
Golden Eagle Populations, Movements, and Landscape Barriers: Insights from Scotland

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
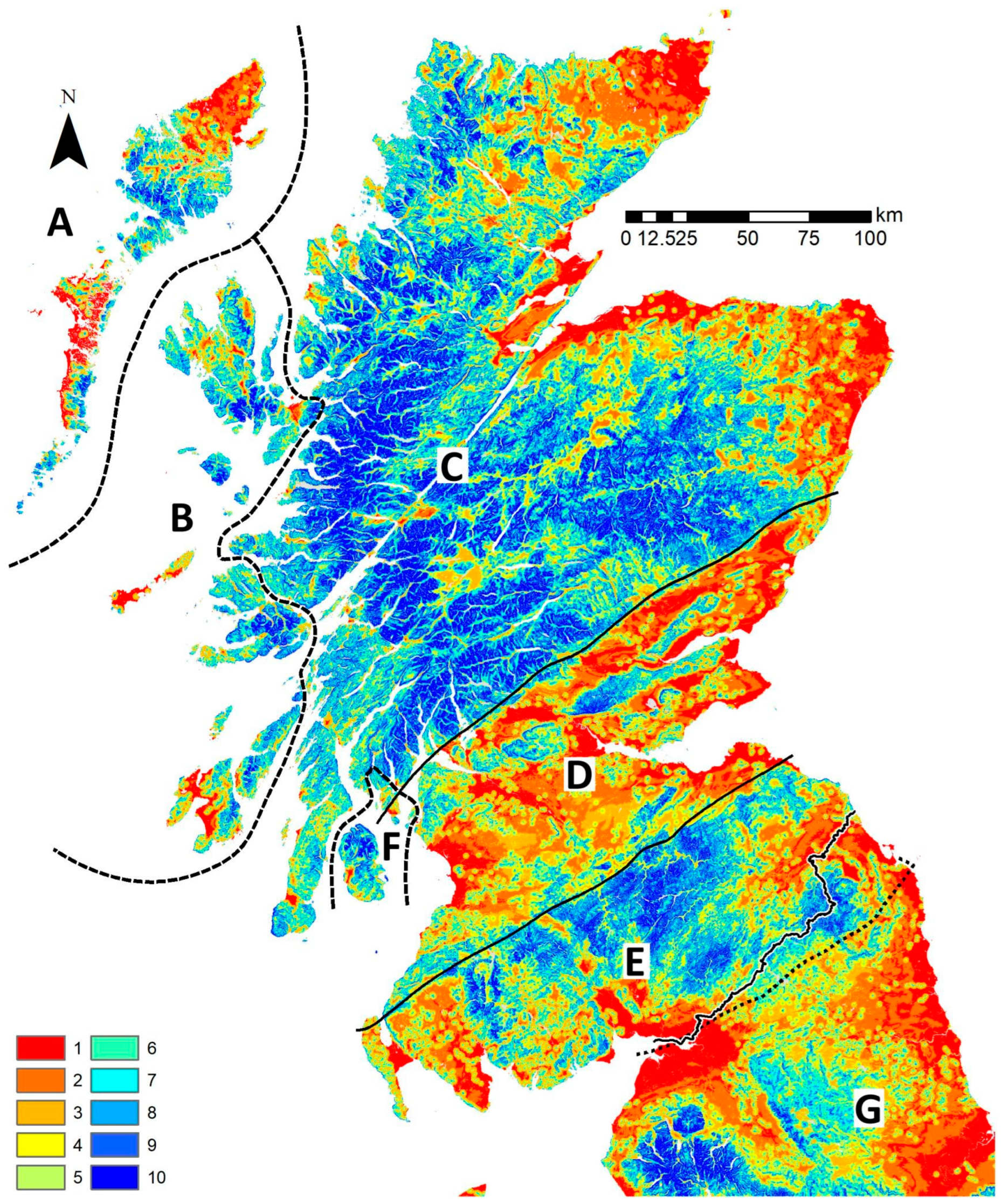
2.1. Study Area
2.2. Study Species and GET Predictions
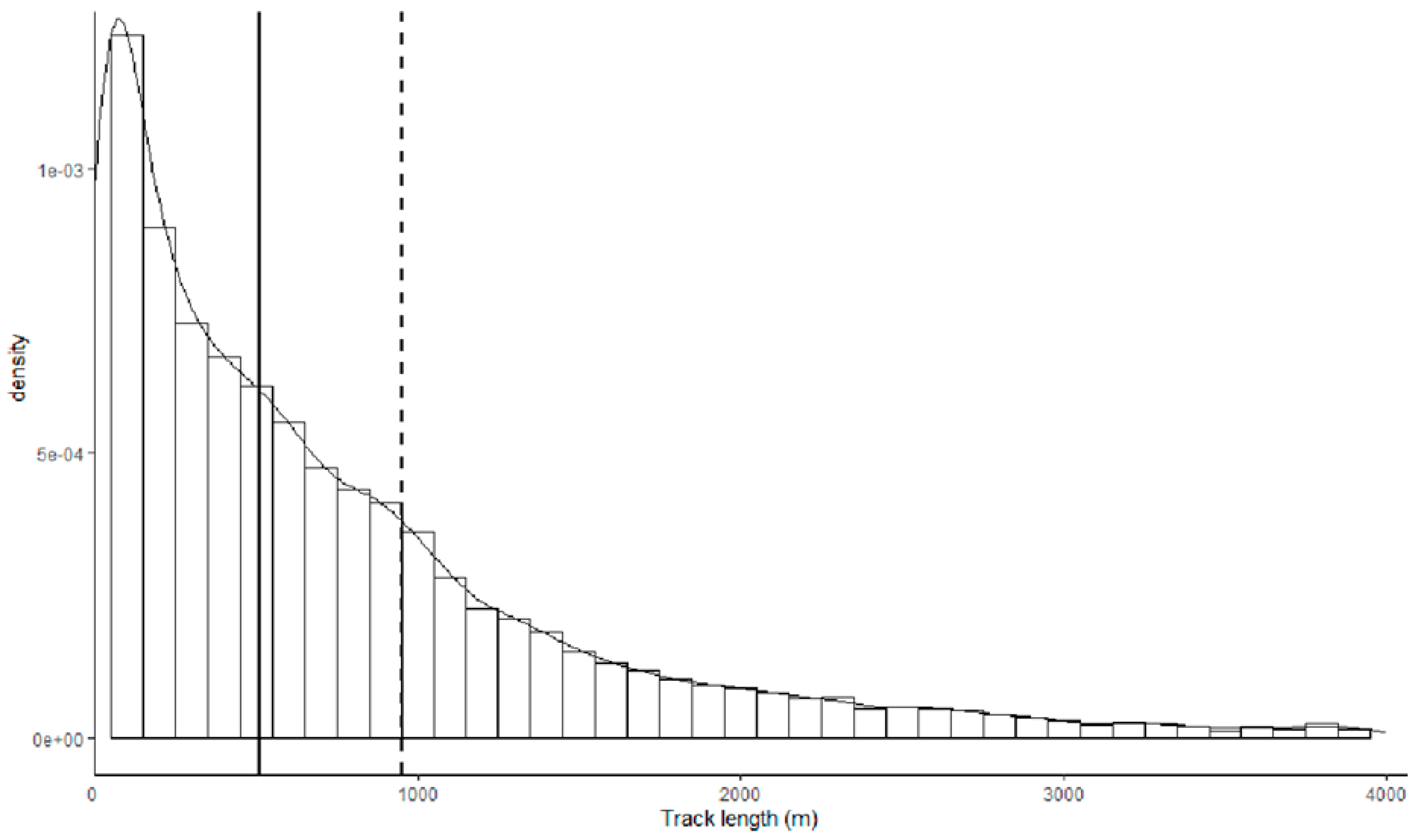
2.3. GPS Tagging and Data Treatments
3. Results
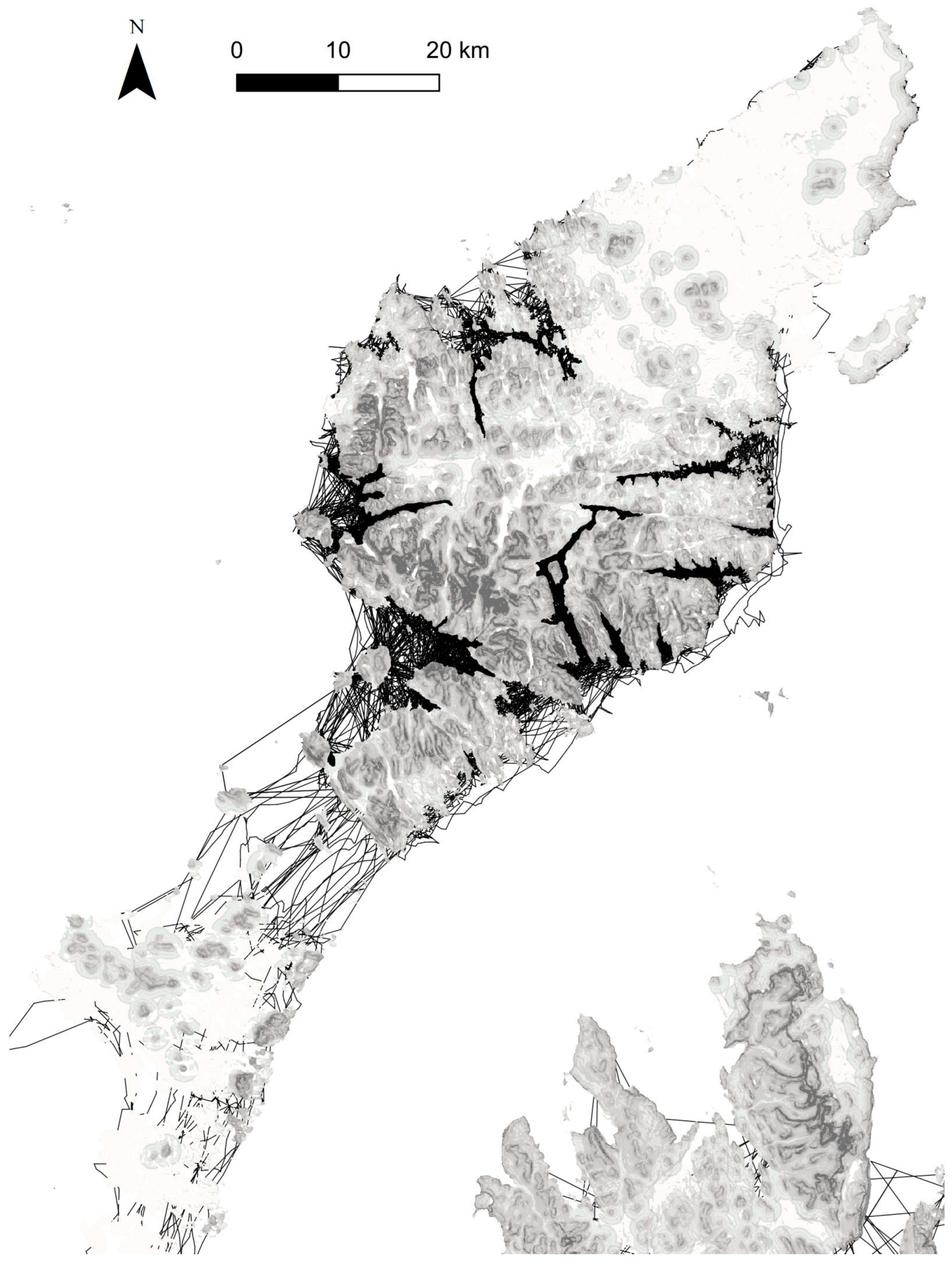
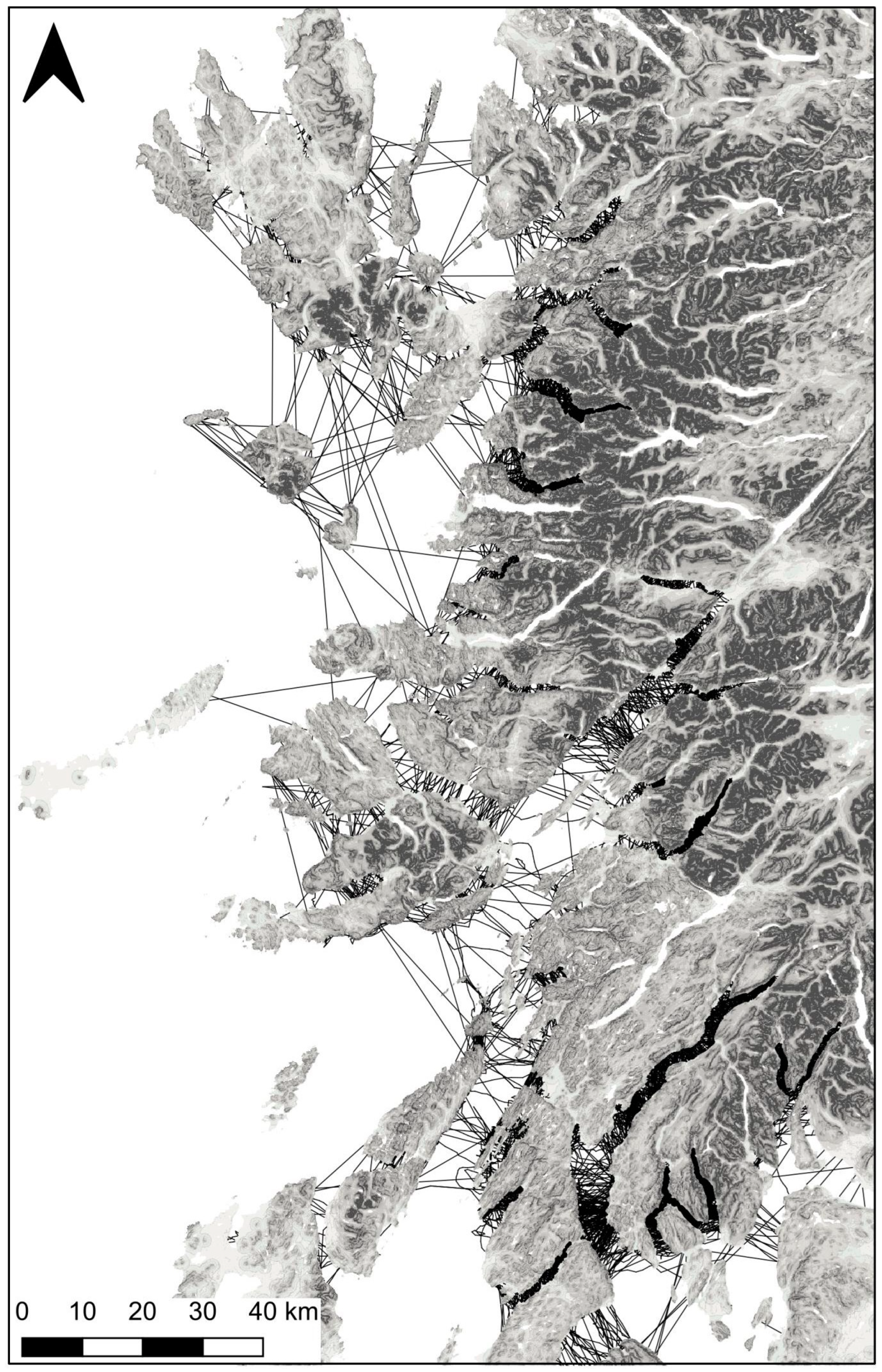
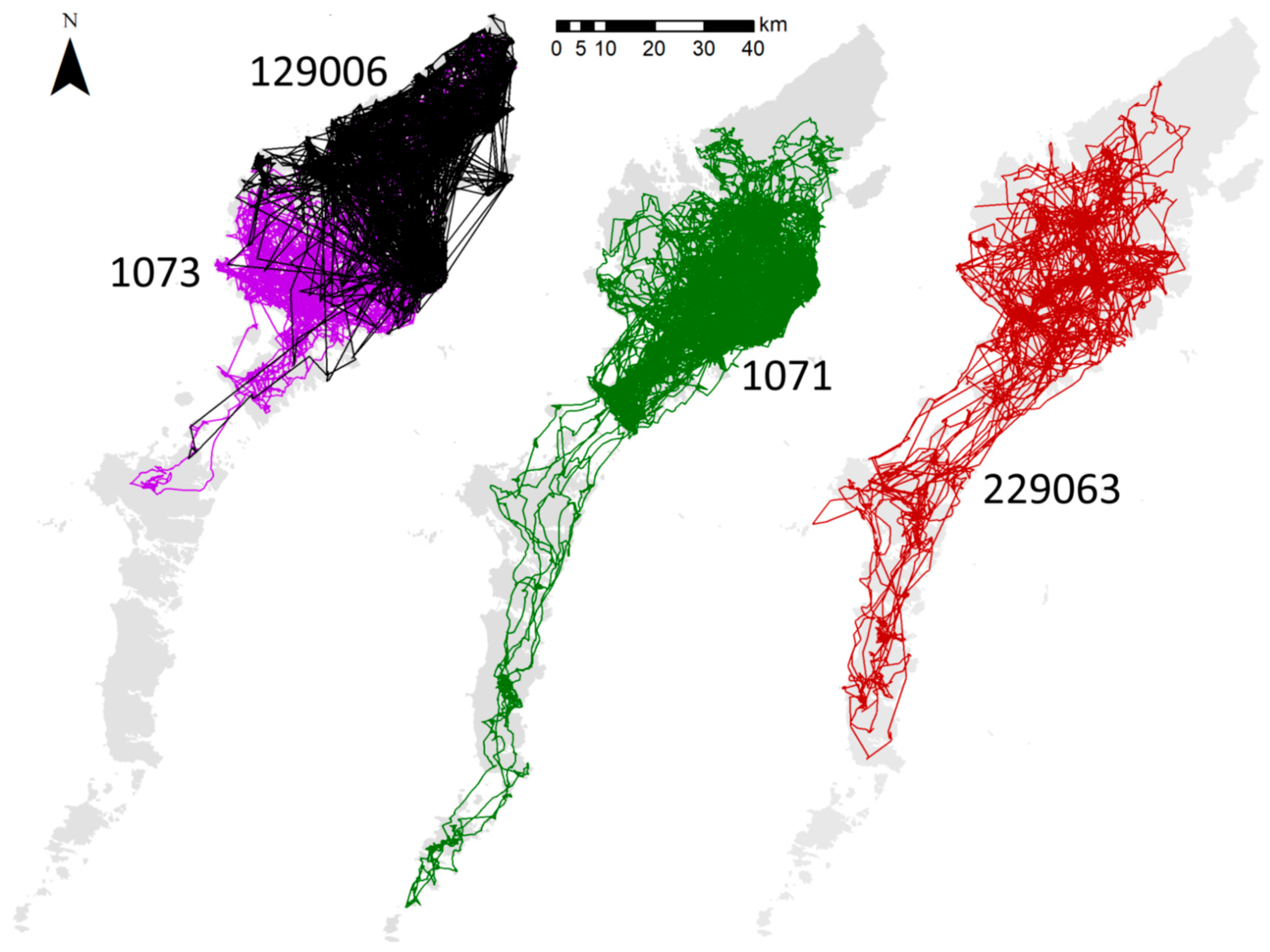
3.1. Hypothesis 1, a Terrestrial Barrier
3.2. Hypothesis 2, Limits to Crossing Seawater
3.3. Hypothesis 3, the Isolation of Southern Scotland
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Details of apparently aborted attempts by three birds tagged on the Outer Hebrides to cross to the Highlands mainland or the Inner Hebrides
- Movements within the Outer Hebrides: few tagged birds went south from the northern islands of Lewis and Harris

- Additional information from ringing in the Outer Hebrides
- Patagial wing-tag record of nestling translocated to Ireland later moving back to Scotland
References
- López-López, P. Individual-based tracking systems in ornithology: Welcome to the era of big data. Ardeola 2016, 63, 103–136. [Google Scholar] [CrossRef]
- Weston, E.D.; Whitfield, D.P.; Travis, J.M.; Lambin, X. When do young birds disperse? Tests from studies of golden eagles in Scotland. BMC Ecol. 2013, 13, 42. [Google Scholar] [CrossRef]
- Fielding, A.H.; Haworth, P.; Anderson, D.; Benn, S.; Dennis, R.; Weston, E.; Whitfield, D.P. A simple topographical model to predict Golden Eagle Aquila chrysaetos space use during dispersal. Ibis 2020, 162, 400–415. [Google Scholar] [CrossRef]
- Fielding, A.H.; Anderson, D.; Benn, S.; Reid, R.; Tingay, R.; Weston, E.D.; Whitfield, D.P. Substantial Variation in Prospecting Behaviour of Young Golden Eagles Aquila chrysaetos Defies Expectations from Potential Predictors. Diversity 2023, 15, 506. [Google Scholar] [CrossRef]
- Whitfield, D.P.; Fielding, A.H.; Anderson, D.; Benn, S.; Dennis, R.; Grant, J.; Weston, E.D. Age of First Territory Settlement of Golden Eagles Aquila chrysaetos in a Variable Competitive Landscape. Front. Ecol. Evol. 2022, 10, 743598. [Google Scholar] [CrossRef]
