
Metarhizium dianzhongense sp. nov. and New Record of M. bibionidarum (Clavicipitaceae, Hyocreales) Attacking Insects from China
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation
2.2. Test Material
2.3. Morphological Observation
2.4. DNA Extraction, PCR Amplification, and Sequencing
2.5. Phylogenetic Analyses
2.6. Pathogenicity Test
3. Results
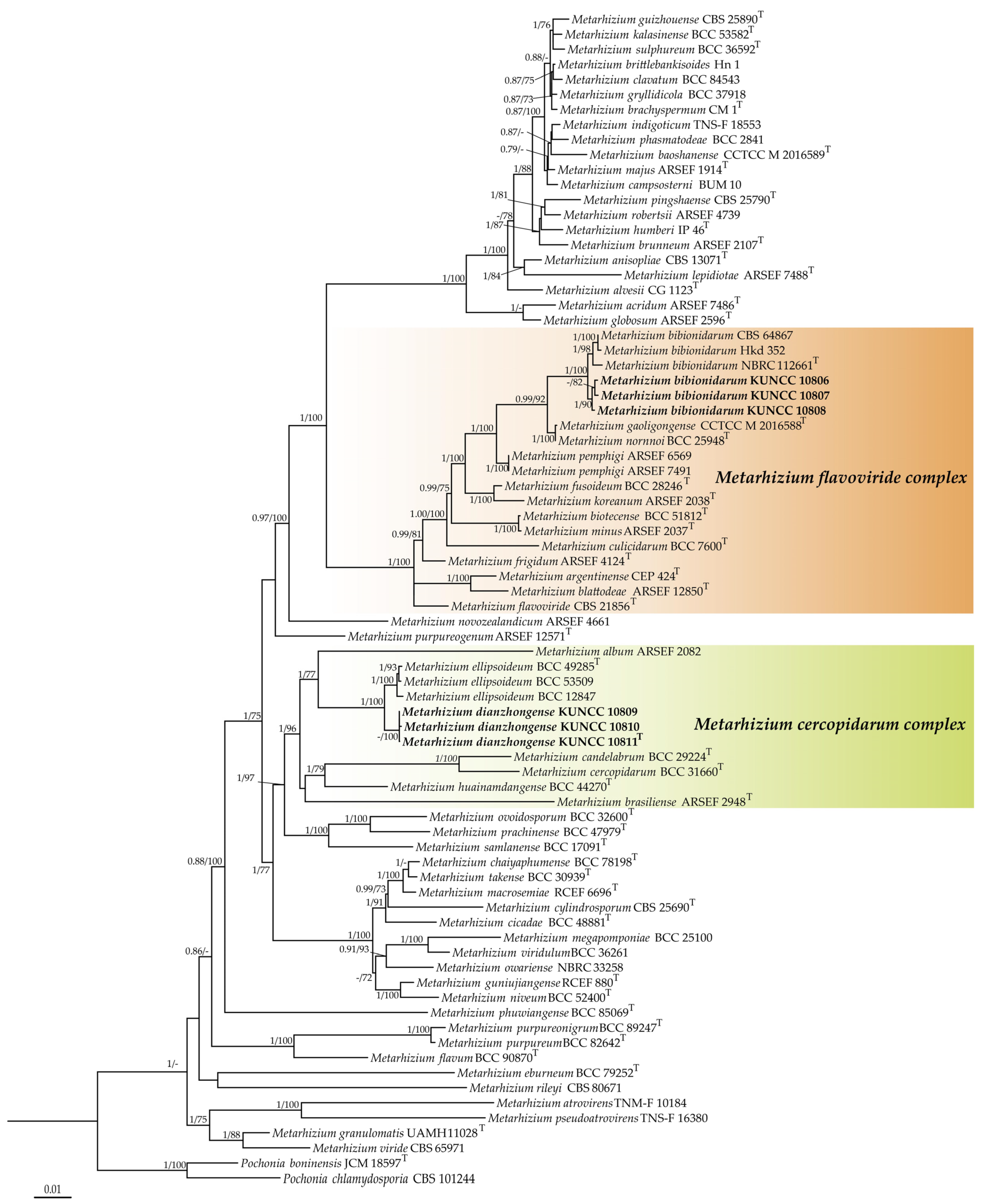
3.1. Phylogenetic Analyses
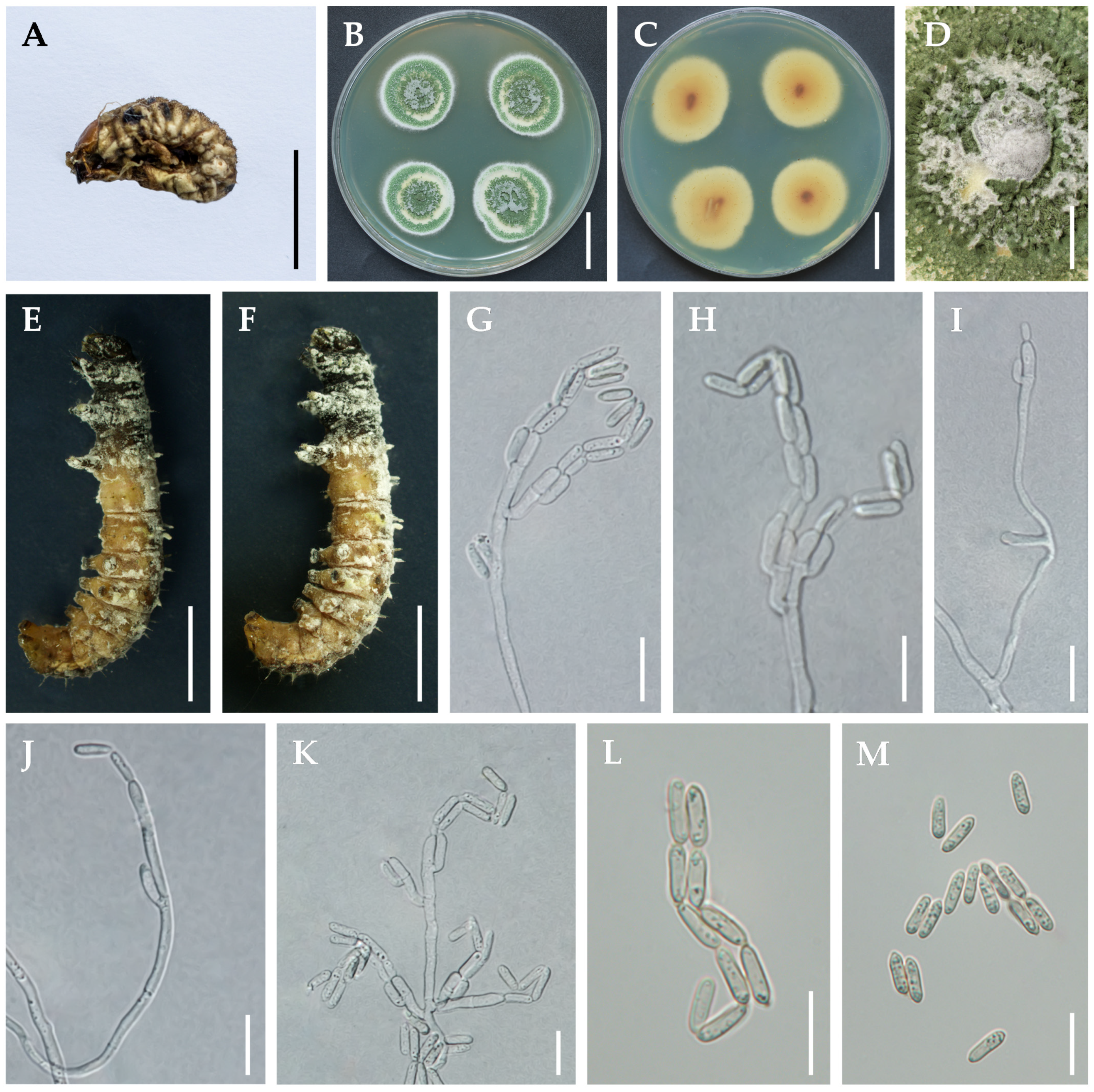
3.2. Taxonomy
3.3. Pathogenicity to Spodoptera frugipera
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tulloch, M. The genus metarhizium. Trans. Br. Mycol. Soc. 1976, 66, 407–411. [Google Scholar] [CrossRef]
- Chen, Z.; Yang, X.; Sun, N.; Xu, L.; Zheng, Y.; Yang, Y. Species diversity and vertical distribution characteristics of Metarhizium in Gaoligong Mountains, southwestern China. Biodivers. Sci. 2018, 26, 1308. [Google Scholar] [CrossRef]
- Sung, G.H.; Hywel-Jones, N.L.; Sung, J.M.; Luangsa-Ard, J.J.; Shrestha, B.; Spatafora, J.W. Phylogenetic classification of Cordyceps and the clavicipitaceous fungi. Stud. Mycol. 2007, 57, 5–59. [Google Scholar] [CrossRef] [PubMed]
- Kepler, R.; Sung, G.-H.; Ban, S.; Nakagiri, A.; Chen, M.-J.; Huang, B.; Li, Z.Z.; Spatafora, J. New teleomorph combinations in the entomopathogenic genus Metacordyceps. Mycologia 2012, 104, 182–197. [Google Scholar] [CrossRef] [PubMed]
- Kepler, R.M.; Humber, R.A.; Bischoff, J.F.; Rehner, S.A. Clarification of generic and species boundaries for Metarhizium and related fungi through multigene phylogenetics. Mycologia 2014, 106, 811–829. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Gautam, N. Metacordyceps dhauladharensis sp. nov., a new entomopathogenic fungus from India. Turk. J. Bot. 2015, 39, 520–526. [Google Scholar] [CrossRef]
- Chu, H.L.; Chen, W.H.; Wen, T.C.; Liang, Z.Q.; Zheng, F.C.; Liang, J.D.; Han, Y.F. Delimitation of a novel member of genus Metarhizium (Clavicipitaceae) by phylogenetic and network analysis. Phytotaxa 2016, 288, 51–60. [Google Scholar] [CrossRef]
- Luangsa-Ard, J.J.; Mongkolsamrit, S.; Thanakitpipattana, D.; Khonsanit, A.; Tasanathai, K.; Noisripoom, W.; Humber, R.A. Clavicipitaceous entomopathogens: New species in Metarhizium and a new genus Nigelia. Mycol. Prog. 2017, 16, 369–391. [Google Scholar] [CrossRef]
- Lopes, R.B.; Souza, D.A.; Rocha, L.F.; Montalva, C.; Luz, C.; Humber, R.A.; Faria, M. Metarhizium alvesii sp. nov.: A new member of the Metarhizium anisopliae species complex. J. Invertebr. Pathol. 2018, 151, 165–168. [Google Scholar] [CrossRef]
- Luz, C.; Rocha, L.F.; Montalva, C.; Souza, D.A.; Botelho, A.B.R.; Lopes, R.B.; Faria, M.; Júnior, I.D. Metarhizium humberi sp. nov. (Hypocreales: Clavicipitaceae), a new member of the PARB clade in the Metarhizium anisopliae complex from Latin America. J. Invertebr. Pathol. 2019, 166, 107216. [Google Scholar] [CrossRef] [PubMed]
- Mongkolsamrit, S.; Khonsanit, A.; Thanakitpipattana, D.; Tasanathai, K.; Noisripoom, W.; Lamlertthon, S.; Himaman, W.; Houbraken, J.; Samson, R.; Luangsa-Ard, J. Revisiting Metarhizium and the description of new species from Thailand. Stud. Mycol. 2020, 95, 171–251. [Google Scholar] [CrossRef]
- Thanakitpipattana, D.; Tasanathai, K.; Mongkolsamrit, S.; Khonsanit, A.; Lamlertthon, S.; Luangsa-Ard, J. Fungal pathogens occurring on Orthopterida in Thailand. Persoonia-Mol. Phylogeny Evol. Fungi 2020, 44, 140–160. [Google Scholar] [CrossRef]
