


Species Composition and Distribution of Terrestrial Herbs in a High Montane Forest in Ecuador

1
Escuela de Ciencias Biológicas, Pontificia Universidad Católica del Ecuador, Quito 170143, Ecuador
2
Ecoinformatics & Biodiversity, Department of Biology, Aarhus University, 8000 Aarhus, Denmark
*
Author to whom correspondence should be addressed.
Diversity 2024, 16(5), 262; https://0-doi-org.brum.beds.ac.uk/10.3390/d16050262
Submission received: 27 January 2024
/
Revised: 15 April 2024
/
Accepted: 16 April 2024
/
Published: 27 April 2024
(This article belongs to the Special Issue Diversity Hotspots)
Abstract
:In mountain tropical forests, understory herbs have received little attention compared to trees, and their commonness and rarity are virtually unknown. We studied ground herbs to explore how they are assembled in a full one-hectare plot and to test the influence of light intensity (LI) and topographic habitats in species composition. The plot is a humid montane forest located in the Pasochoa Volcano, at 3300 m. We found 43 genera and 50 perennial species (30 angiosperms in 17 families, and 20 ferns). Interestingly, herbs are 64% richer in species than trees in the same study plot (50 vs. 32). Herbs were mostly obligately terrestrial (70% of the species), while 30% were fallen climbers and epiphytes rooted in the ground. Across the forest, there were 31,119 individuals that covered 8.5% of the ground. We concluded that both LI and topography shaped the species distribution, the floristic composition, and the community structure of ground herbs. For instance, 12% of the species were exclusively found in places with high LI; the rest of species grew in medium- to low-LI environments. Concerning rarity, we found that 39% of the species are rare (judging by botanical collections; <100), which stresses the need of conservation strategies for this group of plants.
1. Introduction
The richest plant community on Earth is the understory of the tropical rain forest [1,2,3]; however, only few ecological studies have focused on terrestrial herbs. Concerning neotropical rain forests, most of the information about structure, composition, and diversity deals with trees [4,5,6,7]. Few quantitative studies include all life forms or other life forms besides trees [1,2,8,9,10,11,12,13,14,15,16,17,18,19]. These studies have shown that the herbaceous flora is very important for the forest structure, emphasizing the fact that forests are not built just by trees, and that the herbs growing in the forest increased the forest’s species richness.
Researchers have included non-woody plants, such as pteridophytes, in ecological studies of tropical forest structure [19,20,21]. At scales of one hectare or less, the floristic composition of herbs is correlated with topography, soil, biological factors, biogeography, and chance [9,10,13,16,18]. At continental and global scales, the ecological importance of non woody plants like herbs and their high beta diversity have been documented in different regions and forest types [8,11,22].
In the tropical Andean region, ecological, taxonomical, and floristic studies of herbs have been concentrated in the páramos [23,24,25,26,27,28,29,30,31,32]. These studies were focused on alpha and beta diversity patterns, growth form composition, plant community structure, regeneration of páramo vegetation after burning and grazing, environmental factors shaping diversity patterns, ecosystem function, changes, and tolerance to climate change [17,33,34]. There are no quantitative studies of herbs in the high montane forest of the Andes >1000 m. Most studies of herbs in the Andes are concentrated on the lower slopes. The major source of information about herbs are botanical collections and flora inventories carried out at these elevations. At the scale of one hectare, to our knowledge, there are only two studies both carried out in tropical lowland forests, one in Southeast Asia, in Borneo [15], and in the Amazon lowlands, in Ecuador [13]. In these studies, a complete enumeration of herbs allowed us to understand species associations with microhabitats and detect patterns of floristic composition. However, worldwide studies of herbs are usually carried out in smaller areas (<5 ha) [9,19,35,36,37,38].
The drivers that trigger herb species composition are mostly related to light, topography, water, and soil. Mostly, these studies were carried out in lowland tropical forests of Borneo, the Ecuadorian Amazon, and India [15,20,39]. The drivers shaping species composition of Andean herbs are unknown; therefore, this study will shed light on this topic to understand species composition in Ecuadorian high Andes.
Because of the historical deforestation of and pressure placed on open agricultural lands, the high montane forests in Ecuador are scarce, covering less than 5% in Ecuador [40], and they are confined to less accessible areas. It is crucial to understand the little-known ecology of herbs in this endangered ecosystem and offer science-based information to take conservation actions.
The conservation status of endemic herbs will give some us some information for directing scarce conservation resources to the species most in need. Unfortunately, the conservation status of most Ecuadorian herbs is unknown, except for endemic species. How rare and endangered a species is needs to be traced urgently.
We chose to study herbs in a tree study plot that was previously established in a high montane forest [41]. The present study describes the floristic composition of herbs in this one-hectare plot located at high elevation and immediately below the timberline of the Pasochoa volcano (3300 m). We carried out a detailed quantification of the herbs and explored their ecological importance (based on species density and cover) and their habitat associations; in particular, we ask to which extent the species composition, density, and cover is associated with topography and light availability on the forest floor. As part of this survey, we evaluated the conservation status of all the species recorded in the study plot.
2. Materials and Methods
2.1. Study Area
The Pasochoa volcano (0°28 S, 78°28 W) is a dormant volcano covered by volcanic ashes, heavily eroded by Pleistocene glaciations, that created a collapsed volcanic cone [42]. This volcano is situated 35 km south of Ecuador’s capital Quito, in the inter-Andean valleys with an altitudinal range from 2400 to 4200 m, where humid montane forests and paramos are present. The inter-Andean valleys have fertile soils that have been strongly affected for centuries by agriculture. The Pasochoa shelters a unique refuge of 500 ha of protected forest, declared in 1996 by the Ecuadorian National System of Protected Areas (SNAP) (http://areasprotegidas.ambiente.gob.ec/ (accessed on 13 April 2024)) [43]. This refuge is the best-preserved Ecuadorian inter-Andean forest. The most intact forest grows in the collapsed cone due to its inaccessibility. Besides the collapsed cone, there are several erosion gullies that shelter the well-preserved forest surrounding the external slopes of the Pasochoa volcano. The mid and lower slopes of the volcano are bordered by extensive pastures, planted pine forests, and croplands with potatoes, corn, and wheat. Provoked fires and cattle entering the forest areas are the major factors having an impact on the forest. According to the closest meteorological station at Uyumbicho, the mean temperature is 13.8 °C, and the mean annual precipitation is 1489.5 mm [41]. There are only three dry months (i.e., months with <100 mm precipitation): July, August (precipitation range 35–50 mm), and September (<100 mm). All other months are wet [41]. Even in the dry months, the humidity remained high enough to keep epiphytes like orchids and ferns alive. There were no annual herbs recorded in the study site during the fieldwork period (1996). Throughout the year, including the dry months, it was possible to find all the species reported in this study. Our field data collection took place during both the less rainy and the rainy seasons, yet discernible changes in species density or cover were not apparent.
The one ha plot is located at 3300 m elevation in one of the well-preserved erosion gulleys. The area has little accessibility and belongs to a private property (El Olimpo Farm), where the owners control and limit the accessibility to the land for furtive visitors and cattle. The plot area is on steep terrain with branched erosion gullies with slopes that can reach >100% (>45°), which shelter a mature forest [41,44]. The forest reaches 6–8 m in height and is dominated by trees that branch basally and have inclined or horizontal trunks. In the same one-hectare plot, Valencia and Jørgensen (1992) [41] found 32 species of trees > 5 cm dbh. The most species-rich tree families were Melastomataceae and Asteraceae. Miconia thaezans is the most abundant tree species, followed by Piper andreanum and Miconia pustulata. The forest floor is covered by mosses and hepatics, and the litter layer varies in thickness from 2 to 30 cm.
We choose the study plot for several reasons that make it representative of the forest: (1) the forest structure has trees of different sizes, including big trees (>30 cm dbh) that are seldom seen in less-preserved forests, lianas (some of them >10 cm of diameter), epiphytes, and underground herbs; (2) there has been limited human impact there, and it has remained excluded from cattle for more than 40 years [44]; (3) the plot was established in 1988 to study the forest structure and tree species richness, with both structure and species richness being comparable to other well-preserved Andean forests studied in Ecuador (Jorgensen et al. 1995) [45]; and (4) according to fieldwork observations and revision of herbarium specimens, the plot had a good representation of the understory herbaceous flora, which is different in composition of that found in the open paramos located over 3400 m, just above the tree line at the Pasochoa volcano.