- Whitfield, D.P.; Fielding, A.H.; Anderson, D.; Benn, S.; Reid, R.; Tingay, R.; Weston, E.D. Sex difference in natal dispersal distances of Golden Eagles Aquila chrysaetos in Scotland. Ibis 2024, 166, 146–155. [Google Scholar] [CrossRef]
- Whitfield, D.P.; Fielding, A.H.; Anderson, D.; Benn, S.; Reid, R.; Tingay, R.; Weston, E.D. Seasonal Variation in First Territory Settlement of Dispersing Golden Eagles: An Innate Behaviour? Diversity 2024, 16, 82. [Google Scholar] [CrossRef]
- Ogden, R.E.; Heap, E.; McEwing, R.; Tingay, R.; Whitfield, D.P. Population structure and dispersal history in Scottish Golden eagles Aquila chrysaetos revealed by molecular genetic analysis of territorial birds. Ibis 2015, 157, 834–848. [Google Scholar] [CrossRef]
- Nebel, C.; Gamauf, A.; Haring, E.; Segelbacher, G.; Villers, A.; Zachos, F.E. Mitochondrial DNA analysis reveals Holarctic homogeneity and a distinct Mediterranean lineage in the golden eagle (Aquila chrysaetos). Biol. J. Linn. Soc. 2015, 116, 328–340. [Google Scholar] [CrossRef]
- Nebel, C.; Gamauf, A.; Haring, E.; Segelbacher, G.; Väli, Ű.; Zachos, F.E. New insights into population structure of the European golden eagle (Aquila chrysaetos) revealed by microsatellite analysis. Biol. J. Linn. Soc. 2019, 128, 611–631. [Google Scholar] [CrossRef]
- Sato, Y.; Ogden, R.; Kishida, T.; Nakajima, N.; Maeda, T.; Inoue-Murayama, M. Population history of the golden eagle inferred from whole-genome sequencing of three of its subspecies. Biol. J. Linn. Soc. 2020, 120, 826–838. [Google Scholar] [CrossRef]
- Masuda, R.; Noro, M.; Kurose, N.; Nishida-Umehara, C.; Takechi, H.; Yamazaki, T.; Kosuge, M.; Yoshida, M.C. Genetic characteristics of endangered Japanese golden eagles (Aquila chrysaetos japonica) based on mitochondrial DNA D-loop sequences and karyotypes. Zoo Biol. 1998, 17, 111–121. [Google Scholar] [CrossRef]
- Sato, Y.; Humble, E.; Ogden, R. Genomic data reveal strong differentiation and reduced genetic diversity in island golden eagle populations. Biol. J. Linn. Soc. 2023, blad172. [Google Scholar] [CrossRef]