- Yamamoto, K.; Ohmae, M.; Orihara, T. Metarhizium brachyspermum sp. nov. (Clavicipitaceae), a new species parasitic on Elateridae from Japan. Mycoscience 2020, 61, 37–42. [Google Scholar] [CrossRef]
- Chen, M.-J.; Lin, Y.; Wang, T.; Zhang, S.-L.; Huang, B. Metarhizium macrosemiae sp. nov. and the anamorph of M. guniujiangense on adult cicada from Guniujiang Nature Preserve, southeastern China. Phytotaxa 2022, 575, 68–78. [Google Scholar] [CrossRef]
- Chen, Z.H.; Dai, Y.D.; Chen, K.; Zhang, Y.F.; Xu, L.; Wang, Y.B. Papiliomyces puniceum and Metarhizium lymantriidae: Two new species from the Gaoligong Mountains in southwestern China. Phytotaxa 2023, 594, 53–63. [Google Scholar] [CrossRef]
- Driver, F.; Milner, R.J.; Trueman, J.W. A taxonomic revision of Metarhizium based on a phylogenetic analysis of rDNA sequence data. Mycol. Res. 2000, 104, 134–150. [Google Scholar] [CrossRef]
- Bischoff, J.F.; Rehner, S.A.; Humber, R.A. Metarhizium frigidum sp. nov.: A cryptic species of M. anisopliae and a member of the M. flavoviride complex. Mycologia 2006, 98, 737–745. [Google Scholar] [CrossRef]
- Bischoff, J.F.; Rehner, S.A.; Humber, R.A. A multilocus phylogeny of the Metarhizium anisopliae lineage. Mycologia 2009, 101, 512–530. [Google Scholar] [CrossRef]
- Nishi, O.; Iiyama, K.; Yasunaga-Aoki, C.; Shimizu, S. Phylogenetic status and pathogenicity of Metarhizium majus isolated from a fruit beetle larva in Japan. Mycol. Prog. 2015, 14, 58. [Google Scholar] [CrossRef]
- Nishi, O.; Shimizu, S.; Sato, H. Metarhizium bibionidarum and M. purpureogenum: New species from Japan. Mycol. Prog. 2017, 16, 987–998. [Google Scholar] [CrossRef]
- Chen, Z.; Xu, L.; Yang, X.; Zhang, Y.; Yang, Y. Metarhizium baoshanense sp. nov., a new entomopathogen fungus from southwestern China. Pak. J. Zool. 2018, 50, 1739–1746. [Google Scholar] [CrossRef]
- Chen, Z.H.; Zhang, Y.G.; Yang, X.N.; Chen, K.; Liu, Q.; Xu, L. A new fungus Metarhizium gaoligongense from China. Int. J. Agric. Biol. 2018, 20, 2271–2276. [Google Scholar] [CrossRef]
- Perumal, V.; Kannan, S.; Alford, L.; Pittarate, S.; Geedi, R.; Elangovan, D.; Marimuthu, R.; Krutmuang, P. First report on the enzymatic and immune response of Metarhizium majus bag formulated conidia against Spodoptera frugiperda: An ecofriendly microbial insecticide. Front. Microbiol. 2023, 14, 1104079. [Google Scholar] [CrossRef] [PubMed]
- Nishi, O.; Iiyama, K.; Yasunaga-Aoki, C.; Shimizu, S. Comparison of the germination rates of Metarhizium spp. conidia from Japan at high and low temperatures. Lett. Appl. Microbiol. 2013, 57, 554–560. [Google Scholar] [CrossRef] [PubMed]
- González-Hernández, G.A.; Padilla-Guerrero, I.E.; Martínez-Vázquez, A.; Torres-Guzmán, J.C. Virulence Factors of the Entomopathogenic Genus Metarhizium. Curr. Protein Pept. Sci. 2020, 21, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.K.; Sridhar, J.; Murali-Baskaran, R.K.; Senthil-Nathan, S.; Kaushal, P.; Dara, S.K.; Arthurs, S. Microbial biopesticides for insect pest management in India: Current status and future prospects. J. Invertebr. Pathol. 2019, 165, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.B.; Wang, Y.; Fan, Q.; Duan, D.E.; Zhang, G.D.; Dai, R.Q.; Dai, Y.D.; Zeng, W.B.; Chen, Z.H.; Li, D.D. Multigene phylogeny of the family Cordycipitaceae (Hypocreales): New taxa and the new systematic position of the Chinese cordycipitoid fungus Paecilomyces hepiali. Fungal Divers. 