2.2. Species Data
The one-hectare plot was divided into 400 subunits of 5 × 5 m, 100 subunits of 10 × 10 m, and 10 subunits of 10 × 100 m. The latter were oriented from east to west and were used for comparisons purposes. All herb species rooted in the ground were documented with voucher specimens and were identified. This included obligate terrestrial species, which are species that are always rooted in the ground, and facultatively terrestrial species, which are sometimes epiphytic or climbing and not rooted in the ground. The obligate terrestrial species were further divided into prototerrestrials with self-supporting stems and scandents, which depend on others for support. The facultative terrestrials were divided into fallen epiphytes and fallen climbers. The following data were collected for each species in each 5 × 5 m subunit: (1) number of individuals and (2) cover in cm2. For species with vegetative reproductive habits, each stem was counted as one individual. To describe the species importance in the forest community, we modified the Importance Value Index (IVI) used for trees [46] in our herb study. IVI = relative density + relative cover + relative frequency per hectare. We replaced the relative dominance (relative basal area) used for trees with cover (the area projected on the ground that covers a given species, expressed in m2). The sum of the three percentages conforms the index and allows to know and visualize which of these parameters are more relevant in the herb community for a given species.
2.3. Species Endemism and Conservation Status
Information about the endemism and conservation status of the species was searched for all the species. The risk of extinction of most species has not been previously evaluated, and we propose here such an evaluation based on historical collection records, the distribution of the species in Ecuador, and the number of collections recorded in the data bases of Missouri Botanical Garden (MBG; https://www.tropicos.org/home (accessed on 13 April 2024)), QCA Herbarium (Bioweb; https://bioweb.bio/portal/ (accessed on 13 April 2024)), and AAU Herbarium, Denmark (https://www.aubot.dk/search_form.php accessed on (13 April 2024)). The number of collections is a good proxy of the species commonness or rarity in nature. A species was designated as well collected (WC) if it had >100 collections all over Ecuador, medium collected (MC) if it had <100–>30, and scarcely collected (SC) with <30 collections all over Ecuador. To assign the risk of extinction categories, we used the IUCN criteria [47], and, in the case of the endemic species, we used the status reported by León-Yánez et al. (2011) [48].
2.4. Topographic Microhabitats
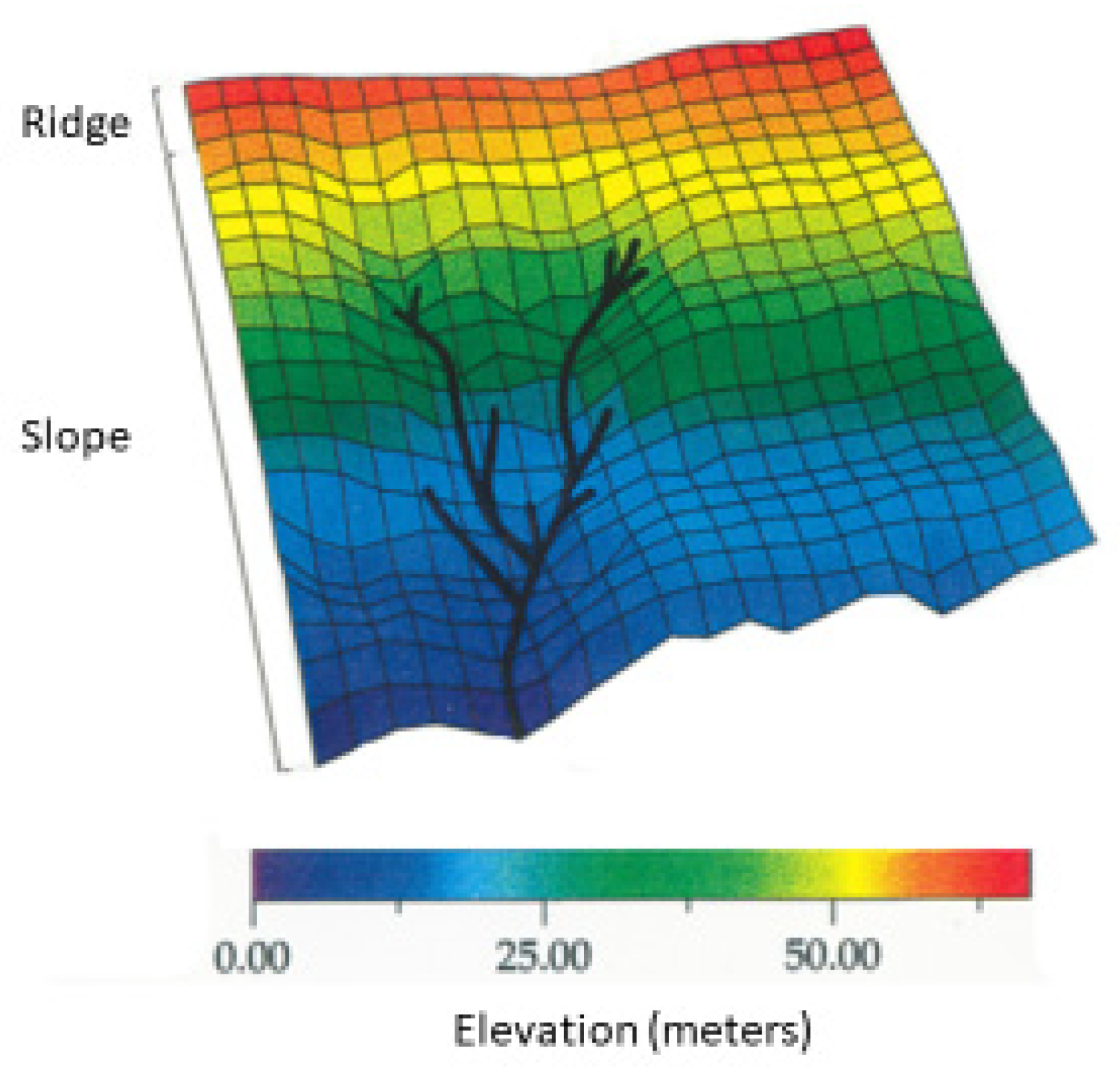
Each 5 × 5 m subunit was classified as belonging to one of three topographic microhabitats: ridge, erosion gully, or slope (Figure 1). The ridge microhabitat is the least steep terrain of the study plot, with relative altitudes of 59–67 m from the lowest point; it is limited to the first four rows of 5 × 100 m. The extension of this microhabitat is 1775 m2 (18% of the plot). The slope microhabitat is the rough part of the plot area, located at altitudes of 0–58.9 m. The extension of this zone is 7025 m2 (70% of the plot). The erosion gully bed is the only microhabitat that has bare rocks as part of the soil structure with rivulets running through it. The extension of this microhabitat is 1200 m2 (12% of the plot) at altitudes of 0–54 m.
2.5. Light Microhabitats
Each 5 × 5 m subunit was classified in one of the following three light microhabitats: low, medium, and high light intensity. The light microhabitats depended on both the woody vegetation structure (closed canopy or open canopy) and the relative cover of the woody scandent bamboo Chusquea scandens, (estimated as: 1 = presence, 2 = ≤50% cover, and 3 = >50% cover); a combination of both was used to determine the accessibility of light to the forest floor. The low light intensity microhabitat (thicket) had groups of stems of Chusquea scandens, tree branches, and lianas that twisted and formed a dense tangle, restricting the access of light to the bottom of the forest. This tangle was found 40–50 cm from the floor and was so compact that it produced an extreme shadow to the forest floor where only few herbs species could survive. The low light microhabitat covers 18% of the plot, and it is mostly (91% of its area) located at the lower part of the plot (0–10 m). The medium light intensity microhabitat has less than 50% cover of Chusquea scandens. This microhabitat was found in all three topographic microhabitats. It covered 60% of the plot. The high light intensity microhabitat had trees that produced little shadow to the forest floor. This microhabitat had limited presence of Chusquea scandens and lianas, no thicket, and was found in all topographic microhabitats covering 22% of the plot area.
2.6. Statistical Analysis
For the statistical analyses, we used the Mantel and Partial Mantel tests [49] to find out the association of different microhabitats with the species composition in the plot. We tested the correlations of the floristic matrix with the space matrix for the topographic habitats. For the floristic matrix, we used 80 units of 5 × 5 m (25 m2). We selected 20 units of each of the three main topographical habitats (ridge top, slope, and erosion gulley) and 20 units of light intensity habitats, 10 for high light intensity and 10 for low light intensity (thicket). Light intensity units were located in all topographical habitats. We built (1) a floristic matrix, (2) a topographic matrix, (3) a space (geographical distance) matrix, and (4) a presence/absence matrix of thicket for light intensity (excluding medium light intensity units).