- Wink, M.; Sauer-Gürth, H. Phylogenetic relationships in diurnal raptors based on nucleotide sequences of mitochondrial and nuclear marker genes. In Raptors Worldwide: Proceedings of the VI World Conference on Birds of Prey and Owls, Budapest, Hungary, 18–23 May 2003; Chancellor, R.D., Meyburg, B.-U., Eds.; Working Group on Birds of Prey/MME-BirdLife: Budapest, Hungary, 2004; pp. 483–495. [Google Scholar]
- Judkins, M.E.; Van den Bussche, R.A. Holarctic phylogeography of golden eagles (Aquila chrysaetos) and evaluation of alternative North American management approaches. Biol. J. Linn. Soc. 2018, 123, 471–482. [Google Scholar] [CrossRef]
- Watson, J. The Golden Eagle, 2nd ed.; Poyser: London, UK, 2010. [Google Scholar]
- Brown, J.L.; Bedrosian, B.; Bell, D.A.; Braham, M.A.; Cooper, J.; Crandall, R.H.; DiDonato, J.; Domenech, R.; Duerr, A.E.; Katzner, T.E.; et al. Patterns of spatial distribution of Golden Eagles across North America: How do they fit into existing landscape-mapping systems? J. Raptor Res. 2017, 51, 197–215. [Google Scholar] [CrossRef]
- Miller, T.A.; Lanzone, M.J.; Braham, M.A.; Duerr, A.E.; Cooper, J.; Somershoe, S.; Hanni, D.; Soehren, E.C.; Threadgill, C.; Maddox, M.; et al. Winter distribution of Golden Eagles in the eastern USA. J. Raptor Res. 2023, 57, 522–532. [Google Scholar] [CrossRef]
- Murphy, R.K.; Millsap, B.A.; Stahlecker, D.W.; Boal, C.W.; Smith, B.W.; Mullican, S.E.; Borgman, C.C. Ectoparasitism and energy infrastructure limit survival of preadult Golden Eagles in the Southern Great Plains. J. Raptor Res. 2023, 57, 505–521. [Google Scholar] [CrossRef]
- Waples, R.S.; Gaggiotti, O. What is a population? An empirical evaluation of some genetic methods for identifying the number of gene pools and their degree of connectivity. Mol. Ecol. 2006, 15, 1419–1439. [Google Scholar] [CrossRef] [PubMed]
- Lowe, W.H.; Allendorf, F.W. What can genetics tell us about population connectivity? Mol. Ecol. 2010, 19, 3038–3051. [Google Scholar] [CrossRef]