2020, 103, 1–46. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; PCR Protoc: A Guide Methods; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Wang, Y.B.; Yu, H.; Dai, Y.D.; Wu, C.K.; Zeng, W.B.; Yuan, F.; Liang, Z.Q. Polycephalomyces agaricus, a new hyperparasite of Ophiocordyceps sp. infecting melolonthid larvae in southwestern China. Mycol. Prog. 2015, 14, 70. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, A.C.; Leclerque, A.; Manfrino, R.G.; Luz, C.; Ferrari, W.A.; Barneche, J.; García, J.J.; Lastra, C.C.L. Natural occurrence in Argentina of a new fungal pathogen of cockroaches, Metarhizium argentinense sp. nov. Fungal Biol. 2019, 123, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Montalva, C.; Collier, K.; Rocha, L.F.N.; Inglis, P.W.; Lopes, R.B.; Luz, C.; Humber, R.A. A natural fungal infection of a sylvatic cockroach with Metarhizium blattodeae sp. nov., a member of the M. flavoviride species complex. Fungal Biol. 2016, 120, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Y.; Liang, Z.; Whalley, A.; Yao, Y.; Liu, A. Cordyceps brittlebankisoides, a new pathogen of grubs and its anamorph, Metarhizium anisopliae var. majus. J. Invertebr. Pathol. 2001, 78, 178–182. [Google Scholar] [CrossRef]
- Sigler, L.; Gibas, C.F.C.; Kokotovic, B.; Bertelsen, M.F. Disseminated mycosis in veiled chameleons (Chamaeleo calyptratus) caused by Chamaeleomyces granulomatis, a new fungus related to Paecilomyces viridis. J. Clin. Microbiol. 2010, 48, 3182–3192. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, K.; Ōmura, S.; Masuma, R.; Kaifuchi, S.; Masuma, R. Three new Pochonia taxa (Clavicipitaceae) from soils in Japan. Mycologia 2013, 105, 1202–1218. [Google Scholar] [CrossRef] [PubMed]
- Petch, T. Notes on entomogenous fungi. Trans. Br. Mycol. Soc. 1931, 16, 55–75. [Google Scholar] [CrossRef]
- Chen, W.; Han, Y.; Liang, J.; Liang, Z.; Jin, D. Metarhizium dendrolimatilis, a novel Metarhizium species parasitic on Dendrolimus sp larvae. Mycosphere 2017, 8, 31–37. [Google Scholar] [CrossRef]
- Chen, W.-H.; Han, Y.-F.; Liang, J.-D.; Liang, Z.-Q. Morphological and phylogenetic characterization of novel Metarhizium species in Guizhou, China. Phytotaxa 2019, 419, 189–196. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, X.-C.; Wu, L.-X.; Wang, Y.; Xu, A.; Lin, W.-F. Blackwellomyces kaihuaensis and Metarhizium putuoense (Hypocreales), Two New Entomogenous Fungi from Subtropical Forests in Zhejiang Province, Eastern China. Forests 2023, 14, 2333. [Google Scholar] [CrossRef]
- Kepler, R.M.; Sung, G.H.; Harada, Y.; Tanaka, K.; Tanaka, E.; Hosoya, T.; Bischoff, J.F.; Spatafora, J.W. Host jumping onto close relatives and across kingdoms by Tyrannicordyceps (Clavicipitaceae) gen. nov. and Ustilaginoidea (Clavicipitaceae). Am. J. Bot. 2012, 99, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Borisade, O.; Magan, N. Growth and sporulation of entomopathogenic Beauveria bassiana, Metarhizium anisopliae, Isaria farinosa and Isaria fumosorosea strains in relation to water activity and temperature interactions. Biocontrol Sci. Technol. 2014, 24, 999–1011. [Google Scholar] [CrossRef]
