Detection of Phytoplankton Temporal Anomalies Based on Satellite Inherent Optical Properties: A Tool for Monitoring Phytoplankton Blooms

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Methodology
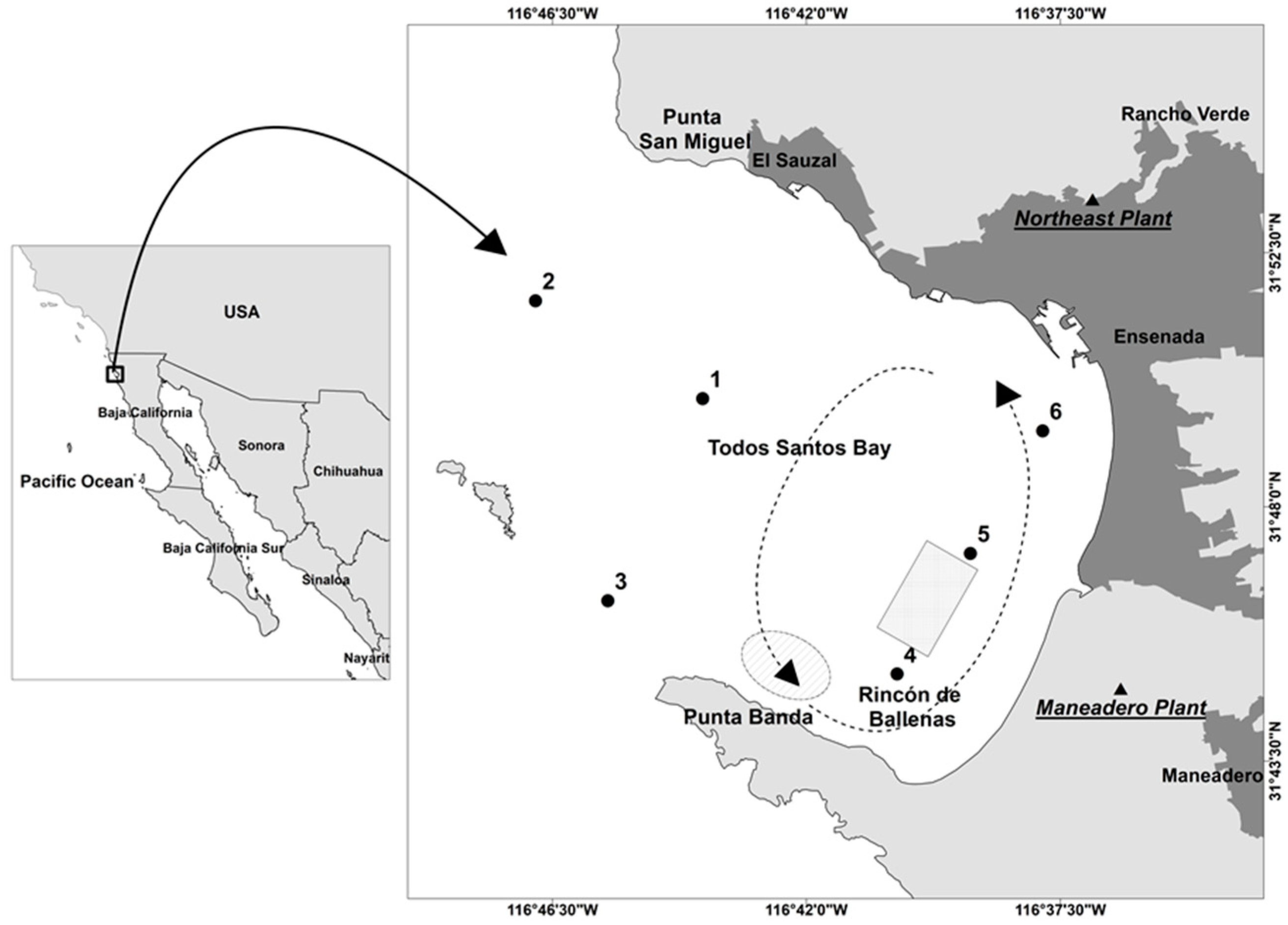
2.1. Study Area and Field Data
2.2. Image Processing and Calculations of the IOP Satellite Index
- is the value to be standardized (each day absorption coefficient);
- is the average of the studied period (for May, all May data since 2003 to 2016; for June, all June data since 2003 to 2016);
- SD is the standard deviation of the studied period (for May, all May data since 2003 to 2016; for June, all June data since 2003 to 2016).