- Katzner, T.; Smith, B.W.; Miller, T.A.; Brandes, D.; Cooper, J.; Lanzone, M.; Brauning, D.; Farmer, C.; Harding, S.; Kramar, D.E.; et al. Status, biology, and conservation priorities for North America’s eastern Golden Eagle (Aquila chrysaetos) population. Auk 2012, 129, 168–176. [Google Scholar]
- Millsap, B.A.; Harmata, A.R.; Stahlecker, D.W.; Mikesic, D.G. Natal dispersal distance of bald and golden eagles originating in the coterminous United States as inferred from band encounters. J. Raptor Res. 2014, 48, 13–23. [Google Scholar] [CrossRef]
- Muriel, R.; Morandini, V.; Ferrer, M.; Balbontín, J. Independence and juvenile dispersal distances in wild and reintroduced Spanish imperial eagles. Biol. Conserv. 2015, 191, 300–305. [Google Scholar] [CrossRef]
- Whitfield, D.P.; Duffy, K.; McLeod, D.R.; Evans, R.J.; MacLennan, A.M.; Reid, R.; Sexton, D.; Wilson, J.D.; Douse, A. Juvenile dispersal of white-tailed eagles in western Scotland. J. Raptor Res. 2009, 43, 110–120. [Google Scholar] [CrossRef]
- Hayhow, D.B.; Benn, S.; Stevenson, A.; Stirling-Aird, P.; Eaton, M. Status of Golden Eagle Aquila chrysaetos in Britain in 2015. Bird Study 2017, 64, 281–294. [Google Scholar] [CrossRef]
- O’Toole, L.; Fielding, A.H.; Haworth, P.F. Re-introduction of the Golden Eagle Aquila chrysaetos into the Republic of Ireland. Biol. Conserv. 2002, 103, 303–312. [Google Scholar] [CrossRef]
- Evans, R.J.; O’Toole, L.; Whitfield, D.P. The history of eagles in Britain and Ireland: An ecological review of placename and documentary evidence from the last 1500 years. Bird Study 2012, 59, 335–349. [Google Scholar] [CrossRef]
- Fielding, A.H.; Anderson, D.; Benn, S.; Taylor, J.; Tingay, R.; Weston, E.D.; Whitfield, D.P. Responses of GPS-Tagged Territorial Golden Eagles Aquila chrysaetos to Wind Turbines in Scotland. Diversity 2023, 15, 917. [Google Scholar] [CrossRef]
- Whitfield, D.P.; Fielding, A.H. Analyses of the Fates of Satellite Tracked Golden Eagles in Scotland; Scottish Natural Heritage Commissioned Report, No. 982; SNH: Scotland, UK, 2017. [Google Scholar]
- Gillen, C. Geology and Landscapes of Scotland; Terra: Harpenden, UK, 2003. [Google Scholar]