- Li, M.; Li, S.; Xu, A.; Lin, H.; Chen, D.; Wang, H. Selection of Beauveria isolates pathogenic to adults of Nilaparvata lugens. J. Insect Sci. 2014, 14, 32. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Nájera, R.E.; Ruiz-Estudillo, R.A.; Sánchez-Yáñez, J.M.; Molina-Ochoa, J.; Skoda, S.R.; Coutiño-Ruiz, R.; Pinto-Ruiz, R.; Guevara-Hernández, F.; Foster, J.E. Occurrence of entomopathogenic fungi and parasitic nematodes on Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae collected in central Chiapas, México. Fla. Entomol. 2013, 96, 498–503. [Google Scholar] [CrossRef]
- Vivekanandhan, P.; Swathy, K.; Alford, L.; Pittarate, S.; Subala, S.P.R.R.; Mekchay, S.; Elangovan, D.; Krutmuang, P. Toxicity of Metarhizium flavoviride conidia virulence against Spodoptera litura (Lepidoptera: Noctuidae) and its impact on physiological and biochemical activities. Sci. Rep. 2022, 12, 16775. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez-García, M.; Bustillos-Rodríguez, J.C.; Loya-Márquez, J.; Ríos-Velasco, C.; Jacobo-Cuellar, J.L. Parasitoides de Spodoptera frugiperda (JE Smith)(Lepidoptera: Noctuidae) en Chihuahua, México. Métodos En Ecol. Y Sist. 2015, 10, 67–72. [Google Scholar]
- Zemek, R.; Konopická, J.; Jozová, E.; Skoková Habuštová, O. Virulence of Beauveria bassiana strains isolated from cadavers of Colorado potato beetle, Leptinotarsa decemlineata. Insects 2021, 12, 1077. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Molecular Marker | Primer Name | Primer Sequence (5′–3′) | Reference |
|---|---|---|---|
| ITS | ITS4 | TCCTCCGCTTATTGATATGC | [28] |
| ITS5 | GGAAGTAAAAGTCGTAACAAGG | [28] | |
| tef-1α | EF1α-EF | GCTCCYGGHCAYCGTGAYTTYAT | [17,18] |
| EF1α-ER | ATGACACCRACRGCRACRGTYTG | [3] | |
| rpb1 | RPB1-5′F | CAYCCWGGYTTYATCAAGAA | [17,18] |
| RPB1-5′R | CCNGCDATNTCRTTRTCCATRTA | [3] | |
| rpb2 | RPB2-5′F | CCCATRGCTTGTYYRCCCAT | [17] |
| RPB2-5′R | GAYGAYMGWGATCAYTTYGG | [3] | |
| nrLSU | LR5 | ATCCTGAGGGAAACTTC | [29] |
| LROR | GTACCCGCTGAACTTAAGC | [30] | |
| nrSSU | nrSSU-CoF | TCTCAAAGATTAAGCCATGC | [31] |
| nrSSU-CoR | TCACCAACGGAGACCTTG | [31] | |
| 5′tef | EF1α-IF | ATGGGTAAGGASGAMAAGAC | This study |
| EF1α-IR | GGARGTACCAGTRATCATGTT | This study |
| Species | Voucher Information | GenBank Accession Number | Reference | ||||||
|---|---|---|---|---|---|---|---|---|---|
| SSU | LSU | tef | rpb1 | rpb2 | ITS | 5′tef | |||
| Metarhizium acridum | ARSEF 7486T | – | – | EU248845 | EU248897 | EU248925 | HQ331458 | EU248845 | [18] |
| M. acridum | ARSEF 324 | – | – | EU248844 | EU248896 | EU248924 | HM055449 | EU248844 | [18] |
| M. album | ARSEF 2082 | DQ522560 | DQ518775 | DQ522352 | DQ522398 | DQ522452 | AY375446 | – | [3] |
| M. album | ARSEF 2179 | – | – | KJ398807 | KJ398618 | – | HM055452 | – | [4] |
| M. alvesii | CG 1123T | – | – | KY007614 | KY007612 | KY007613 | – | KC520541 | [9] |
| M. anisopliae | ARSEF 7487 | – | – | DQ463996 | DQ468355 | DQ468370 | MH604974 | DQ463996 | [18] |
| M. anisopliae | CBS 130.71T | MT078868 | MT078853 | MT078845 | MT078861 | MT078918 | MT078884 | MT078928 | [11] |
| M. argentinense | CEP 424T | – | – | MF966624 | MF966625 | MF966626 | – | – | [37] |
| M. argentinense | CEP 414 | – | – | – | – | – | MF784813 | – | [37] |
| M. atrovirens | TNM-F 10184 | JF415950 | JF415966 | – | JN049884 | – | JN049882 | – | [4] |
| M. baoshanense | BUM 63.4 | KY264178 | KY264175 | KY264170 | KY264181 | KY264184 | KY264173 | – | [21] |
| M. baoshanense | CCTCC M 2016588 | KY087812 | KY087816 | KY087820 | KY087824 | KY087826 | KY087808 | – | [22] |
