electrophoresis (DGGE) and Sequencing. Mol. Ecol. 2000, 9, 215–219. [Google Scholar] [CrossRef]
- Olsén, K.H.; Grahn, M.; Lohm, J.; Langefors, Å. MHC and Kin Discrimination in Juvenile Arctic Charr, Salvelinus alpinus (L.). Anim. Behav. 1998, 56, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Belkhir, K.; Borsa, P.; Chikhi, L.; Raufaste, N.; Bonhomme, F. 1996–2004 GENETIX 4.05, Logiciel Sous Windows TM Pour La Genetique Des Populations. 2004. Available online: http://www.univmontp2.fr/~genetix/genetix/genetix.htm (accessed on 10 April 2023).
- Oosterhout, C.V.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-Checker: Software for Identifying and Correcting Genotyping Errors in Microsatellite Data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Goudet, J. Fstat (Ver. 2.9.4), a Program to Estimate and Test Population Genetics Parameters. Updated from Goudet (1995). 2003. Available online: http://www.unil.ch/izea/softwares/fstat.html (accessed on 10 April 2023).
- Rousset, F. Genepop’007: A Complete Re-Implementation of the Genepop Software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Coleman, R.A.; Weeks, A.R.; Hoffmann, A.A. Balancing Genetic Uniqueness and Genetic Variation in Determining Conservation and Translocation Strategies: A Comprehensive Case Study of Threatened Dwarf Galaxias, Galaxiella Pusilla (Mack) (Pisces: Galaxiidae). Mol. Ecol. 2013, 22, 1820–1835. [Google Scholar] [CrossRef]
- Verity, R.; Nichols, R.A. Estimating the Number of Subpopulations (K) in Structured Populations. Genetics 2016, 203, 1827–1839. [Google Scholar] [CrossRef] [Green Version]
- Burnham, K.P.; Anderson, D.R. Practical Use of the Information-Theoretic Approach. In Model Selection and Inference: A Practical Information-Theoretic Approach; Burnham, K.P., Anderson, D.R., Eds.; Springer: New York, NY, USA, 2002; pp. 75–117. ISBN 978-1-4757-2917-7. [Google Scholar]
- Barton, K. MuMIn: Multi-Model Inference. 2019. Available online: http://r-forge.r-project.org/projects/mumin/ (accessed on 10 April 2023).
- Grueber, C.E.; Nakagawa, S.; Laws, R.J.; Jamieson, I.G. Multimodel Inference in Ecology and Evolution: Challenges and Solutions. J. Evol. Biol. 2011, 24, 699–711. [Google Scholar] [CrossRef]
- Best, D.J.; Roberts, D.E. Algorithm AS 89: The Upper Tail Probabilities of Spearman’s Rho. J. R. Stat. Soc. Ser. C (Appl. Stat.) 1975, 24, 377–379. [Google Scholar] [CrossRef]
- Rousset, F. Genetic Differentiation and Estimation of Gene Flow from F-Statistics under Isolation by Distance. Genetics 1997, 145, 1219–1228. [Google Scholar] [CrossRef]
- Mantel, N. The Detection of Disease Clustering and a Generalized Regression Approach. Cancer Res. 1967, 27, 209–220. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 978-0-444-53869-7. [Google Scholar]
- Raeymaekers, J.A.M.; Van Houdt, J.K.J.; Larmuseau, M.H.D.; Geldof, S.; Volckaert, F.A.M. Divergent Selection as Revealed by PST and QTL-Based FST in Three-Spined Stickleback (Gasterosteus aculeatus) Populations along a Coastal-Inland Gradient. Mol. Ecol. 2007, 16, 891–905. [Google Scholar] [CrossRef]
- Tatarenkov, A.; Healey, C.I.M.; Avise, J.C. Microgeographic Population Structure of Green Swordail Fish: Genetic Differentiation despite Abundant Migration. Mol. Ecol. 2010, 19, 257–268. [Google Scholar] [CrossRef] [Green Version]