- Trewin, N.H. Geology of Scotland, 4th ed.; Geological Society: London, UK, 2002. [Google Scholar]
- Hunter, A.; Easterbrook, G. The Geological History of the British Isles; The Open University: Milton Keynes, UK, 2004. [Google Scholar]
- British Geological Survey. Bedrock Geology UK North, 1:625,000 Scale; British Geological Survey: Keyworth, Nottingham, UK, 2007. [Google Scholar]
- Bildstein, K.L. Migrating Raptors of the World: Their Ecology and Conservation; Cornell University Press: Ithaca, NY, USA, 2006. [Google Scholar]
- Newton, I. The Migration Ecology of Birds; Academic Press: London, UK, 2008. [Google Scholar]
- Pennycuick, C.J. Modelling the Flying Bird, 5th ed.; Elsevier: Oxford, UK, 2008. [Google Scholar]
- Diehl, R.H. The airspace is habitat. Trends Ecol. Evol. 2013, 28, 377–379. [Google Scholar] [CrossRef]
- Bildstein, K.L.; Bechard, M.J.; Farmer, C.; Newcomb, L. Narrow sea crossings present major obstacles to migrating Griffon Vultures Gyps fulvus. Ibis 2009, 151, 382–391. [Google Scholar] [CrossRef]
- Evans, R.J.; Reid, R.; Whitfield, P. Pre-First World War persistence of a Golden Eagle population in the Outer Hebrides. Scott. Birds 2013, 33, 34–36. [Google Scholar]
- Haswell-Smith, H. The Scottish Islands; Canongate: Edinburgh, UK, 2004. [Google Scholar]
- National Records of Scotland. Other National Records of Scotland (NRS) Geographies Datasets; National Records of Scotland: Edinburgh, UK, 2023. Available online: nrscotland.gov.uk (accessed on 14 December 2023).
- Dennis, R.H.; Ellis, P.M.; Broad, R.A.; Langslow, D.R. The status of the Golden Eagle in Britain. Br. Birds 1984, 77, 592–607. [Google Scholar]
- Green, R.E. The status of the Golden Eagle in Britain in 1992. Bird Study 1996, 43, 20–27. [Google Scholar] [CrossRef]
- Eaton, M.A.; Dillon, I.A.; Stirling-Aird, P.K.; Whitfield, D.P. Status of Golden Eagle Aquila chrysaetos in Britain in 2003. Bird Study 2007, 54, 212–220. [Google Scholar] [CrossRef]
- Whitfield, D.P.; Fielding, A.H.; McLeod, D.R.A.; Morton, K.; Stirling-Aird, P.; Eaton, M. Factors constraining the distribution of Golden Eagles Aquila chrysaetos in Scotland. Bird Study 2007, 54, 199–211. [Google Scholar] [CrossRef]