| M. baoshanense | CCTCC M 2016589T | KY264177 | KY264174 | KY264169 | KY264180 | KY264183 | KY264172 | – | [21] |
| M. bibionidarum | CBS 64867 | – | – | LC126075 | LC125907 | LC125923 | – | AB807450 | [20] |
| M. bibionidarum | NBRC 112661T | – | – | LC126076 | LC125908 | LC125924 | – | LC187676 | [20] |
| M. bibionidarum | Hkd35-2 | – | – | LC126077 | LC125912 | LC125925 | – | AB807804 | [20] |
| M. bibionidarum | KUNCC 10806 | PP256146 | PP256148 | PP328481 | PP294690 | PP314008 | PP256140 | PP413396 | This study |
| M. bibionidarum | KUNCC 10807 | – | PP256149 | PP328482 | PP294691 | PP314009 | PP256141 | PP413397 | This study |
| M. bibionidarum | KUNCC 10808 | PP256145 | PP256147 | PP328480 | PP294689 | PP314007 | PP256139 | PP413395 | This study |
| M. biotecense | BCC 51812T | MN781937 | MN781838 | MN781693 | MN781745 | MN781792 | MN781878 | – | [11] |
| M. biotecense | BCC 51813 | MN781938 | MN781839 | MN781694 | MN781746 | MN781793 | MN781879 | – | [11] |
| M. blattodeae | ARSEF 12850T | – | – | KU182917 | KU182918 | KU182916 | KU182915 | – | [38] |
| M. blattodeae | MY 00896 | HQ165657 | HQ165719 | HQ165678 | HQ165739 | HQ165638 | HQ165697 | – | [8] |
| M. brachyspermum | CM 1T | – | LC469749 | LC469751 | – | – | LC469747 | LC469752 | [13] |
| M. brasiliense | ARSEF 2948T | – | – | KJ398809 | KJ398620 | – | – | – | [5] |
| M. brittlebankisoides | Hn 1 | – | – | AB778556 | AB778555 | AB778554 | – | – | [19] |
| M. brittlebankisoides | G 97025 | – | – | – | – | – | AJ309332 | – | [39] |
| M. brunneum | ARSEF 2107T | – | – | EU248855 | EU248907 | EU248935 | KC178691 | EU248855 | [18] |
| M. brunneum | CBS 316.51 | MT078875 | MT078860 | MT078852 | – | – | MT078888 | MT078927 | [11] |
| M. campsosterni | BUM 10 | MH143832 | MH143815 | MH143849 | MH143864 | MH143879 | MH143798 | – | [2] |
| M. candelabrum | BCC 29224T | MN781952 | MN781853 | MN781708 | MN781755 | MN781804 | MN781881 | – | [11] |
| M. cercopidarum | BCC 31660T | MN781953 | – | MN781709 | MN781756 | MN781805 | MN781880 | – | [11] |
| M. chaiyaphumense | BCC 28241 | MN781932 | MN781831 | MN781684 | MN781740 | MN781784 | MN781884 | – | [11] |
| M. chaiyaphumense | BCC 78198T | KX369596 | KX369593 | KX369592 | KX369594 | KX369595 | MT078881 | – | [8] |
| M. cicadae | BCC 48696 | MN781948 | MN781848 | MN781703 | – | MN781800 | MN781885 | – | [11] |
| M. cicadae | BCC 48881T | MN781949 | MN781849 | MN781704 | MN781752 | – | – | – | [11] |
| M. clavatum | BCC 84543T | – | MN781834 | MN781689 | MN781741 | MN781789 | MN781886 | MT078929 | [11] |
| M. clavatum | BCC 84558 | – | MN781835 | MN781690 | MN781742 | – | – | – | [11] |
| M. culicidarum | BCC 2673 | MN781950 | MN781851 | MN781706 | MN781753 | MN781802 | MN781887 | – | [11] |
| M. culicidarum | BCC 7600T | MN781951 | MN781852 | MN781707 | MN781754 | MN781803 | MN781889 | – | [11] |
| M. culicidarum | BCC 7625 | – | MN781850 | MN781705 | – | MN781801 | MN781888 | – | [11] |
| M. cylindrosporum | CBS 256.90T | – | MH873892 | KJ398783 | KJ398594 | KJ398691 | MH862209 | – | [5] |
| M. cylindrosporum | RCEF 3632 | JF415964 | JF415987 | JF416022 | – | – | JN049872 | – | [4] |
| M. cylindrosporum | TNS-F 16371 | JF415963 | JF415986 | JF416027 | JN049902 | – | – | – | [4] |
| M. dianzhongense | KUNCC 10809 | – | PP256151 | PP328484 | PP294693 | PP314011 | PP256143 | - | This study |
| M. dianzhongense | KUNCC 10810 | – | PP256152 | PP328485 | PP294694 | PP314012 | PP256144 | - | This study |