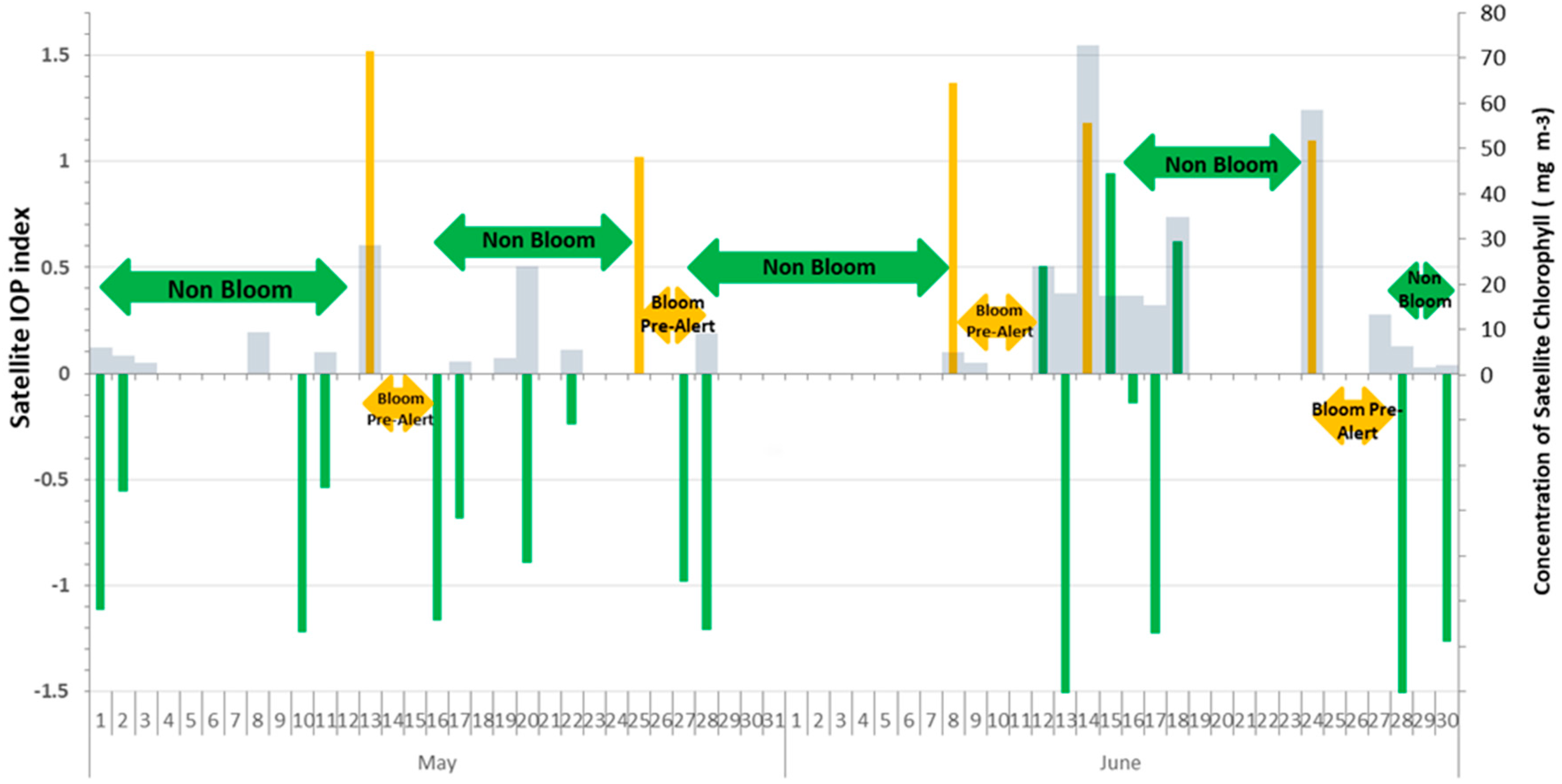
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sanseverino, I.; Conduto, D.; Pozzoli, L.; Dobricic, S.; Lettieri, T. Algal bloom and its economic impact. European Commission; Joint Research Centre Institute for Environment and Sustainability: Ispra, Italy, 2016. [Google Scholar]
- Valentin, J.L. Numerical modelling of phytoplankton bloom in the upwelling ecosystem of Cabo Frio (Brazil). Ecol. Modell. 1999, 116, 135–148. [Google Scholar] [CrossRef]
- Dore, J.E.; Letelier, R.M.; Church, M.J.; Lukas, R.; Karl, D.M. Summer phytoplankton blooms in the oligotrophic North Pacific Subtropical Gyre: Historical perspective and recent observations. Prog. Oceanogr. 2008, 76, 2–38. [Google Scholar] [CrossRef]
- Santamaría-del-Angel, E.; Sebastiá-Frasquet, M.T.; Millán-Nuñez, R.; González-Silvera, A.; Cajal-Medrano, R. Anthropocentric bias in management policies. Are we efficiently monitoring our ecosystem? In Coastal Ecosystems: Experiences and Recommendations for Environmental Monitoring Programs; Sebastiá-Frasquet, M.T., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2015; p. 220. [Google Scholar]
- Santamaría-del-Angel, E.; Soto, I.; Millán-Nuñez, R.; González-Silvera, A.; Wolny, J.; Cerdeira-Estrada, S.; Cajal-Medrano, R.; Muller-Karger, F.; Cannizzaro, J.; Padilla-Rosas, Y.; et al. Experiences and Recommendations for Environmental Monitoring Programs. In Environmental Science, Engineering and Technology; Sebastia-Frasquet, M.T., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2015; p. 32. [Google Scholar]
- Cox, R.F.; Issa, R.R.; Ahrens, D. Management’s Perception of Key Performance Indicators for Construction. J. Constr. Eng. Manag. 2003, 129, 142–151. [Google Scholar] [CrossRef]
- Sebastiá-Frasquet, M.T.; Estornell-Cremades, J.; Rodilla-Alamá, M.; Marti-Gavila, J.; Falco-Giaccaglia, S.L. Estimation of chlorophyll «A» on the Mediterranean coast using a QuickBird image. Revista de Teledetección 2012, 37, 23–33. [Google Scholar] [CrossRef]
- Zheng, G.; DiGiacomo, P.M. Remote sensing of chlorophyll-a in coastal waters based on the light absorption coefficient of phytoplankton. Remote Sens. Environ. 2017, 201, 331–341. [Google Scholar] [CrossRef]