- Whitfield, D.P.; Fielding, A.H.; McLeod, D.R.A.; Haworth, P.F. A Conservation Framework for Golden Eagles: Implications for Their Conservation and Management in Scotland; Scottish Natural Heritage Commissioned Report No.193; SNH: Scotland, UK, 2008. [Google Scholar]
- Benn, S.; Whitfield, D.P. Golden Eagle Ecology and Conservation in the Central Highlands of Scotland (NHZ 10) in 2022; Natural Research: Banchory, Scotland, UK, 2023. [Google Scholar]
- Whitfield, D.P.; MacLeod, D.R.A.; Watson, J.; Fielding, A.H.; Haworth, P.F. The association of grouse moor in Scotland with the illegal use of poisons to control predators. Biol. Conserv. 2003, 114, 157–163. [Google Scholar] [CrossRef]
- Fielding, A.H.; Haworth, P.F. Golden Eagles in the South of Scotland: An Overview; Scottish Natural Heritage Commissioned Report No. 626; Scottish Natural Heritage: Battleby, Scotland, UK, 2014. [Google Scholar]
- Baxter, E.V.; Rintoul, L.J. The Birds of Scotland: Their History, Distribution and Migration; Oliver and Boyd: Edinburgh, Scotland, UK, 1953; Volume 1. [Google Scholar]
- Marquiss, M.; Ratcliffe, D.A.; Roxburgh, R. The numbers, breeding success and diet of golden eagles in southern Scotland in relation to changes in land-use. Biol. Conserv. 1985, 34, 121–140. [Google Scholar] [CrossRef]
- Ratcliffe, D.A. Galloway and the Borders; Collins New Naturalist: London, UK, 2007. [Google Scholar]
- Morandini, V.; de Benito, E.; Newton, I.; Ferrer, M. Natural expansion versus translocation in a previously human-persecuted bird of prey. Ecol. Evol. 2017, 7, 3682–3688. [Google Scholar] [CrossRef]
- Evans, R.J.; Wilson, J.D.; Amar, A.; Douse, A.; MacLennan, A.; Ratcliffe, N.; Whitfield, D.P. Growth and demography of a reintroduced population of white-tailed eagles Haliaeetus albicilla. Ibis 2009, 151, 244–254. [Google Scholar] [CrossRef]
- Ferrer, M.; Morandini, V.; Baguena, G.; Newton, I. Reintroducing endangered raptors: A case study of supplementary feeding and removal of nestlings from wild populations. J. Appl. Ecol. 2018, 55, 1360–1367. [Google Scholar] [CrossRef]