| M. dianzhongense | KUNCC 10811 | – | PP256150 | PP328483 | PP294692 | PP314010 | PP256142 | - | This study |
| M. eburneum | BCC 79252T | – | MN781829 | MN781682 | MN781736 | – | MN781914 | – | [11] |
| M. eburneum | BCC 79267 | – | MN781826 | – | MN781735 | – | MN781915 | – | [11] |
| M. ellipsoideum | BCC 12847 | MN781959 | MN781860 | MN781715 | MN781761 | MN781810 | MN781925 | – | [11] |
| M. ellipsoideum | BCC 49285T | MN781957 | MN781858 | MN781713 | MN781759 | MN781808 | MT078876 | – | [11] |
| M. ellipsoideum | BCC 53509 | MN781958 | MN781859 | MN781714 | MN781760 | MN781809 | MT078877 | – | [11] |
| M. flavoviride | CBS 700.74 | MT078870 | MT078855 | MT078847 | MT078863 | MT078920 | – | MT078925 | [11] |
| M. flavoviride | ARSEF 2133 | – | – | – | – | – | NR131992 | DQ463988 | [17] |
| M. flavoviride | CBS 125.65 | MT078869 | MT078854 | MT078846 | MT078862 | MT078919 | MT078885 | MT078926 | [11] |
| M. flavoviride | CBS 218.56T | – | – | KJ398787 | KJ398598 | – | – | – | [5] |
| M. flavum | BCC 90870T | MN781965 | MN781874 | MN781731 | MN781776 | MN781822 | – | – | [11] |
| M. flavum | BCC 90874 | MN781966 | MN781875 | MN781732 | MN781777 | MN781823 | – | – | [11] |
| M. frigidum | ARSEF 4124T | – | – | DQ464002 | DQ468361 | DQ468376 | NR132012 | – | [17] |
| M. fusoideum | BCC 28246T | MN781944 | MN781844 | MN781699 | MN781749 | MN781796 | MN781893 | – | [11] |
| M. fusoideum | BCC 41242 | MN781942 | MN781825 | MN781679 | – | MN781780 | – | – | [11] |
| M. fusoideum | BCC 53130 | MN781943 | MN781843 | MN781698 | – | MN781795 | MN781894 | – | [11] |
| M. gaoligongense | BUM 3.5 | KY087810 | KY087814 | KY087818 | KY087822 | – | KY087806 | – | [22] |
| M. gaoligongense | CCTCC M 2016588T | KY087812 | KY087816 | KY087820 | KY087824 | KY087826 | KY087808 | – | [22] |
| M. globosum | ARSEF 2596T | – | – | EU248846 | EU248898 | EU248926 | NR132020 | – | [18] |
| M. granulomatis | UAMH 11028T | HM635076 | HM195304 | KJ398781 | – | – | NR132013 | – | [40] |
| M. granulomatis | UAMH 11176 | – | HM635078 | KJ398782 | KJ398593 | – | HM195306 | – | [40] |
| M. gryllidicola | BCC 37915 | – | – | – | – | – | MN781896 | – | [12] |
| M. gryllidicola | BCC 37918 | MN781935 | MN781836 | MN781691 | MN781743 | MN781790 | MN781897 | – | [12] |
| M. gryllidicola | BCC 82988 | MK632117 | MK632091 | MK632062 | MK632166 | MK632143 | – | MT078891 | [12] |
| M. guizhouense | ARSEF 6238 | – | – | EU248857 | EU248909 | EU248937 | – | EU248857 | [18] |
| M. guizhouense | CBS 258.90T | – | – | EU248862 | EU248914 | EU248942 | HQ331448 | EU248862 | [18] |
| M. guniujiangense | RCEF 6740 | MW718230 | MW718256 | MW723108 | MW723140 | MW723161 | – | – | [14] |
| M. guniujiangense | RCEF 880T | MW718229 | MW718255 | MW723106 | MW723139 | MW723160 | – | – | [14] |
| M. huainamdangense | BCC 32190 | MN781954 | MN781855 | MN781710 | MN781757 | – | MN781899 | – | [11] |
| M. huainamdangense | BCC 44270T | MN781956 | MN781857 | MN781712 | – | MN781807 | MN781898 | – | [11] |
| M. huainamdangense | BCC 7672 | MN781955 | MN781856 | MN781711 | MN781758 | MN781806 | MN781901 | – | [11] |
| M. humberi | IP 46T | – | – | MH837574 | MH837556 | MH837565 | – | JQ061205 | [10] |
| M. indigoticum | TNS-F 18553 | JF415953 | JF415968 | JF416010 | JN049886 | JF415992 | JN049874 | – | [4] |
| M. indigoticum | TNS-F 18554 | JF415952 | JF415969 | JF416011 | JN049887 | JF415993 | JN049875 | – | [4] |