- Cao, F.; Tzortziou, M.; Hu, C.; Mannino, A.; Fichot, C.G.; Del Vecchio, R.; Novak, M. Remote sensing retrievals of colored dissolved organic matter and dissolved organic carbon dynamics in North American estuaries and their margins. Remote Sens. Environ. 2018, 205, 151–165. [Google Scholar] [CrossRef]
- Blondeau-Patissier, D.; Gower, J.F.; Dekker, A.G.; Phinn, S.R.; Brando, V.E. A review of ocean color remote sensing methods and statistical techniques for the detection, mapping and analysis of phytoplankton blooms in coastal and open oceans. Prog. Oceanogr. 2014, 123, 123–144. [Google Scholar] [CrossRef] [Green Version]
- Garver, S.A.; Siegel, D.A. Inherent optical property inversion of ocean color spectra and its biogeochemical interpretation: 1. Time series from the Sargasso Sea. J. Geophys. Res. Oceans 1997, 102, 18607–18625. [Google Scholar] [CrossRef]
- Kahru, M.; Lee, Z.; Kudela, R.M.; Manzano-Sarabia, M.; Greg Mitchell, B. Multi-satellite time series of inherent optical properties in the California Current. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 112, 91–106. [Google Scholar] [CrossRef]
- Kratzer, S.; Moore, G. Inherent optical properties of the baltic sea in comparison to other seas and oceans. Remote Sens. 2018, 10, 418. [Google Scholar] [CrossRef]
- Werdell, P.J.; McKinna, L.I.; Boss, E.; Ackleson, S.G.; Craig, S.E.; Gregg, W.W.; Stramski, D. An overview of approaches and challenges for retrieving marine inherent optical properties from ocean color remote sensing. Prog. Oceanogr. 2018, 160, 186–212. [Google Scholar] [CrossRef] [PubMed]
- Goela, P.C.; Icely, J.; Cristina, S.; Newton, A.; Moore, G.; Cordeiro, C. Specific absorption coefficient of phytoplankton off the Southwest coast of the Iberian Peninsula: A contribution to algorithm development for ocean colour remote sensing. Cont. Shelf Res. 2013, 52, 119–132. [Google Scholar] [CrossRef]
- Soja-Woźniak, M.; Craig, S.; Kratzer, S.; Wojtasiewicz, B.; Darecki, M.; Jones, C. A novel statistical approach for ocean colour estimation of inherent optical properties and cyanobacteria abundance in optically complex waters. Remote Sens. 2017, 9, 343. [Google Scholar] [CrossRef]
- Pavlov, A.K.; Taskjelle, T.; Kauko, H.M.; Hamre, B.; Hudson, S.R.; Assmy, P.; Granskog, M.A. Altered inherent optical properties and estimates of the underwater light field during an Arctic under-ice bloom of Phaeocystis pouchetii. J. Geophys. Res. Oceans 2017, 122, 4939–4961. [Google Scholar] [CrossRef]