- Dowling, T.E.; Saltzgiver, M.J.; Marsh, P.C. Genetic Structure within and among Populations of the Endangered Razorback Sucker (Xyrauchen texanus) as Determined by Analysis of Microsatellites. Conserv Genet 2012, 13, 1073–1083. [Google Scholar] [CrossRef]
- Heggenes, J.; Røed, K.H. Do Dams Increase Genetic Diversity in Brown Trout (Salmo trutta)? Microgeographic Differentiation in a Fragmented River. Ecol. Freshw. Fish 2006, 15, 366–375. [Google Scholar] [CrossRef]
- Carrara, F.; Altermatt, F.; Rodriguez-Iturbe, I.; Rinaldo, A. Dendritic Connectivity Controls Biodiversity Patterns in Experimental Metacommunities. PNAS 2012, 109, 5761–5766. [Google Scholar] [CrossRef] [Green Version]
- Frantz, A.C.; Cellina, S.; Krier, A.; Schley, L.; Burke, T. Using Spatial Bayesian Methods to Determine the Genetic Structure of a Continuously Distributed Population: Clusters or Isolation by Distance? J. Appl. Ecol. 2009, 46, 493–505. [Google Scholar] [CrossRef]
- Perez, M.F.; Franco, F.F.; Bombonato, J.R.; Bonatelli, I.A.S.; Khan, G.; Romeiro-Brito, M.; Fegies, A.C.; Ribeiro, P.M.; Silva, G.A.R.; Moraes, E.M. Assessing Population Structure in the Face of Isolation by Distance: Are We Neglecting the Problem? Divers. Distrib. 2018, 24, 1883–1889. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | River | Site | Distance from the Confluence (km) | Centrality | N | |
|---|---|---|---|---|---|---|
| ARABas | L’Arac | Bas des gorges | 117.8 | −0.518 | 20 | |
| ARIPam | L’Ariège | Pamiers | 63.3 | 2.354 | 22 | |
| ARISav | L’Ariège | Savignac | 132.1 | 0.035 | 16 | |
| ARIVar | L’Ariège | Varilhes | 76.8 | 2.163 | 20 | |
| ARZDur | L’Arize | Durban | 95.8 | −0.518 | 25 | |
| BOUArg | La Bouigane | Argein | 119.9 | −0.615 | 20 | |
| COUPis | La Courbière | Aval pisciculturé | 105.8 | −0.711 | 20 | |
| HERPey | L’Hers | Peyrat | 134.3 | −0.330 | 20 | |
| LEZAub | Le Lez | Aubert | 107.8 | −0.145 | 20 | |
| ORIAx | L’Oriège | Ax-les-Thermes | 135.4 | −0.615 | 22 | |
| SALGir | Le Salat | St-Girons | 104.8 | 0.124 | 20 | |
| SALTau | Le Salat | Taurignan-Castet | 97.9 | 1.335 | 13 | |
| SAUAri | Le Saurat | Arignac | 106.2 | −0.810 | 20 | |
| SIOTra | Le Sios | Tramezaïgues | 92.4 | −0.712 | 20 | |
| TOULar | Le Touyre | Laroque-d’Olmes | 127.7 | −0.615 | 20 | |
| VICNia | Vicdessos | Niaux | 110.4 | −0.424 | 20 | |
| Total | 16 | 318 |
| SNPs | Candidate SNPs | MHC ClassIIβ | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Population | Na | Ar | He | Ho | FIS | FST | Na | Ar | He | Ho | FIS | FST | Na | Ar | He | Ho | FIS | FST |
| ARABas | 1.51 | 1.48 | 0.16 | 0.16 | 0.027 | 0.067 | 1.47 | 1.47 | 0.16 | 0.17 | −0.007 | 0.070 | 19 | 14.56 | 0.93 | 0.70 | 0.253 ** | 0.060 |
| ARIPam | 1.69 | 1.63 | 0.21 | 0.22 | −0.037 | 0.063 | 1.68 | 1.62 | 0.19 | 0.19 | 0.021 | 0.069 | 16 | 12.27 | 0.91 | 0.86 | 0.053 | 0.076 |
| ARISav | 1.72 | 1.67 | 0.17 | 0.18 | −0.015 | 0.114 | 1.63 | 1.60 | 0.17 | 0.16 | 0.076 | 0.060 | 12 | 11.00 | 0.90 | 1.00 | −0.111 | 0.088 |
| ARIVar | 1.69 | 1.62 | 0.19 | 0.20 | −0.034 | 0.062 | 1.68 | 1.63 | 0.18 | 0.17 | 0.100 | 0.039 | 13 | 11.38 | 0.91 | 0.84 | 0.077 | 0.075 |
| ARZDur | 1.50 | 1.45 | 0.14 | 0.15 | −0.013 | 0.130 | 1.63 | 1.54 | 0.16 | 0.16 | 0.045 | 0.132 | 10 | 8.44 | 0.84 | 0.83 | 0.013 | 0.126 |
| BOUArg | 1.50 | 1.48 | 0.17 | 0.17 | −0.005 | 0.081 | 1.53 | 1.46 | 0.14 | 0.14 | 0.046 | 0.072 | 8 | 6.71 | 0.70 | 0.60 | 0.146 | 0.180 |