For the floristic matrices, we used Sørensen and Steinhouse indices, but the results are presented with the Sørensen index alone because there were no differences between these two similarity matrices. We used Euclidean distances between centers of pairwise units for the space matrix. The statistical significance was estimated using the Montecarlo permutation test, allowing 999 permutations in each comparison.
3. Results
3.1. Species Composition and Richness
We found 50 herb species (Table 1) in 43 genera and 18 families (ferns were treated as one group). All ground herbs registered in the plot were perennials. Orchidaceae was the most species-rich family for the study area with six species, followed by Cyperaceae, Piperaceae, Rubiaceae, Caryophyllaceae, Urticaceae, and Asteraceae. The remaining 10 families were all represented by a single species (Table 1, Figure 2). Among the ferns (15 gen./20 spp.), Asplenium (6 spp.) was the only genus with more than one species (Table 1).
3.2. Life Forms
3.3. Species Endemism and Conservation Status
The Pasochoa herb flora presented just three endemic species, two of them being orchids, Stellaria recurvata and Baskervilla auriculata, and the third species being a Begoniaceae, Begonia sodiroi. All the statuses of the orchids are presented in the CITES Appendix II [51], which includes species not necessarily threatened with extinction, but in which trade must be controlled in order to avoid utilization incompatible with their survival [51]. Regarding rarity and commonness, 39% of the non-endemic species were scarcely collected, 32% were medium collected, and 29% were well collected (Table 1). Just one third of the non-endemic species had more than 100 collections in our survey.
3.4. Density
The entire plot had 31,119 individuals, i.e., three individuals/m2. Most of the 50 species formed dense groups, and it was sometimes difficult to count individual plants. The three species with the highest densities in our study area were Carex hamata (N = 7116), Polypodium subandino (N = 4620), and Peperomia rotundata (N = 4860). These three species contributed 53% of the total number of individual herbs. Five species had densities of 1000–2100 individuals and contributed 26% of the total number. There were 16 species with densities of 100–1000 individuals. In the other extreme, 26 species had <100 individuals and contributed only 2.5% of the total numbers of individuals (14 of these species had densities < to 30 individuals) (Table 1).
3.5. Cover
The total cover of herbs in the study site was 815 m2 (8.15%). In units of 25 m2, 100 m2, and 1000 m2 (10 × 100 m), the cover averaged 2, 8, and 81.5 m2, respectively (Table 2). Carex hamata (175 m2), Rhynchospora macrochaeta (90 m2), Thelypteris rigescens (68 m2), Peperomia rotundata (58 m2), and Polystichum muricatum (50 m2) contributed 52% to the total cover (Table 1). Most of the species in this list have relatively big leaves, except Carex hamata and Peperomia rotundata, whose high cover is due to their high number of individuals.
3.6. Indices of Importance Value
The Importance Value Index (IVI) for the 10 most important species (Figure 4) varies from 50 for Carex hamata to 8 for Alloplectus dielsii. Of these 10 species, 8 are prototerrestrials and 2 are climbers.
3.7. Microhabitats and Species Preferences
The spatial distribution maps (Figure 5a–d) show that only 8 species were restricted to specific topographic microhabitats, while the 42 remaining species were found in more than 1 microhabitat.
The species that were most associated with the ridge were Hydrocotyle alchemilloides (N = 260, Figure 5b), Galium obovatum Kunth (N = 39), and Leptostigma pilosum (N = 1159). These species were also found in other microhabitats, but they were denser in the ridge. Almost all the species that occurred in the slope were generalists that were also found in the other three topographic microhabitats (Table 1). The species that occurred exclusively on the slope all had very low densities (N < 10)—Hypolepis bogotensis (N = 8), Elaphoglossum papilosum (N = 2), and Pleurothalis sp. (N = 2), with the last two being facultative epiphytes. There were five species that occurred exclusively in the erosion gully—Asplenium squamosum (N = 10), Pteris muricata (N = 35), Equisetum bogotense (N = 48), Urtica leptophylla. (N = 8), and Peperomia sp. 2 (N = 24). These species were found in soils with high humidity, especially in water sources. Begonia sodiroi (N = 428) and Asplenium pteropus (N = 347) occurred especially in the erosion gully but also in the other two sites (ridge and slope).
The low light microhabitat (Figure 5e) had 12 species, none of which were exclusive to it. The most frequent species were Thelypteris rigescens, Polypodium subandino, Peperomia rotundata, Solanum dalibardiforme, Oxalis lotoides, Tournonia hookeriana, Arenaria lanuginose, and Stellaria recurvata. The density and cover of Thelypteris rigescens was lower in this microhabitat. Five species were exclusive to the middle light microhabitat (Figure 5f): Urtica leptophylla. (N = 8), Campyloneurum amphostenon (N = 31), Lomaria fragilis (N = 23), Adiantum concinnum (N = 6), and Peperomia sp. 2 (N = 24). In the high light microhabitat, we encountered species such as Baskervilla auriculata (N = 30), Cranichis ciliata (N = 208), Malaxis hoppii (N = 406), Hydrocotyle alchemilloides (N = 260), Anthurium pulchrum (N = 276), and Leptostigma pilosum (N = 1159), which were all exclusive to the microhabitat.
3.8. Statistical Analysis
There were positive Mantel correlations between the floristic composition of the herb flora in the montane forest plot that we studied at Pasochoa with the topographic, the light (presence/absence of thicket), and the spatial matrices (Table 3). The strongest correlations were seen with topography and light. The correlation with space was very weak and not significant.
4. Discussion
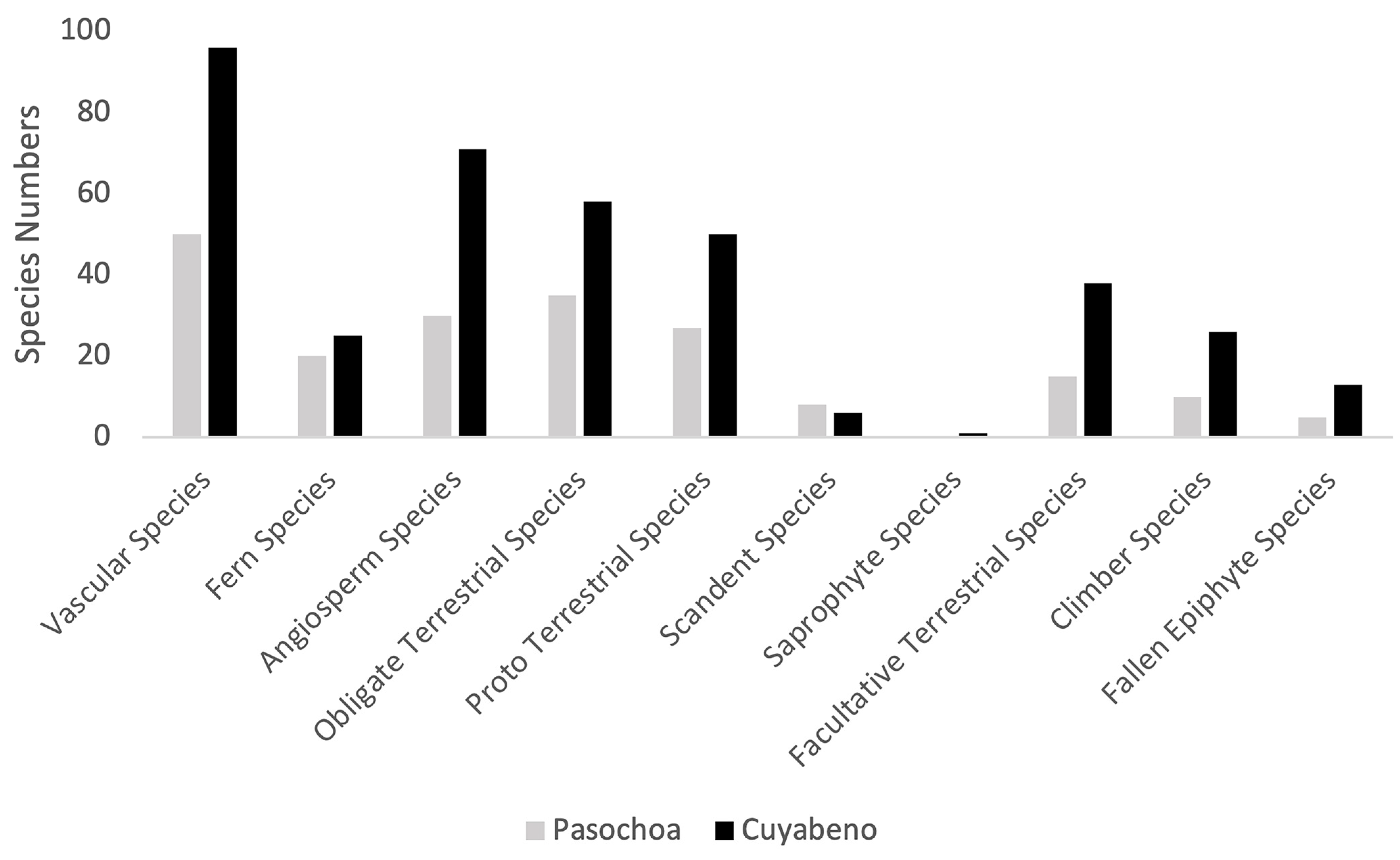
The high number of genera of terrestrial herbs (43) in the Pasochoa one-hectare plot stands out. The largest genus comprised six species. This is hard to understand unless the situation is focused on a large scale. In the entire Andean region of Ecuador above a 2400 m elevation, it is common to find families represented by a single genus (46%) and genera represented by a single species (21%) [52]. This overall pattern of the high Andean flora is reflected in the Pasochoa plot. The pattern is different from that found in Amazonian Ecuador, where most families are represented by more than one genus and more than one species [53]. Even in small plots (e.g., one hectare), such as the plot in the Amazon lowlands at Cuyabeno (250 m), where 96 herb species were found, the richest family, Araceae, had 28 species, followed by ferns (24) and Marantaceae (7) [13]. Pasochoa had 1.11 species/genus, while Cuyabeno had 2.08 species/genus. The most species-rich genera in Cuyabeno had >10 species compared with only 6 species found in the largest genus in Pasochoa.