- Ratcliffe, D.A.; Thompson, D.B.A. The British uplands: Their ecological character and international significance. In Ecological Change in the Uplands; Usher, M.B., Thompson, D.B.A., Eds.; Blackwell Scientific Publications: Oxford, UK, 1988; pp. 9–36. [Google Scholar]
- Newton, I. Uplands and Birds; Collins: London, UK, 2020. [Google Scholar]
- Benn, S.; Fielding, A.H.; Whitfield, D.P. Golden Eagles in Scotland. In The Golden Eagle around the World: A Monograph on a Holarctic Raptor; Bautista, J., Ellis, D.H., Eds.; Hancock House Publishers: Surrey, BC, Canada, 2024; pp. 265–281. [Google Scholar]
- Ferrer, M. Juvenile dispersal behavior and natal philopatry of a long-lived raptor, the Spanish Imperial Eagle Aquila adalberti. Ibis 1993, 135, 132–138. [Google Scholar] [CrossRef]
- Clobert, J.; Le Galliard, J.F.; Cote, J.; Meylan, S.; Massot, M. Informed dispersal, heterogeneity in animal dispersal syndromes and the dynamics of spatially structured populations. Ecol. Lett. 2009, 12, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.E.; Holland, A.E.; Bryan, A.L.; Beasley, J.C. Environmental conditions and animal behavior influence performance of solar-powered GPS-GSM transmitters. Condor 2017, 119, 389–404. [Google Scholar] [CrossRef]
- Fielding, A.H.; Anderson, D.; Benn, S.; Dennis, R.; Geary, M.; Weston, E.; Whitfield, D.P. Responses of GPS-tagged Golden Eagles (Aquila chrysaetos) to multiple wind farms across Scotland. Ibis 2022, 164, 102–117. [Google Scholar] [CrossRef]
- Mee, A.; Breen, D.; Clarke, D.; Heardman, C.; Lyden, J.; McMahon, F.; O’Sullivan, P.; O’Toole, L. Reintroduction of white-tailed eagles Haliaeetus albicilla to Ireland. Irish Birds 2016, 10, 301–314. [Google Scholar]
- Whitfield, D.P.; Douse, A.; Evans, R.J.; Grant, J.; Love, J.; McLeod, D.R.A.; Reid, R.; Wilson, J.D. Natal and breeding dispersal in a reintroduced population of White-tailed Eagles Haliaeetus albicilla. Bird Study 2009, 56, 177–186. [Google Scholar] [CrossRef]
- Weston, E. Juvenile Dispersal Behaviour in the Golden Eagle (Aquila chrysaetos). Ph.D. Thesis, University of Aberdeen, Aberdeen, UK, 2014. [Google Scholar]