- Aguilar-Maldonado, J.A.; Santamaría-del-Ángel, E.; Sebastiá-Frasquet, M.T. Reflectances of SPOT multispectral images associated with the turbidity of the Upper Gulf of California. Revista de Teledetección 2017, 50, 1–16. [Google Scholar] [CrossRef]
- Binding, C.E.; Greenberg, T.A.; McCullough, G.; Watson, S.B.; Page, E. An analysis of satellite-derived chlorophyll and algal bloom indices on Lake Winnipeg. J. Great Lakes Res. 2018, 44, 436–446. [Google Scholar] [CrossRef]
- Aguilar-Maldonado, J.A.; Santamaría-Del-Ángel, E.; González-Silvera, A.; Cervantes-Rosas, O.D.; Sebastiá-Frasquet, M.T. Mapping Satellite Inherent Optical Properties Index in Coastal Waters of the Yucatán Peninsula (Mexico). Sustainability 2018, 10, 1894. [Google Scholar] [CrossRef]
- Kirk, J.T.O. Light and Photosynthesis in Aquatic Ecosystems, 3rd ed.; Cambridge University Press: Cambridge, UK, 2010; p. 638. [Google Scholar]
- Brewin, R.J.; Sathyendranath, S.; Müller, D.; Brockmann, C.; Deschamps, P.Y.; Devred, E.; Doerffer, R.; Fomferra, N.; Franz, B.; Grant, M. The Ocean Colour Climate Change Initiative: III. A round-robin comparison on in-water bio-optical algorithms. Remote Sens. Environ. 2013, 162, 271–294. [Google Scholar] [CrossRef] [Green Version]
- IOCCG. Remote Sensing of Ocean Colour in Coastal, and Other Optically-Complex, Waters. In Reports of the International Ocean-Colour Coordinating Group; Sathyendranath, S., Ed.; No. 3; IOCCG: Dartmouth, NH, Canada, 2000. [Google Scholar]
- Aguilar-Maldonado, J.A.; Santamaría-del-Ángel, E.; González-Silvera, A.; Cervantes-Rosas, O.; López, L.M.; Gutiérrez-Magness, A.; Cerdeira-Estrada, S.; Sebastiá-Frasquet, M.T. Identification of Phytoplankton Blooms under the Index of Inherent Optical Properties (IOP Index) in Optically Complex Waters. Water 2018, 10, 129. [Google Scholar] [CrossRef]
- Cavole, L.M.; Demko, A.M.; Diner, R.E.; Giddings, A.; Koester, I.; Pagniello, C.M.L.S.; Paulsen, M.L.; Ramirez-Valdez, A.; Schwenck, S.M.; et al. Biological impacts of the 2013–2015 warm-water anomaly in the Northeast Pacific: Winners, losers, and the future. Oceanography 2016, 29, 273–285. [Google Scholar] [CrossRef]
- Di Lorenzo, E.; Mantua, N. Multi-year persistence of the 2014/15 North Pacific marine heatwave. Nat. Clim. Chang. 2016, 6, 1042–1047. [Google Scholar] [CrossRef]