The high number of rare species (71%) (Table 1), or not easily found within their geographical distribution range [54], should have high priority in conservation programs. Botanical exploration, collections [55,56,57], and quantitative inventories focused on Andean forest herbs can help to evaluate their conservation status and take action to preserve these groups that are frequently overlooked life forms in forest surveys. These findings showed how poor our knowledge of the Andean herbs is and raised a concern of the urgent need of a conservation plan.
In the Cuyabeno plot, 33% of the prototerrestrials were pteridophytes [14] (vs. 60% in our study site). This finding emphasizes the fact that even areas as small as one hectare have a relatively high number of fern species due to mountain-covered habitats that enable fern coexistence [58]. The high humidity encountered here is mostly due to fog; moreover, the low temperatures of the Andean mountains create appropriate conditions for the development of many pteridophyte species. The highest pteridophyte species richness is found in Ecuador as well as in other neotropical areas are between 1500 and 3000 m [58]. Other studies carried out at a large scale also showed that mountains are richer than lowlands in pteridophyte species [18,19,58,59,60]. If the fern flora of the entire Andes (2000 species) is compared with that of the entire Amazonian Brazil (300 species), it is obvious that ferns in the lowlands have a poor representation in the Neotropics [58].
Concerning the entire group of herb species, botanical accounts document that species richness increases with increasing altitude, reaching a maximum at 2400–3000 m [52]. However, in small areas like the one-hectare study at Cuyabeno, this pattern is the opposite: Cuyabeno included nearly twice as many species in comparison to Pasochoa at this scale (Figure 3). Whitmore and collaborators (1985) [12] used a category of free-standing herbs, which is equivalent to obligate terrestrials in the present study (Table 2). Gentry and Dodson (1987) [1] as well as Linares-Palomino and collaborators (2009) [19] used the classification of “terrestrial herbs”, which is the same as our obligate terrestrials, as the authors had separate categories for climbers and for epiphytes. Comparing the species richness of these studies with the Pasochoa plot (Figure 3, Table 2), we conclude that the Pasochoa plot had the lowest species richness, except if it would be compared with the dry forest of Jauneche located on the coast of Ecuador. This make sense, considering that most herb species, especially ferns, thrive in humid habitats.
4.1. Density
The density of herbs at the Pasochoa plot is three times as high as the density found at the Cuyabeno plot in the Amazon lowlands (Table 2). The density of herbs found at Pasochoa is higher than the densities found in most other study sites [1,12,15], but the density reported by Gentry and Dodson (1987) [1] for a 0.5 ha in a low mountain forest is comparable to our findings (Table 2).
In other tropical forests, such as the Barro Colorado Forest, trees had low densities because of the scarcity of habitats and because of unfavorable regeneration conditions. Therefore, the rare species do not have reproductive advantages over the common species [61]. We believe this also happens in herbs. The low densities of 26 species (<100 individuals/ha) reported in our study might be explained by the low ability of those species to grow in unstable soils, especially on the slopes (Table 1). For a comparison, species tend to be rarer in the lowland Amazon forest at Cuyabeno [13], where 50% of herb species contribute 7% of the total number of individuals.
In temperate and dry forests, alterations in seasonality can impact the density and cover of the species [39]. At our study site, even in the drier months of July and August, all the species, including the rare ones, were consistently present throughout the year. The lack of annual herbs is a characteristic of tropical humid forests [13,62]. Any potential fluctuation in density and cover between seasons likely did not substantially affect the patterns reported in our study.
4.2. Cover
The Amazonian forest plot at Cuyabeno and the plots in Borneo had lower covers compared to the Pasochoa forest: 2.50% and 0.72% vs. 8.15% (in this study) [13,15]. The vegetative growth (clones) of certain species found in the Pasochoa plot can explain this high cover. In contrast, in the Cuyabeno plot, only two species formed dense groups, Geophila and Araceae [13]; therefore, in the montane forests at Pasochoa, more species produce dense groups, constituting to larger covers.
4.3. Importance Value of Species and Microhabitat Associations
The high importance value of Orchidaceae (Figure 2) coincides with the fact that this family is the richest in species in Andean forests over 2400 m [52,63]. The high number of herb genera (Figure 2) represent a great phylogenetic richness in the study area, showing that diversification in the Andes is a continuous and actual process [64]. Certain species like Carex hamata, Polypodium subandinum, and Peperomia rotundata provide high density values, but lower values of frequency can be explained by the fact that vegetative spread gives herbs on the ground the mobility to find suitable edaphic and climatic microhabitats (Figure 4).
The 12 species thriving in the low-light microhabitat were primarily climbers facing challenges in finding exposed areas to reach sunlight. The Andean bamboo, Chusquea scandens, not only created shade but facilitated a framework for climbers (Oxalis lotoides, Arenaria lanuginose, and Stellaria recurvata) to eventually access light. In contrast, the prototerrestrial Thelypteris rigescens could survive in this challenging microhabitat due to its preference for shade, as described by Richards [65].
In terms of the floristic association with habitats (Mantel correlations), the composition of herbs exhibited a strong correlation with light availability (presence/absence of thicket) and topography. Although thicket units are concentrated in the steep slopes at the lower half of the plot (Figure 1; ~90% of the thicket/presence units are located here), light intensity is independently associated with species composition. The thicket structure had a significant effect on the species composition of ground herbs. Thicket creates low light intensity microhabitats where the species distribution and community structure is distinct. The impact of light availability in the understory herbs aligns with documented findings in Singapore [66] and in seasonally dry tropical forests of India, at local and landscape scales [39].
5. Conclusions
The species composition of the Pasochoa herb community appeared to be structured by topographic and light intensity microhabitat variables. Lack of understory light seems to shape the herb ground cover. Light intensity was more important than topographic microhabitats. Both light and topography influenced the distribution of species and the community structure of herbs. Other variables, such as soil nutrients and tree biomass, which were not measured, could be important drivers of the floristic composition of herbs. Herbs had more species than trees in the present study site (50 vs. 32); at the same time, the herb genera represent a great phylogenetic richness, which illustrates that the Andean herbs deserve to be focused on strongly in conservation strategies as most of them are poorly collected and underestimated in botanical surveys. There is an urgent need to improve the knowledge of the composition of tropical plant herbs, which is limited to floristic inventories, without ecological information. In the future, new studies with information on epiphytes and ferns will shed some light on the floristic composition of Andean forests and will predict changes in plant communities under rapid climate change conditions in order to design strategies for tropical Andean ecosystems.
Author Contributions
Conceptualization, C.Q., R.V. and H.B.; methodology, C.Q. and R.V.; formal analysis, C.Q. and R.V.; investigation, C.Q. and R.V.; resources, C.Q.; data curation, C.Q.; writing—original draft preparation, C.Q.; writing—review and editing, C.Q., R.V. and H.B.; funding acquisition, C.Q. and H.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Pontificia Universidad Católica of Ecuador, grant number QINV0450-IINV529010100.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Botanical collections can be found at https://bioweb.bio/ (accessed on 13 April 2024.