| COUPis | 1.60 | 1.57 | 0.19 | 0.19 | 0.042. | 0.066 | 1.58 | 1.56 | 0.20 | 0.18 | 0.105 | 0.085 | 18 | 14.79 | 0.95 | 0.90 | 0.051 | 0.068 |
| HERPey | 1.52 | 1.49 | 0.15 | 0.14 | 0.082 ** | 0.116 | 1.53 | 1.48 | 0.13 | 0.14 | −0.005 | 0.061 | 11 | 9.51 | 0.87 | 0.50 | 0.430 *** | 0.098 |
| LEZAub | 1.52 | 1.50 | 0.18 | 0.17 | 0.047 * | 0.064 | 1.47 | 1.45 | 0.16 | 0.16 | 0.007 | 0.055 | 17 | 13.70 | 0.91 | 0.50 | 0.459 *** | 0.071 |
| ORIAx | 1.72 | 1.64 | 0.17 | 0.17 | 0.013 | 0.114 | 1.68 | 1.61 | 0.18 | 0.19 | −0.034 | 0.070 | 11 | 8.43 | 0.79 | 0.76 | 0.036 | 0.141 |
| SALGir | 1.52 | 1.50 | 0.17 | 0.17 | −0.006 | 0.137 | 1.58 | 1.53 | 0.17 | 0.18 | −0.037 | 0.166 | 14 | 12.08 | 0.91 | 0.56 | 0.396 *** | 0.091 |
| SALTau | 1.48 | 1.47 | 0.17 | 0.17 | 0.078 * | 0.063 | 1.53 | 1.52 | 0.16 | 0.16 | 0.036 | 0.042 | 10 | 10.00 | 0.86 | 0.54 | 0.380 ** | 0.070 |
| SAUAri | 1.62 | 1.56 | 0.18 | 0.17 | 0.092 *** | 0.060 | 1.68 | 1.60 | 0.15 | 0.15 | 0.037 | 0.040 | 19 | 14.09 | 0.90 | 1.00 | −0.114 | 0.094 |
| SIOTra | 1.66 | 1.61 | 0.19 | 0.19 | 0.047 * | 0.083 | 1.58 | 1.54 | 0.17 | 0.18 | −0.065 | 0.082 | 18 | 13.90 | 0.91 | 0.85 | 0.072 | 0.078 |
| TOULar | 1.56 | 1.53 | 0.17 | 0.17 | 0.040 | 0.060 | 1.53 | 1.51 | 0.16 | 0.16 | 0.067 | 0.052 | 15 | 12.74 | 0.92 | 0.95 | −0.028 | 0.078 |
| VICNia | 1.52 | 1.49 | 0.16 | 0.16 | 0.050 * | 0.078 | 1.53 | 1.51 | 0.17 | 0.19 | −0.055 | 0.054 | 15 | 12.90 | 0.93 | 0.95 | −0.023 | 0.070 |
| df | Loglik. | AICc | ΔAICc | Weight | |
|---|---|---|---|---|---|
| Genetic diversity (Ar) | |||||
| c + c2 | 4 | −62.957 | 134.8 | 0.00 | 0.246 |
| d + d2 | 4 | −63.368 | 135.7 | 0.82 | 0.163 |
| d | 3 | −64.979 | 136.5 | 1.66 | 0.107 |
| Genetic differentiation (FST) | |||||
| c + c2 + d + d2 | 6 | −60.599 | 135.2 | 0.00 | 0.319 |
| c | 3 | −65.047 | 136.6 | 1.39 | 0.159 |
| Estimate ± Se | CI | RI | |
|---|---|---|---|
| Genetic diversity (Ar) | |||
| Centrality | −0.41 ± 0.32 | −1.05, 0.23 | 0.48 |
| Centrality2 | 0.42 ± 0.19 | 0.03, 0.81 | 0.48 |
| Distance from confluence | −0.18 ± 0.15 | −0.49, 0.12 | 0.52 |
| Distance from confluence2 | 0.20 ± 0.11 | −0.03, 0.42 | 0.32 |
| Genetic differentiation (FST) | |||
| Centrality | 0.08 ± 0.35 | −0.62, 0.79 | 1 |
| Centrality2 | −0.90 ± 0.32 | −1.54, −0.26 | 0.67 |
| Distance from confluence | −0.51 ± 0.26 | −1.04, 0.02 | 0.67 |
| Distance from confluence2 | 0.53 ± 0.20 | 0.12, 0.94 | 0.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gouthier, L.; Duval, E.; Blanchet, S.; Loot, G.; Veyssière, C.; Galan, M.; Quéméré, E.; Jacquin, L. Spatial Patterns of Neutral and Functional Genetic Variations along Dendritic Networks of Riverscape in Brown Trout Populations. Diversity 2023, 15, 784. https://0-doi-org.brum.beds.ac.uk/10.3390/d15060784
Gouthier L, Duval E, Blanchet S, Loot G, Veyssière C, Galan M, Quéméré E, Jacquin L. Spatial Patterns of Neutral and Functional Genetic Variations along Dendritic Networks of Riverscape in Brown Trout Populations. Diversity. 2023; 15(6):784. https://0-doi-org.brum.beds.ac.uk/10.3390/d15060784
Chicago/Turabian StyleGouthier, Laurine, Eloïse Duval, Simon Blanchet, Géraldine Loot, Charlotte Veyssière, Maxime Galan, Erwan Quéméré, and Lisa Jacquin. 2023. "Spatial Patterns of Neutral and Functional Genetic Variations along Dendritic Networks of Riverscape in Brown Trout Populations" Diversity 15, no. 6: 784. https://0-doi-org.brum.beds.ac.uk/10.3390/d15060784