- Mkrtchyan, F.A.; Varotsos, C.A. A New Monitoring System for the Surface Marine Anomalies. Water Air Soil Pollut. 2018, 229, 273. [Google Scholar] [CrossRef]
- Gan, R.; Yang, Y.; Ma, Y. Modelling the impacts of the Pacific Ocean sea surface temperature anomalies on a drought event in southwestern China with a piecewise-integration method. Int. J. Clim. 2018, 39, 799–813. [Google Scholar] [CrossRef]
- Santamaría-del-Ángel, E.; Sebastia-Frasquet, M.T.; Gonzalez-Silvera, A.; Aguilar-Maldonado, J.; Mercado-Santana, A.; Herrera-Carmona, J. Uso Potencial de las Anomalías Estandarizadas en la Interpretación de Fenómenos Oceanográficos Globales a Escalas Locales. In Costas y Mares Mexicanos: Construyendo la Línea Base para su Futuro Sostenible, Oceanografía Fisicoquímica; Rivera-Arriaga, E., Sánchéz-Gil, P., Gutiérrez, J., Eds.; Universidad Autónoma de Colima: Colima, México, 2019. [Google Scholar]
- Russell, J.; Benway, H.; Bracco, A.; Deutsch, C.; Ito, T.; Kamenkovich, I.; Patterson, M. Ocean’s Carbon and Heat Uptake: Uncertainties and Metrics; US CLIVAR Report: San Francisco, CA, USA, March 2015. [Google Scholar]
- Yeh, S.W.; Kug, J.S.; Dewitte, B.; Kwon, M.H.; Kirtman, B.; Jin, F.F. El Niño in a changing climate. Nature 2009, 461, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Peña-Manjarrez, J.L.; Gaxiola-Castro, G.; Helenes-Escamilla, J. Environmental factors influencing the variability of Lingulodinium polyedrum and Scrippsiella trochoidea (Dinophyceae) cyst production. Cienc. Mar. 2009, 35, 1–14. [Google Scholar] [CrossRef]
- Cepeda-Morales, J.; Durazo, R.; Millán-Nuñez, E.; De la Cruz-Orozco, M.; Sosa-Ávalos, R.; Espinosa-Carreón, T.L.; Soto-Mardones, L.; Gaxiola-Castro, G. Response of primary producers to the hydrographic variability in the southern region of the California Current System. Cienc. Mar. 2017, 43, 123–135. [Google Scholar] [CrossRef] [Green Version]
- Barocio-León, O.A.; Millán-Núñez, R.; Santamaría-del-Ángel, E.; González-Silvera, A. Phytoplankton primary productivity in the euphotic zone of the California Current System estimated from CZCS imagery. Cienc. Mar. 2007, 33, 59–72. [Google Scholar] [CrossRef] [Green Version]
- INEGI 2015 (Instituto Nacional de Estadística y Geografía/National Institute of Statistic and Geography). Available online: http://www.beta.inegi.org.mx/programas/intercensal/2015/default.html#Tabulados (accessed on 2 August 2018).