Acknowledgments
We would like to thank V. Quitigüiña, P. Moreno, S. Aldaz, D. Vela, G. Basántez, S. León, C. Vélez, I. Yépez, M. Guerrón, S. Valencia, L. Medina, M. Valencia, B. Larrea, J. Celi, J. Barahona, and P. Vélez for their help during the field work. Moreover, we thank B. Øllgaard, J. Jaramillo †, H. Navarrete, K. Romoleroux, B. Ståhl, S. León, and L. Endara for the identification of species. Mapresa Ltd.a. permitted access to the study area.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gentry, A.H.; Dodson, C. Contribution of Nontrees to Species Richness of a Tropical Rain Forest. Biotropica 1987, 19, 149–156. [Google Scholar] [CrossRef]
- Galeano, G.; Suárez, S.; Balslev, H. Vascular Plant Species Count in a Wet Forest in the Choco Area on the Pacific Coast of Colombia. Biodivers. Conserv. 1998, 7, 1563–1575. [Google Scholar] [CrossRef]
- Wright, J.S. Plant Diversity in Tropical Forests: A Review of Mechanisms of Species Coexistence. Oecologia 2002, 130, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Valencia, R.; Condit, R.; Foster, R.B.; Romoleroux, K.; Villa Munoz, G.; Svenning, J.C.; Magard, E.; Bass, M.; Losos, E.C.; Balslev, H. Yasuni Forest Dynamics Plot, Ecuador. In Tropical Forest Diversity and Dynamism: Findings from a Large-Scale Plot Network; The University of Chicago Press: Chicago, IL, USA, 2004. [Google Scholar]
- Esquivel-Muelbert, A.; Pugh, T.; Baker, T.; Dexter, K.; Lewis, S.; Galbraith, D.; Phillips, O. Vulnerability of Amazonian Tree Communities to Global Change. In Proceedings of the EGU General Assembly Conference, Online, 4–8 May 2020. [Google Scholar]
- Banda, K.R.; Delgado-Salinas, A.; Dexter, K.G.; Linares-Palomino, R.; Oliveira-Filho, A.; Prado, D.; Pullan, M.; Quintana, C.; Riina, R.; Rodríguez, G.M.; et al. Plant Diversity Patterns in Neotropical Dry Forests and Their Conservation Implications. Science 2016, 353, 1383–1387. [Google Scholar] [CrossRef]
- Guevara Andino, J.E.; Hernández, C.; Valencia, R.; Forrister, D.; Endara, M.J. Accelerating the Discovery of Rare Tree Species in Amazonian Forests: Integrating Long Monitoring Tree Plot Data with Metabolomics and Phylogenetics for the Description of a New Species in the Hyperdiverse Genus Inga Mill. PeerJ 2022, 10, e13767. [Google Scholar] [CrossRef] [PubMed]
- Spicer, M.E.; Mellor, H.; Carson, W.P. Seeing beyond the Trees: A Comparison of Tropical and Temperate Plant Growth Forms and Their Vertical Distribution. Ecology 2020, 101, e02974. [Google Scholar] [CrossRef] [PubMed]
- Cicuzza, D.; Kessler, M.; Pitopang, R.; Tjitrosoedirdjo, S.; Gradstein, S.R. Terrestrial Herb Communities of Tropical Submontane and Tropical Montane Forests in Central Sulawesi, Indonesia. In Environmental Science and Engineering; Springer Science and Business Media Deutschland GmbH: Berlin/Heidelberg, Germany, 2010; pp. 377–390. [Google Scholar]
- Willinghöfer, S.; Cicuzza, D.; Kessler, M. Elevational Diversity of Terrestrial Rainforest Herbs: When the Whole Is Less than the Sum of Its Parts. Plant Ecol. 2012, 213, 407–418. [Google Scholar] [CrossRef]
- Vieira, L.T.A.; Polisel, R.T.; Ivanauskas, N.M.; Shepherd, G.J.; Waechter, J.L.; Yamamoto, K.; Martins, F.R. Geographical Patterns of Terrestrial Herbs: A New Component in Planning the Conservation of the Brazilian Atlantic Forest. Biodivers. Conserv. 2015, 24, 2181–2198. [Google Scholar] [CrossRef]
- Whitmore, T.C.; Peralta, R.; Brown, K. Total Species Count in a Costa Rican Tropical Rain Forest. J. Trop. Ecol. 1985, 1, 375–378. [Google Scholar] [CrossRef]
- Poulsen, A.D.; Balslev, H. Abundance and Cover of Ground Herbs in an Amazonian Rain Forest. J. Veg. Sci. 1991, 2, 315–322. [Google Scholar] [CrossRef]
- Poulsen, A.D.; Pendry, C.A. Inventories of Ground Herbs at Three Altitudes on Bukit Belalong, Brunei, Borneo. Biodivers. Conserv. 1995, 4, 745–757. [Google Scholar] [CrossRef]
- Poulsen, A.D. Species Richness and Density of Ground Herbs within a Plot of Lowland Rainforest in North-West Borneo. J. Trop. Ecol. 1996, 12, 177–190. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K. Distribution of Pteridophyta and Melastomataceae along an Edaphic Gradient in an Amazonian Rain Forest. J. Veg. Sci. 1994, 5, 25–34. [Google Scholar] [CrossRef]
- Grubb, P.J.; Lloyd, J.R.; Pennington, T.D.; Whitmore, T.C. A Comparison of Montane and Lowland Rain Forest in Ecuador I. The Forest Structure, Physiognomy, and Floristics. J. Ecol. 1963, 51, 567–601. [Google Scholar] [CrossRef]
- Young, K.R.; León, B. Pteridophyte Species Diversity in the Central Peruvian Amazon: Importance of Edaphic Specialization. Brittonia 1989, 41, 388–395. [Google Scholar] [CrossRef]
- Linares-Palomino, R.; Cardona, V.; Hennig, E.I.; Hensen, I.; Hoffmann, D.; Lendzion, J.; Soto, D.; Herzog, S.K.; Kessler, M. Non-Woody Life-Form Contribution to Vascular Plant Species Richness in a Tropical American Forest. Plant Ecol. 2009, 201, 87–99. [Google Scholar] [CrossRef]
- Tuomisto, H.; Poulsen, A.D.; Ruokolainen, K.; Moran, R.C.; Quintana, C.; Celi, J.; Cañas, G. Linking Floristic Patterns with Soil Heterogeneity and Satellite Imagery in Ecuadorian Amazonia. Ecol. Appl. 2003, 13, 352–371. [Google Scholar] [CrossRef]
- Tuomisto, H.; Ruokolainen, K.; Poulsen, A.D.; Moran, R.C.; Quintana, C.; Cañas, G.; Celi, J. Distribution and Diversity of Pteridophytes and Melastomataceae along Edaphic Gradients in Yasuní National Park, Ecuadorian Amazonia. Biotropica 2002, 34, 516–533. [Google Scholar] [CrossRef]
- Quintana, C.; Girardello, M.; Barfod, A.S.; Balslev, H. Diversity Patterns, Environmental Drivers and Changes in Vegetation Composition in Dry Inter-Andean Valleys. J. Plant Ecol. 2017, 10, rtw036–475. [Google Scholar] [CrossRef]
- Llambí, L.D.; Rada, F. Ecological Research in the Tropical Alpine Ecosystems of the Venezuelan Páramo: Past, Present and Future. Plant Ecol. Divers. 2019, 12, 519–538. [Google Scholar] [CrossRef]
- Esteban Suárez, R.; Medina, G. Vegetation Structure and Soil Properties in Ecuadorian Páramo Grasslands with Different Histories of Burning and Grazing. Arctic, Antarct. Alp. Res. 2001, 33, 158–164. [Google Scholar] [CrossRef]
- Grubb, P.J.; Lloyd, J.R.; Pennington, T.D.; Páez-Bimos, S. A Historical Baseline Study of the Páramo of Antisana in the Ecuadorian Andes Including the Impacts of Burning, Grazing and Trampling. Plant Ecol. Divers. 2020, 13, 225–256. [Google Scholar] [CrossRef]
- Cruz, M.; Lasso, E. Insights into the Functional Ecology of Páramo Plants in Colombia. Biotropica 2021, 53, 1415–1431. [Google Scholar] [CrossRef]
- Keating, P.L.; Young, K.R.; León, B. Variation in High Andean Vegetation at a Site in Southwestern Ecuador. Pa. Geogr. 2002, 40, 15–35. [Google Scholar]
- Ramsay, P.M.; Oxley, E.R.B. The Growth Form Composition of Plant Communities in the Ecuadorian Paramos. Plant Ecol. 1997, 131, 173–192. [Google Scholar] [CrossRef]
- Leon-Garcia, I.V.; Lasso, E. High Heat Tolerance in Plants from the Andean Highlands: Implications for Paramos in a Warmer World. PLoS ONE 2019, 14, e0224218. [Google Scholar] [CrossRef] [PubMed]
- Campos, P.V.; Villa, P.M.; Nunes, J.A.; Schaefer, C.E.G.R.; Porembski, S.; Neri, A.V. Plant Diversity and Community Structure of Brazilian Páramos. J. Mt. Sci. 2018, 15, 1186–1198. [Google Scholar] [CrossRef]