| M. kalasinense | BCC 53582T | KC011175 | KC011183 | KC011189 | – | – | KC011179 | KX823945 | [8] |
| M. koreanum | ARSEF 2038T | – | – | KJ398805 | KJ398615 | KJ398713 | HM055431 | – | [5] |
| M. koreanum | BCC 27998 | MN781945 | MN781845 | MN781700 | – | MN781797 | MN781903 | – | [11] |
| M. koreanum | BCC 30455 | MN781946 | MN781846 | MN781701 | MN781750 | MN781798 | MN781904 | – | [11] |
| M. lepidiotae | ARSEF 7488T | – | – | EU248865 | EU248917 | EU248945 | HQ331456 | EU248865 | [18] |
| M. macrosemiae | RCEF 6696T | MW718233 | MW718259 | MW723113 | MW723144 | MW723164 | MW718317 | – | [14] |
| M. majus | ARSEF 1015 | – | – | EU248866 | EU248918 | EU248946 | HQ331444 | EU248866 | [18] |
| M. majus | ARSEF 1914T | – | – | EU248868 | EU248920 | EU248948 | HQ331445 | EU248868 | [18] |
| M. megapomponiae | BCC 25100T | MN781947 | MN781847 | MN781702 | MN781751 | MN781799 | MN781906 | – | [11] |
| M. minus | ARSEF 1099 | – | – | KJ398799 | KJ398608 | KJ398706 | – | – | [5] |
| M. minus | ARSEF 2037T | AF339580 | AF339531 | DQ522353 | DQ522400 | DQ522454 | AF138271 | – | [3] |
| M. niveum | BCC 52400T | MN781933 | MN781832 | MN781685 | – | MN781785 | MN781907 | – | [11] |
| M. nornnoi | BCC 19364 | MN781940 | MN781841 | MN781696 | MN781747 | – | MN781891 | – | [11] |
| M. nornnoi | BCC 25948T | MN781941 | MN781842 | MN781697 | MN781748 | – | MN781892 | – | [11] |
| M. novozealandicum | ARSEF 4661 | – | – | KJ398811 | KJ398622 | – | – | – | [5] |
| M. novozealandicum | ARSEF 4674 | – | – | KJ398812 | KJ398623 | – | – | – | [5] |
| M. ovoidosporum | BCC 29223 | MN781960 | MN781861 | MN781716 | MN781762 | – | MN781909 | – | [11] |
| M. ovoidosporum | BCC 32600T | MN781961 | MN781862 | MN781717 | MN781763 | – | MN781910 | – | [11] |
| M. ovoidosporum | BCC 7634 | MN781962 | MN781863 | MN781718 | MN781764 | MN781811 | MN781908 | – | [11] |
| M. owariense | NBRC 33258 | HQ165669 | HQ165730 | JF416017 | KJ398596 | JF415996 | JN049883 | – | [4] |
| M. pemphigi | ARSEF 6569 | – | – | KJ398813 | KJ398624 | DQ468378 | – | – | [5] |
| M. pemphigi | ARSEF 7491 | – | – | KJ398819 | KJ398629 | DQ468379 | – | – | [5] |
| M. pemphigi | BUM 1 | – | – | – | – | – | MH143795 | – | [22] |
| M. pemphigi | BUM 39.4 | – | – | – | – | – | KY087809 | – | [22] |
| M. phasmatodeae | BCC 2841 | MN781931 | MN781828 | MN781681 | MN781738 | MN781782 | MN781911 | – | [12] |
| M. phasmatodeae | BCC 49272 | MK632119 | MK632093 | MK632064 | – | MK632145 | MK632035 | MT078893 | [12] |
| M. phuwiangense | BCC 78206 | – | – | MN781719 | MN781765 | MN781812 | MT078879 | – | [11] |
| M. phuwiangense | BCC 85068 | – | MN781864 | MN781720 | MN781766 | MN781813 | MN781912 | – | [11] |
| M. phuwiangense | BCC 85069T | – | MN781865 | MN781721 | MN781767 | MN781814 | MN781913 | – | [11] |
| M. pingshaense | CBS 257.90T | – | – | EU248850 | EU248902 | EU248930 | HQ331450 | EU248850 | [18] |
| M. prachinense | BCC 47950 | KC011172 | KC011180 | KC011186 | KC011184 | – | KC011176 | – | [8] |
| M. prachinense | BCC 47979T | KC011173 | KC011181 | KC011187 | KC011185 | – | KC011177 | – | [8] |
| M. pseudoatrovirens | TNS-F 16380 | – | JF415977 | KJ398785 | JN049893 | JF415997 | JN049870 | – | [4] |
| M. purpureogenum | ARSEF 12570 | – | – | LC126079 | LC125911 | LC125922 | – | – | [8] |
| M. purpureogenum | ARSEF 12571T | – | AB700552 | LC126078 | LC125913 | LC125920 | – | – | [8] |