- Almazán-Becerril, A.; Aké-Castillo, J.A.; García-Mendoza, E.; Sánchez-Bravo, Y.A.; Escobar-Morales, S.; Valadez-Cruz, F. Catálogo de Microalgas de Bahía de Todos Santos, Baja California; CICESE: Ensenada, Mexico, 2016; ISBN 978-607-95688-7-0. [Google Scholar]
- Gutierrez-Mejia, E.; Lares, M.L.; Huerta-Diaz, M.A.; Delgadillo-Hinojosa, F. Cadmium and phosphate variability during algal blooms of the dinoflagellate Lingulodinium polyedrum in Todos Santos Bay, Baja California, Mexico. Sci. Total Environ. 2016, 541, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Mateos, E.; Marinone, S.G.; Parés-Sierra, A. Towards the numerical simulation of the summer circulation in Todos Santos Bay, Ensenada, B.C. Mexico. Ocean Modell. 2009, 27, 107–112. [Google Scholar] [CrossRef]
- Sommer, U.; Lengfellner, K. Climate change and the timing, magnitude, and composition of the phytoplankton spring bloom. Glob. Chang. Biol. 2008, 14, 1199–1208. [Google Scholar] [CrossRef]
- Winder, M.; Sommer, U. Phytoplankton response to a changing climate. Hydrobiologia 2012, 698, 5. [Google Scholar] [CrossRef]
- IOCCG. Mission Requirements for Future Ocean-Colour Sensors. Reports of the International Ocean-Colour Coordinating Group; McClain, C.R., Meister, G., Eds.; No. 13; IOCCG: Dartmouth, NH, Canada, 2012. [Google Scholar]
- CONAGUA (Comisión Nacional del Agua/National Water Comission of Mexico). Inventory of Wastewater Plants, PTAR for Its Initials in Spanish; CONAGUA: Mexico City, Mexico, 2017. [Google Scholar]
- Orozco-Borbón, M.V.; Rico-Mora, R.; Weisberg, S.B; Noble, R.T.; Dorsey, J.H; Leecaster, M.K.; McGee, C.D. Bacteriological water quality along the Tijuana-Ensenada, Baja California, México shoreline. Mar. Pollut. Bull. 2006, 52, 1190–1196. [Google Scholar] [CrossRef] [PubMed]
- Norma Oficial Mexicana NOM-001-SEMARNAT-1996, Que Establece Los Límites Máximos Permisibles de Contaminantes en las Descargas de Aguas Residuales en Aguas y Bienes Nacionales; SEMARNAT (Secretaría de Medio Ambiente y Recursos Naturales/Ministry of Environment and Natural Resources): Ciudad de México, Mexico, 1997.
- CONAPESCA (Comisión Nacional de Acuacultura y Pesca/National Commission of Aquaculture and Fisheries of Mexico). Statistical Yearbook of Aquaculture and Fisheries Edition 2017; CONAPESCA: Mazatlan, Mexico, 2017. [Google Scholar]
- Dame, R. Bivalve Filter Feeders and Estuarine and Coastal Ecosystem Processes: Conclusions. In Bivalve Filter Feeders; Dame, R.F., Ed.; Nato ASI Series (Series G: Ecological Sciences); Volume 33, Springer: Berlin/Heidelberg, Germany, 1993. [Google Scholar]
- Gregg, W.W.; Casey, N.W. Global and regional evaluation of the SeaWiFS chlorophyll data set. Remote Sens. Environ. 2004, 93, 463–479. [Google Scholar] [CrossRef]
- Santamaría-del-Ángel, E.; Millán Núñez, R.; González-Silvera, A.; Cajal-Medrano, R. Comparison of In Situ and Remotely-Sensed Chl-a concentrations: A Statistical Examination of the Match-up Approach. In Handbook of Satellite Remote Sensing Image Interpretation: Applications for Marine Living Resources Conservation and Management; Morales, J., Stuart, V., Platt, T., Sathyendranath, S., Eds.; EU PRESPO and IOCCG: Dartmouth, NH, Canada, 2011; Chapter 17; pp. 221–238. [Google Scholar]
- COFEPRIS (Comisión Federal Para la Protección Contra Riesgos Sanitarios/Federal Commission for Protection against Health Risks). Lineamiento de Trabajo Para el Muestreo de Fitoplancton y Detección de Biotoxinas Marinas. Available online: http://www.cofepris.gob.mx/AZ/Documents/Lineamiento%20de%20Trabajo%20Muestreo%20y%20Deteccion.pdf (accessed on 27 July 2018).