- Lozano, P.; Cabrera, O.; Peyre, G.; Cleef, A.; Toulkeridis, T. Plant Diversity and Composition Changes along an Altitudinal Gradient in the Isolated Volcano Sumaco in the Ecuadorian Amazon. Diversity 2020, 12, 229. [Google Scholar] [CrossRef]
- Sklenár, P.; Ramsay, P.M. Diversity of Zonal Páramo Plant Communities in Ecuador. Divers. Distrib. 2001, 7, 113–124. [Google Scholar] [CrossRef]
- Ramsay, P.M. Gradients in a Tropical Mountain Ecosystem of Ecuador. Mt. Res. Dev. 2008, 28, 340–341. [Google Scholar] [CrossRef]
- Tokarz, E.L.; Álvia, P.; Valencia, R.; Queenborough, S.A. Habitat Filtering of Six Coexisting Heliconia Species in a Lowland Tropical Rain Forest in Amazonian Ecuador. J. Trop. Ecol. 2019, 35, 91–94. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, J.; Xiao, L.; Zhong, Z.; Wang, Q.; Wang, W. Dominant Species Abundance, Vertical Structure and Plant Diversity Response to Nature Forest Protection in Northeastern China: Conservation Effects and Implications. Forests 2020, 11, 295. [Google Scholar] [CrossRef]
- Dar, J.A.; Sundarapandian, S. Patterns of Plant Diversity in Seven Temperate Forest Types of Western Himalaya, India. J. Asia-Pacific Biodivers. 2016, 9, 280–292. [Google Scholar] [CrossRef]
- Royo, A.A.; Carson, W.P. The Herb Community of a Tropical Forest in Central Panamá: Dynamics and Impact of Mammalian Herbivores. Oecologia 2005, 145, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.Y.; Yang, K.C.; Hsieh, C.F.; Hsieh, C.-H.; Su, M.H. Local Neighborhood Communities in the Understory Play a Critical Role by Affecting Regeneration Niches and Subsequent Community Assembly in a Montane Cloud Forest. Ecol. Res. 2017, 32, 821–833. [Google Scholar] [CrossRef]
- Radhamoni, H.V.N.; Queenborough, S.A.; Arietta, A.Z.A.; Suresh, H.S.; Dattaraja, H.S.; Kumar, S.S.; Sukumar, R.; Comita, L.S. Local- and Landscape-Scale Drivers of Terrestrial Herbaceous Plant Diversity along a Tropical Rainfall Gradient in Western Ghats, India. J. Ecol. 2023, 111, 1021–1036. [Google Scholar] [CrossRef]
- Sierra, R. Patrones Y Factores De Deforestación En El Ecuador Continental. Conserv. Int. Ecuador For. Trends 2013, 1, 22. [Google Scholar]
- Valencia, R.; Jorgensen, P.M. Composition and Structure of a Humid Montane Forest on the Pasochoa Volcano, Ecuador. Nord. J. Bot. 1992, 12, 239–247. [Google Scholar] [CrossRef]
- Hall, M.L.; Samaniego, P.; Le Pennec, J.L.; Johnson, J.B. Ecuadorian Andes Volcanism: A Review of Late Pliocene to Present Activity. J. Volcanol. Geotherm. Res. 2008, 176, 1–6. [Google Scholar] [CrossRef]
- Ministerio del Ambiente Areas Protegidas Del Ecuador. Available online: http://areasprotegidas.ambiente.gob.ec/ (accessed on 13 April 2024).
- Stern, M.J. An Inter-Andean Forest Relict: Vegetation Change on Pasochoa Volcano, Ecuador. Mt. Res. Dev. 1995, 15, 339–348. [Google Scholar] [CrossRef]
- Jorgensen, P.M.; Ulloa Ulloa, C.; Madsen, J.E.; Valencia, R., R. A Floristic Analysis of the High Andes of Ecuador. In Biodiversity and Conservation of Neotropical Montane Forests; New York Botanical Garden: Bronx, NY, USA, 1995. [Google Scholar]
- Curtis, J.T.; McIntosh, R.P. An Upland Forest Continuum in the Prairie-Forest Border Region of Wisconsin. Ecology 1951, 32, 476–496. [Google Scholar] [CrossRef]
- IUCN Standards and Petitions Committee. Guidelines for Using the IUCN Red List Categories and Criteria. Eur. J. Taxon. 2021, 2021, 1–122. [Google Scholar] [CrossRef]
- León-yánez, S.; Valencia, R.; Pitman, N.; Endara, L.; Ulloa, C.U.; Navarrete, H. Libro Rojo de Las Plantas Endémicas del Ecuador; Ediciones del Herbario QCA, Pontificia Universidad Católica del Ecuador, Quito 2011; Pages 1-440.
- Legendre, P. Comparison of Permutation Methods for the Partial Correlation and Partial Mantel Tests. J. Stat. Comput. Simul. 2000, 67, 37–73. [Google Scholar] [CrossRef]
- IUCN IUCN Red List Categories and Criteria: Version 3.1.; Gland. 2001.
- UNEP World Conservation Monitoring Centre Checklist of CITES Species; UNEP World Conservation Monitoring Centre: Cambidge, UK, 2003.
- Jørgensen, P.M.; Ulloa Ulloa, C. Seed Plants of the High Andes of Ecuador—A Checklist. AAU reports 1994, 34, 1–443. [Google Scholar]
- Renner, S.S. Reproduction and Evolution in Some Genera of Neotropical Melastomataceae. Mem. N. Y. Bot. Gard. 1990, 55, 143–152. [Google Scholar]
- Zizka, A.; ter Steege, H.; Pessoa, M.d.C.R.; Antonelli, A. Finding Needles in the Haystack: Where to Look for Rare Species in the American Tropics. Ecography 2018, 41, 321–330. [Google Scholar] [CrossRef]
- Prather, L.A.; Alvarez-Fuentes, O.; Mayfield, M.H.; Ferguson, C.J. Implications of the Decline in Plant Collecting for Systematic and Floristic Research. Syst. Bot. 2004, 29, 216–220. [Google Scholar] [CrossRef]
- Lendemer, J.; Thiers, B.; Monfils, A.K.; Zaspel, J.; Ellwood, E.R.; Bentley, A.; Levan, K.; Bates, J.; Jennings, D.; Contreras, D.; et al. The Extended Specimen Network: A Strategy to Enhance US Biodiversity Collections, Promote Research and Education. Bioscience 2020, 70, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Monfils, A.K.; Powers, K.E.; Marshall, C.J.; Martine, C.T.; Smith, J.F.; Prather, L.A. Natural History Collections: Teaching about Biodiversity Across Time, Space, and Digital Platforms. Southeast. Nat. 2017, 16, 47–57. [Google Scholar] [CrossRef]
- Moran, R.C. The Importance of Mountains to Pteridophytes, with Emphasis on Neotropical Montane Forests. In Biodiversity and Conservation of Neotropical Montane Forests; New York Botanical Garden: Bronx, NY, USA, 1995; pp. 359–363. [Google Scholar]
- Ollgaard, B. Diversity of Huperzia (Lycopodiaceae) in Neotropical Montane Forests. In Biodiversity and Conservation of Neotropical Montane Forests; New York Botanical Garden: Bronx, NY, USA, 1995; pp. 349–358. [Google Scholar]
- Mandl, N.; Lehnert, M.; Kessler, M.; Gradstein, S.R. A Comparison of Alpha and Beta Diversity Patterns of Ferns, Bryophytes and Macrolichens in Tropical Montane Forests of Southern Ecuador. Biodivers. Conserv. 2010, 19, 2359–2369. [Google Scholar] [CrossRef]
- Hubbell, S.P. Tropical Rain Forest Conservation and the Twin Challenges of Diversity and Rarity. Ecol. Evol. 2013, 3, 3263–3274. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.B.; Swaine, M.D. Distribution and Ecology of Vascular Plants in a Tropical Rain Forest; Springer Science and Business Media LLC: Dordrecht, Netherlands, 1981. [Google Scholar]
- YOUNG, K. Floristic Diversity on the Eastern Slopes of the Peruvian Andes. Candollea 1991, 46, 125–143. [Google Scholar]
- Pérez-Escobar, O.A.; Zizka, A.; Bermúdez, M.A.; Meseguer, A.S.; Condamine, F.L.; Hoorn, C.; Hooghiemstra, H.; Pu, Y.; Bogarín, D.; Boschman, L.M.; et al. The Andes through Time: Evolution and Distribution of Andean Floras. Trends Plant Sci. 2022, 27, 364–378. [Google Scholar] [CrossRef] [PubMed]
- G.S.P.H. The Tropical Rain Forest. By, P.W. Richards. Cambridge University Press, 1952. Pp. Xviii, 450; 43 Figs., 15 Plates, 39 Tables. 63s. Q. J. R. Meteorol. Soc. 1953, 79, 312. [Google Scholar] [CrossRef]
- Kabakoff, R.P.; Chazdon, R.L. Effects of Canopy Species Dominance on Understorey Light Availability in Low-Elevation Secondary Forest Stands in Costa Rica. J. Trop. Ecol. 1996, 12, 779–788. [Google Scholar] [CrossRef]
Figure 1.