| M. purpureonigrum | BCC 89247T | – | – | MN781725 | MN781771 | MN781817 | – | – | [11] |
| M. purpureonigrum | BCC 89248 | MN781964 | MN781870 | MN781728 | – | MN781819 | – | – | [11] |
| M. purpureonigrum | BCC 89249 | MN781963 | MN781869 | MN781726 | MN781772 | MN781818 | – | – | [11] |
| M. purpureum | BCC 82173 | – | MN781866 | MN781722 | MN781768 | MN781815 | MN781919 | – | [11] |
| M. purpureum | BCC 82642T | – | MN781867 | MN781723 | MN781769 | MN781816 | MN781918 | – | [11] |
| M. purpureum | BCC 83548 | – | MN781868 | MN781724 | MN781770 | – | MN781920 | – | [11] |
| M. reniforme | ARSEF 429 | HQ165671 | HQ165733 | HQ165690 | – | HQ165650 | DQ069284 | – | [8] |
| M. reniforme | ARSEF 577 | HQ165672 | HQ165734 | HQ165691 | – | HQ165651 | DQ069283 | – | [8] |
| M. reniforme | IndGH 96 | HQ165670 | HQ165732 | – | – | HQ165649 | – | – | [8] |
| M. rileyi | AF 368501 | – | – | – | – | – | AF368501 | – | [16] |
| M. rileyi | CBS 806.71 | AY526491 | MH872111 | EF468787 | EF468893 | EF468937 | AY624205 | – | [3] |
| M. rileyi | NBRC 8560 | HQ165667 | HQ165729 | HQ165688 | – | – | – | – | [8] |
| M. robertsii | ARSEF 4739 | – | – | EU248848 | EU248900 | EU248928 | – | EU248848 | [18] |
| M. samlanense | BCC 17091T | HQ165665 | HQ165727 | HQ165686 | – | HQ165646 | HQ165707 | – | [8] |
| M. samlanense | BCC 39752 | MN781939 | MN781840 | MN781695 | – | MN781794 | MT078880 | – | [8] |
| M. sulphureum | BCC 36585 | – | – | MN781686 | – | MN781786 | – | MT078931 | [11] |
| M. sulphureum | BCC 36592T | – | – | MN781687 | – | MN781787 | – | – | [11] |
| M. sulphureum | BCC 39045 | MK632120 | MK632095 | MK632066 | – | MK632147 | MK632037 | MT078930 | [11] |
| M. takense | BCC 30934 | HQ165658 | HQ165720 | HQ165679 | HQ165740 | HQ165639 | HQ165698 | – | [8] |
| M. takense | BCC 30939T | HQ165659 | HQ165721 | – | HQ165741 | HQ165640 | HQ165699 | – | [8] |
| M. viride | CBS 659.71 | HQ165673 | HQ165735 | HQ165692 | – | HQ165652 | HQ165714 | – | [14] |
| M. viridulum | ARSEF 6927 | – | – | KJ398815 | KJ398681 | – | – | – | [5] |
| M. viridulum | BCC 36261 | MN781930 | MN781827 | MN781680 | MN781737 | MN781781 | MT078878 | – | [11] |
| Pochonia boninensis | JCM 18597T | AB758255 | AB709831 | AB758463 | AB758666 | AB758693 | AB709858 | – | [41] |
| P. chlamydosporia | CBS 101244 | DQ522544 | DQ518758 | DQ522327 | DQ522372 | DQ522424 | JN049821 | – | [41] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, C.-Y.; Tang, M.; Xie, L.-Y.; Fan, Q.; Shen, S.-K.; Yang, Z.-L.; Deng, G.; Wang, Y.-B. Metarhizium dianzhongense sp. nov. and New Record of M. bibionidarum (Clavicipitaceae, Hyocreales) Attacking Insects from China. Diversity 2024, 16, 201. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040201
Wei C-Y, Tang M, Xie L-Y, Fan Q, Shen S-K, Yang Z-L, Deng G, Wang Y-B. Metarhizium dianzhongense sp. nov. and New Record of M. bibionidarum (Clavicipitaceae, Hyocreales) Attacking Insects from China. Diversity. 2024; 16(4):201. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040201
Chicago/Turabian StyleWei, Cui-Yuan, Mei Tang, Liu-Yi Xie, Qi Fan, Shi-Kang Shen, Zhu-Liang Yang, Gang Deng, and Yuan-Bing Wang. 2024. "Metarhizium dianzhongense sp. nov. and New Record of M. bibionidarum (Clavicipitaceae, Hyocreales) Attacking Insects from China" Diversity 16, no. 4: 201. https://0-doi-org.brum.beds.ac.uk/10.3390/d16040201