- Sebastiá, M.T.; Rodilla, M.; Sanchis, J.A.; Altur, V.; Gadea, I.; Falco, S. Influence of nutrient inputs from a wetland dominated by agriculture on the phytoplankton community in a shallow harbour at the Spanish Mediterranean coast. Agric. Ecosyst. Environ. 2012, 152, 10–20. [Google Scholar] [CrossRef]
- Wilson, C. Late summer chlorophyll blooms in the oligotrophic North Pacific subtropical gyre. Geophys. Res. Lett. 2003, 30. [Google Scholar] [CrossRef]
- Villareal, T.A.; Adornato, L.; Wilson, C.; Schoenbaechler, C.A. Summer blooms of diatom-diazotroph assemblages and surface chlorophyll in the North Pacific gyre: A disconnect. J. Geophys. Res. Oceans 2011, 116. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Point | Month | # Observed Days (2003–2016) |
|---|---|---|
| 4 | May | 111 |
| June | 146 | |
| 6 | May | 101 |
| June | 115 |
| Frequency of IOP Index Values (%) | Minimum IOP Index | Maximum IOP Index | |||
|---|---|---|---|---|---|
| <1 | 1–1.6 | >1.6 | |||
| Point 4 May | 81 | 13 | 6 | −1.31 | 5.20 |
| Point 4 June | 85 | 8 | 7 | −0.88 | 5.16 |
| Point 6 May | 79 | 15 | 6 | −1.24 | 3.54 |
| Point 6 June | 87 | 7 | 6 | −1.12 | 4.29 |
| Point 4 May | Point 4 June | Point 6 May | Point 6 June | |
|---|---|---|---|---|
| 2003 | 0 | 17 | 38 | 0 |
| 2004 | 8 | 18 | 9 | 25 |
| 2005 | 25 | 11 | 17 | 20 |
| 2006 | 29 | 10 | 0 | 0 |
| 2007 | 0 | 27 | 0 | 14 |
| 2008 | 11 | 0 | 0 | 0 |
| 2009 | 0 | 20 | 25 | 0 |
| 2010 | 0 | 0 | 0 | 0 |
| 2011 | 0 | 0 | 0 | 11 |
| 2012 | 0 | 0 | 0 | 0 |
| 2013 | 0 | 0 | 0 | 0 |
| 2014 | 0 | 0 | 0 | 0 |
| 2015 | 0 | 0 | 0 | 0 |
| 2016 | 0 | 0 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilar-Maldonado, J.A.; Santamaría-del-Ángel, E.; Gonzalez-Silvera, A.; Sebastiá-Frasquet, M.T. Detection of Phytoplankton Temporal Anomalies Based on Satellite Inherent Optical Properties: A Tool for Monitoring Phytoplankton Blooms. Sensors 2019, 19, 3339. https://0-doi-org.brum.beds.ac.uk/10.3390/s19153339
Aguilar-Maldonado JA, Santamaría-del-Ángel E, Gonzalez-Silvera A, Sebastiá-Frasquet MT. Detection of Phytoplankton Temporal Anomalies Based on Satellite Inherent Optical Properties: A Tool for Monitoring Phytoplankton Blooms. Sensors. 2019; 19(15):3339. https://0-doi-org.brum.beds.ac.uk/10.3390/s19153339
Chicago/Turabian StyleAguilar-Maldonado, Jesús Antonio, Eduardo Santamaría-del-Ángel, Adriana Gonzalez-Silvera, and María Teresa Sebastiá-Frasquet. 2019. "Detection of Phytoplankton Temporal Anomalies Based on Satellite Inherent Optical Properties: A Tool for Monitoring Phytoplankton Blooms" Sensors 19, no. 15: 3339. https://0-doi-org.brum.beds.ac.uk/10.3390/s19153339