Topographic map of the one-hectare forest plot at 3300 m on the Pasochoa volcano, Ecuador, showing the subdivision into 5 × 5 m units. The relative height varies from 0 to 67 m. Each subunit was classified as one of the three topographic microhabitats. The ridge covered 18% of the study area, the gully bed covered 12% of the study area (black lines in the map), and the slope covered 70% of the study area. Each subunit was classified as one of three light intensity microhabitats.
Figure 1.
Topographic map of the one-hectare forest plot at 3300 m on the Pasochoa volcano, Ecuador, showing the subdivision into 5 × 5 m units. The relative height varies from 0 to 67 m. Each subunit was classified as one of the three topographic microhabitats. The ridge covered 18% of the study area, the gully bed covered 12% of the study area (black lines in the map), and the slope covered 70% of the study area. Each subunit was classified as one of three light intensity microhabitats.

Figure 2.
Genus and species richness for the 18 herb families found in the one-hectare plot in the Pasochoa volcano. The pteridophytes are represented by one group.
Figure 2.
Genus and species richness for the 18 herb families found in the one-hectare plot in the Pasochoa volcano. The pteridophytes are represented by one group.

Figure 3.
Comparison of the species herb number according to the life forms in a montane forest (3300 m) in this study and a tropical rain forest plot 265 m in Cuyabeno [13].
Figure 3.
Comparison of the species herb number according to the life forms in a montane forest (3300 m) in this study and a tropical rain forest plot 265 m in Cuyabeno [13].

Figure 4.
Species with the highest Importance Value Index of the one-hectare plot at Pasochoa volcano. (IVI) = relative frequency + relative cover + relative density.
Figure 4.
Species with the highest Importance Value Index of the one-hectare plot at Pasochoa volcano. (IVI) = relative frequency + relative cover + relative density.

Figure 5.
Examples of the topographic distribution of four species of herbs found in the study area: (a) Begonia sodiroi, climber, N = 428; (b) Hydrocotyle alchemilloides, prototerrestrial, N = 260; (c) Carex hamata, prototerrestrial, N = 7116; (d) Asplenium pteroporus, prototerrestrial, N = 347. (e) Low light microhabitats (thicket) and (f) medium light intensity microhabitats. Level curves each 2 m.
Figure 5.
Examples of the topographic distribution of four species of herbs found in the study area: (a) Begonia sodiroi, climber, N = 428; (b) Hydrocotyle alchemilloides, prototerrestrial, N = 260; (c) Carex hamata, prototerrestrial, N = 7116; (d) Asplenium pteroporus, prototerrestrial, N = 347. (e) Low light microhabitats (thicket) and (f) medium light intensity microhabitats. Level curves each 2 m.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Terrestrial herbs found in the 1 ha plot in a montane rain forest at 3300 m on the Pasochoa volcano, Ecuador. The 50 herb species are grouped into obligate terrestrial (O-P = prototerrestrial, O-S = scandent) and facultative terrestrial (F-C = climber F-E = epiphyte). N = individuals number/ha. C = coverage in m2/ha. Microhabitat in which the species were found, as follows: r = ridge, s = slope, g = gully bed, h = high light intensity, m = medium light intensity, and l = low light intensity. Conservation status according to the IUCN categories [50]: NT = near threatened, LC = least concern, CA II = CITES Appendix II [51]. Proposed conservation status over Ecuador: WC = well collected >100 collections all over Ecuador, MC = medium collected <100–>30, SC = scarcely collected <30 collections. The collection number for the vouchers is between parentheses, CQ = Catalina Quintana et al. The vouchers are deposited in the QCA and QCNE herbaria.
Table 1.
Terrestrial herbs found in the 1 ha plot in a montane rain forest at 3300 m on the Pasochoa volcano, Ecuador. The 50 herb species are grouped into obligate terrestrial (O-P = prototerrestrial, O-S = scandent) and facultative terrestrial (F-C = climber F-E = epiphyte). N = individuals number/ha. C = coverage in m2/ha. Microhabitat in which the species were found, as follows: r = ridge, s = slope, g = gully bed, h = high light intensity, m = medium light intensity, and l = low light intensity. Conservation status according to the IUCN categories [50]: NT = near threatened, LC = least concern, CA II = CITES Appendix II [51]. Proposed conservation status over Ecuador: WC = well collected >100 collections all over Ecuador, MC = medium collected <100–>30, SC = scarcely collected <30 collections. The collection number for the vouchers is between parentheses, CQ = Catalina Quintana et al. The vouchers are deposited in the QCA and QCNE herbaria.
| Family | Species | Life Form | N | C | Topographic Micro Habitat | Light Micro Habitat | Iucn Conservation Status | Proposed Conservation Status | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | ARALIACEAE | Hydrocotyle alchemilloides A. Rich. (CQ 60) | O-P | 260 | 1.3 | s,r | h | - | SC |
| 2 | ARACEAE | Anthurium pulchrum Engl. (CQ 217) | F-C | 276 | 18.9 | s,r | h | - | WC |
| 3 | ASTERACEAE | Asteraceae sp. (CQ 126) | O-S | 41 | 1.5 | s | m | - | - |
| 4 | Ageratum sp. (CQ 233) | F-C | 16 | 0.7 | s,r,g | m,l | - | - | |
| 5 | BASELLACEAE | Tournonia hookeriana Moq. (CQ 211) | F-C | 189 | 7.8 | s | m,l | - | SC |
| 6 | BEGONIACEAE | Begonia sodiroi C. DC. (CQ 206) | F-C | 428 | 39.2 | s,r,g | h,m | NT | SC |
| 7 | CARYOPHYLLACEAE | Arenaria lanuginosa (Michx.) Rohrb. (CQ 110) | O-S | 35 | 1.5 | s,r | h,m,l | - | MC |
| 8 | Stellaria recurvata Willd. ex D.F.K. Schltdl. (CQ 143) | O-S | 59 | 3.1 | s,r,g | h,m,l | LC | WC | |
| 9 | CYPERACEAE | Carex hamata Sw. (CQ 213) | O-P | 7116 | 175 | s,r,g | h,m,l | - | WC |
| 10 | Carex muricata L. (CQ 191) | O-P | 2099 | 28.9 | s,r,g | h,m,l | - | MC | |
| 11 | Rhynchospora macrochaeta Steudel ex Boeckeler (CQ 209) | O-P | 1702 | 90 | s,r,g | h,m | - | WC | |
| 12 | FERNS | Adiantum concinnum Humb. and Bonpl. ex Willd. (CQ 245) | O-P | 6 | 0.7 | r,g | m | - | MC |
| 13 | Asplenium pteropus Kaulf (CQ 150) | O-P | 347 | 19.8 | s,r,g | h,m | - | WC | |
| 14 | Asplenium sessilifolium Desv. (CQ 190) | O-P | 1466 | 46.7 | s,r,g | h,m,l | - | MC | |
| 15 | Asplenium monanthes L. (CQ 153) | O-P | 187 | 7.5 | s,r,g | h,m | - | WC | |
| 16 | Asplenium castaneum Schltdl. and Cham. (CQ 132) | O-P | 475 | 8.1 | s,r,g | h,m | - | SC | |
| 17 | Asplenium peruvianum Desv. (CQ 124) | O-P | 110 | 1.9 | s,r,g | h,m | - | SC | |