- Greenwood, P.J. Mating systems, philopatry and dispersal in birds and mammals. Anim. Behav. 1980, 28, 1140–1162. [Google Scholar] [CrossRef]
- Greenwood, P.; Harvey, P.H. The natal and breeding dispersal of birds. Annu. Rev. Ecol. Evol. Syst. 1982, 13, 1–21. [Google Scholar] [CrossRef]
- Newton, I. Population Ecology of Raptors; Poyser: Berkhamstead, UK, 1979. [Google Scholar]
- Struwe-Juhl, B.; Grünkorn, T. Results of colour-ringing White-tailed Sea Eagles Haliaeetus albicilla in Schleswig-Holstein: Site fidelity, movements, dispersal, age of first breeding, age structure and breeding of siblings. Vogelwelt 2007, 128, 117–129. [Google Scholar]
- Muriel, R.; Morandini, V.; Ferrer, M.; Balbontín, J.; Morlanes, V. Juvenile dispersal behaviour and conspecific attraction: An alternative approach with translocated Spanish imperial eagles. Anim. Behav. 2016, 116, 17–29. [Google Scholar] [CrossRef]
- Sansom, A.; Evans, R.; Roos, S. Population and Future Range Modelling of Reintroduced Scottish White-Tailed Eagles (Haliaeetus albicilla); Scottish Natural Heritage Commissioned Report No. 898; SNH: Scotland, UK, 2016. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fielding, A.H.; Anderson, D.; Barlow, C.; Benn, S.; Reid, R.; Tingay, R.; Weston, E.D.; Whitfield, D.P. Golden Eagle Populations, Movements, and Landscape Barriers: Insights from Scotland. Diversity 2024, 16, 195. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040195
Fielding AH, Anderson D, Barlow C, Benn S, Reid R, Tingay R, Weston ED, Whitfield DP. Golden Eagle Populations, Movements, and Landscape Barriers: Insights from Scotland. Diversity. 2024; 16(4):195. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040195
Chicago/Turabian StyleFielding, Alan H., David Anderson, Catherine Barlow, Stuart Benn, Robin Reid, Ruth Tingay, Ewan D. Weston, and D. Philip Whitfield. 2024. "Golden Eagle Populations, Movements, and Landscape Barriers: Insights from Scotland" Diversity 16, no. 4: 195. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040195