| 18 | Asplenium squamosum L. (CQ 247) | O-P | 10 | 0.8 | g | h,m | - | SC | |
| 19 | Athyrium dombeyi Desv. (CQ 116) | O-P | 86 | 9.3 | s,r,g | h,m | - | SC | |
| 20 | Blechnum fragile (Liebm.) C.V. Morton and Lellinger (CQ 255) | O-S | 23 | 1.5 | g | m | - | MC | |
| 21 | Botrychium virginianum (L.) Sw. (CQ 144) | O-P | 41 | 1.4 | s | h,m | - | SC | |
| 22 | Campyloneurum amphostenon (Klotzch) Fée (CQ 155) | F-E | 31 | 0.9 | s,r,g | m | - | MC | |
| 23 | Dryopteris paleacea (Sw.) Hand.-Mazz. (CQ 112) | O-P | 72 | 40.5 | s,r,g | h,m | - | SC | |
| 24 | Elaphoglossum papillosum (Baker) Christ (CQ 100) | F-E | 2 | 0.1 | s | h,m | - | WC | |
| 25 | Equisetum bogotense Kunth (CQ 231) | O-P | 48 | 0.7 | s | h,m | - | WC | |
| 26 | Hypolepis bogotensis H. Karst. (CQ 258) | O-P | 260 | 0.9 | s | h,m | - | MC | |
| 27 | Pecluma divaricata (E. Fourn.) Mickel and Beitel (CQ 229) | O-P | 8 | 1 | s,g | h,m | - | MC | |
| 28 | Polypodium subandinum Sodiro (CQ 193) | F-C | 4620 | 43.7 | s,r,g | h,m,l | - | WC | |
| 29 | Polystichum muricatum (L.) Fée (CQ 67) | O-P | 737 | 50.6 | s,r,g | h,m | - | SC | |
| 30 | Pteris muricata Hook. (CQ 196) | O-P | 35 | 4.5 | g | h,m | - | MC | |
| 31 | Thelypteris rigescens (Sodiro) A.R. Sm. (CQ 23) | O-P | 102 | 68 | s,r,g | h,m,l | - | SC | |
| 32 | GERANIACEAE | Geranium laxicaule R. Knuth (CQ21) | O-S | 27 | 0.5 | s,r | h,m | - | SC |
| 33 | GESNERIACEAE | Alloplectus dielsii (Mansf.) Wiehler (CQ 197) | O-S | 451 | 19.3 | s,r,g | h,m | - | WC |
| 34 | ORCHIDACEAE | Baskervilla auriculata Garay (CQ 113) | O-P | 30 | 0.9 | s,r | h | NT | SC |
| 35 | Cranichis ciliata Kunth (CQ 135) | O-P | 208 | 6.2 | s,r | h | CA II | MC | |
| 36 | Malaxis hoppii (Schltr.)Løjtnant (CQ 119) | O-P | 406 | 6.8 | s,r,g | h | CA II | SC | |
| 37 | Stelis pusilla Kunth (CQ 111) | F-E | 59 | 3.1 | s,g | h,m | CA II | WC | |
| 38 | Odontoglossum pardinum (Lindl.)Lindl.(CQ 223) | F-E | 10 | 0.1 | s,r | h,m | CA II | MC | |
| 39 | Pleurothallis sp. (CQ 240) | F-E | 2 | 0.01 | s | h,m | - | - | |
| 40 | OXALIDACEAE | Oxalis lotoides Kunth (CQ 75) | O-S | 115 | 7.5 | s,r,g | h,m,l | - | WC |
| 41 | PHYTOLACCACEAE | Phytolacca bogotensis Kunth (CQ 270) | O-P | 14 | 0.6 | s,r | h,m | - | MC |
| 42 | PIPERACEAE | Peperomia rotundata Kunth (CQ 33) | F-C | 4860 | 58.1 | s,r,g | h,m,l | - | WC |
| 43 | Peperomia sp. 1 (CQ 224) | F-C | 102 | 1.5 | s,g | h,m | - | - | |
| 44 | Peperomia sp. 2 (CQ 243) | F-C | 24 | 0.6 | g | m | - | - | |
| 45 | POACEAE | Poa aequatoriensis Hack. (CQ 114) | O-P | 130 | 3.7 | s,r | h,m | - | SC |
| 46 | RUBIACEAE | Galium obovatum Kunth (CQ 133) | O-S | 39 | 0.5 | s,r | h,m | - | SC |
| 47 | Leptostigma pilosum (Benth.) Fosberg (CQ 106) | O-P | 1159 | 19.9 | s,r | h | - | SC | |
| 48 | SOLANACEAE | Solanum dalibardiforme Bitter (CQ 138) | F-C | 97 | 3.6 | s,r,g | m,l | - | SC |
| 49 | URTICACEAE | Pilea pubescens Liebm. (CQ 117) | F-C | 1883 | 7 | s,r | h,m | - | MC |
| 50 | Urtica leptophylla Kunth (CQ 249) | O-P | 8 | 0.9 | s | m | - | MC |
Table 2.
Species richness of herbs in different studies and altitudes. Plot area varies in most studies. All studies conducted in Ecuador to date are listed, including the present study (in bold). (-) Studies where the number of individuals per species is not reported and (*) studies where obligate terrestrial herbs are not distinguished.
Table 2.
Species richness of herbs in different studies and altitudes. Plot area varies in most studies. All studies conducted in Ecuador to date are listed, including the present study (in bold). (-) Studies where the number of individuals per species is not reported and (*) studies where obligate terrestrial herbs are not distinguished.
| Herbs Species/Individuals | Obligate Terrestrial Herbs Species/Individuals | Density (Number Individuals/m2) | Plot Area m2 | Altitude m | Study Area | Reference |
|---|---|---|---|---|---|---|
| 0.046 | * | 1.54 | 464 | 1710 | Ecuador (Borja) | [17] |
| 0.087 | * | 1.53 | 464 | 520 | Ecuador (Shinguipino) | [17] |
| - | 0.134 | 1.19 | 100 | 100 | Costa Rica | [12] |
| 0.04 | 0.03 | 0.03 | 100 | 3300 | Ecuador (Pasochoa) | This study |
| - | 0.018 | 2.85 | 1000 | 220 | Ecuador (Capeira) | [1] |
| - | 0.019 | 0.94 | 1000 | 220 | Ecuador (Jauneche) | [1] |
| - | 0.041 | 1.22 | 1000 | 220 | Ecuador (Palenque) | [1] |
| 0.009 | 0.005 | 1.1 | 10,000 | 250 | Ecuador (Cuyabeno) | [13] |
| 0.015 | * | 0.63 | 10,000 | 250 | Borneo (Brunei) | [15] |
| 0.027 | * | 0.24 | 10,000 | 250 | Borneo (Brunei) | [15] |
| 0.014 | 0.008 | 0.31 | 1000 | 3300 | Ecuador (Pasochoa) | This study |
| 0.008 | * | 2.67 | 30,000 | 900–1100 | Bolivia (Los Volcanes Reserve) | [19] |
| 0.002 | 0.011 | 3.11 | 10,000 | 3300 | Ecuador (Pasochoa) | This study |
Table 3.
Mantel and partial Mantel correlations of the herb composition similarity matrix with space and microhabitat matrices. Matrix B is the matrix correlated with the herb flora composition similarity matrix. Matrix C is the matrix whose effect is removed when calculating the partial correlations.
Table 3.
Mantel and partial Mantel correlations of the herb composition similarity matrix with space and microhabitat matrices. Matrix B is the matrix correlated with the herb flora composition similarity matrix. Matrix C is the matrix whose effect is removed when calculating the partial correlations.
| Matrix B | r | P | |
|---|---|---|---|
| Topography | 0.2 | 0.01 | |
| Space | 0.09 | 0.04 | |
| Light (presence/absence thicket) | 0.21 | 0.001 | |
| Matrix B | Matrix C | Partial r | P |
| Topography | Space | 0.187 | 0.0001 |
| Space | Topography | 0.02 | 0.33 |
| Topography | Light (presence/absence thicket) | 0.183 | 0.001 |
| Light (presence/absence thicket) | Topography | 0.193 | 0.007 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Quintana, C.; Balslev, H.; Valencia, R. Species Composition and Distribution of Terrestrial Herbs in a High Montane Forest in Ecuador. Diversity 2024, 16, 262. https://0-doi-org.brum.beds.ac.uk/10.3390/d16050262
AMA Style
Quintana C, Balslev H, Valencia R. Species Composition and Distribution of Terrestrial Herbs in a High Montane Forest in Ecuador. Diversity. 2024; 16(5):262. https://0-doi-org.brum.beds.ac.uk/10.3390/d16050262
Chicago/Turabian StyleQuintana, Catalina, Henrik Balslev, and Renato Valencia. 2024. "Species Composition and Distribution of Terrestrial Herbs in a High Montane Forest in Ecuador" Diversity 16, no. 5: 262. https://0-doi-org.brum.beds.ac.uk/10.3390/d16050262
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.